 2016, Vol. 36
2016, Vol. 36文章信息
- 徐满厚, 刘敏, 翟大彤, 刘彤
- XU Manhou, LIU Min, ZHAI Datong, LIU Tong.
- 植物种间联结研究内容与方法评述
- A review of contents and methods used to analyze various aspects of plant interspecific associations
- 生态学报[J]. 2016, 36(24): 8224-8233
- Acta Ecologica Sinica[J]. 2016, 36(24): 8224-8233
- http://dx.doi.org/10.5846/stxb201505311092
-
文章历史
- 收稿日期: 2015-05-31
- 网络出版日期: 2016-04-12





2. 石河子大学, 生命科学学院, 石河子 832003
2. Shihezi University, College of Life Sciences, Shihezi 832003, China
植物群落中各种群间的相互作用关系既有互助性[1],也有对抗性[2],它反映了种群在群落中的地位[3]。种间联结是不同物种在空间分布上的相互关联性,它是植物群落重要的数量和结构特征之一,是群落结构形成和演化的重要基础[4]。植物种间联结性的研究,可以确定植物的种间关系,预测种群的消长动态,揭示群落演替中植物替代关系的机制,对植被的恢复和重建、环境和物种多样性的保护、退化生态系统的防治及其相关政策法规的实施等具有重要的理论意义和应用价值[5]。
种间联结有正联结和负联结之分。种间联结理论认为,正联结是:(1)由于一个物种依赖于另一个物种;(2)由于在异质的环境内几个物种对环境条件有相似的适应和反应;负联结是:(1)二者在竞争资源中互相排斥;(2)环境需求的不相似性。在同一环境条件下,正联结可能在某种程度上指示相互作用的存在对一方或双方种是有利的,例如互惠共生或资源划分方面的互补;负联结可能表明不利于一方或双方的相互作用,例如种间竞争、干扰。20世纪以来,关于植物群落种间关系的研究屡有报道。在我国,植物种间联结研究始于20世纪70年代末,主要以二元数据为主;近年来,种间联结趋向于采用定量数据进行研究。研究对象涉及到山地[5-6]、草地[7]、湿地[8]、迹地[9]、荒漠[10]等群落,对高寒地区植物群落种间联结性的研究却较少报道。在国外,种间联结理论已被广泛应用于生态学的许多领域,常见的如优势种群的关联性分析[11-12]、群落的动态研究[13-15]等。然而,种间联结是对物种关系外在现象的描述,展现的是种间竞争的结果或群落的现状,不能揭示其过程。为了便于探讨群落演替中植物替代关系的内在机制,本文对种间联结的研究内容、计算方法及其影响因素进行了评述,并指出了现研究存在的问题及今后的研究方向。
1 种间联结的研究内容 1.1 利他作用从干旱-半干旱灌丛地到稀树大草原、湿地、沼泽、沙漠等,植物的“利他作用(Facilitation)”都存在,且随着环境恶劣程度的增加而增强。储诚进课题组[16-18]研究认为,植物可通过竞争水分和光照对林下植被产生伤害,但当水分的增加和可用营养的提高超过对水分和光照的竞争时,“利他作用”便产生了。灌丛植物的树冠对降雨截留形成的灌丛雨和茎流含有丰富的矿质营养,再加之冠层的残枝落叶,从而增强植冠下土壤的肥力,进而为其它定居物种的种子萌发与幼苗建成提供充足的养分和水分。Benard和Toft[1]通过实验发现,在灌丛植冠下萌发的幼苗比灌丛间地萌发的幼苗长得大且存活率高,因为灌丛植物可以利用其树冠为幼苗提供荫凉处,减弱强烈的光照,降低土壤表面的温度,减少土壤水分的蒸发,从而促进幼苗的萌发和存活。可见,植物“利他作用”体现了重要的种间联结性。
1.2 生物入侵生物入侵是全球生物多样性丧失的重要原因,是21世纪全球最棘手的生态问题之一,已备受国际社会的广泛关注。西方国家对入侵种研究较多,内容包括普遍性的入侵路径、载体、自然和人工干扰的作用,以及入侵种带来的危害等等。国内对入侵种的调查及防除的研究也有较多报道,入侵植物方面大多是对薇甘菊(Mikania micrantha)[19]、紫茎泽兰(Eupatorium adenophorum)[20]、凤眼蓝(Eichhornia crassipes)[21]、空心莲子草(Alternanthera philoxeroides)[22]、胜红蓟(Ageratum conyzoides)[23]等物种的入侵性和危害性进行研究,而对入侵种与周围物种和环境关系的研究相对较少,还需加强入侵种种间联结的研究。
1.3 生态种组生态种组是群落生态习性相似的种的联合,能够更真实地反映群落和种群的关系以及种群对环境和主导生态因素的适应方式。现主要有两种方法划分生态种组:(1)根据种间联结和生态习性划分生态种组。杨兆静等[24]按照相关性关系,根据秩相关系数检验结果,并结合物种各自的生态习性,将暴马丁香(Syringa reticulata)群落的34个优势种划分为两大生态种组。(2)利用聚类与排序方法划分生态种组。CCA(Canonical Correspondence Analysis)是当今生态学应用最广泛的直接梯度排序方法[25]。在进行CCA排序前要对次数据集的环境变量进行标准化,然后选用Correlation距离系数,利用PC-ORD组平均法对样地内的所有种群进行聚类[26]。聚类分析与CCA排序共同揭示环境因子影响物种的分布,并通过生态习性表达出来。因此,以排序和聚类方法为手段,结合植物种的生态习性及种间联结的相关分析,更能准确地划分生态种组。
1.4 不同演替阶段的种间关系群落种间总体关联性及其变化规律反映了群落演替的阶段和群落的稳定性。然而,当群落逐渐向着顶极群落发展时,种间联结性如何发生改变,不同学者说法不一。黄世能和王伯荪[27]认为,随着演替进展群落中的物种总体向无关联发展;而张明霞等[28]认为,种对间正联结比例越高,群落结构越趋于稳定。导致结论不一致的原因可能与生态系统类型、群落类型、群落所处的演替阶段、取样面积以及样方数量等有关。一般而言,随着植被群落演替的进展,群落结构及其种类组成将逐渐趋于完善和稳定,种间关系也将逐步趋向于正相关,以求得物种间的稳定共存。但事实上,群落种间联结性在群落演替的不同阶段有所不同。群落发育初期,物种间趋向于随机性,未形成特定的种间关系;随着演替进展到中期,群落中物种间的关系随着种间竞争的不断加剧而发生复杂变化,表现较强的正联结和负联结。
1.5 不同尺度格局的种间关系自Watt[29]《植物群落格局与过程》一文发表之后,有关植被格局的观点逐渐统一,绝大部分现代格局研究方法均受Watt观点的影响。其基本思想是“自然界的许多植物种在不同尺度上是以斑块镶嵌式存在的,并且这种斑块有一定的可预测性,可以进行定量描述”,即空间格局。空间格局是自然植被研究的重要方面,对植物种群间的联结关系产生重要影响,是现代植物生态学研究的主题之一[30]。物种间的联结关系会由于其它种的出现或消失而改变[31],而多种群格局能更真实地反映植被空间结构的实质。由Ver Hoef和Glenn-Lewin[32]修正的多尺度排序法,能有效地鉴别出多种群多尺度的空间格局,并能分析不同尺度格局上复杂的种间联结关系,因而成为国内外研究多种群多尺度格局应用频率最高的方法之一。
2 种间联结的计算方法虽然人们很早就注意到植物间的相关现象,但对其定量研究是随着本世纪初概率论与数理统计学科的发展和种间联结测定技术在植物生态学、植被分析中获得广泛应用而发展起来的。种间联结的测定方法很多,它们都各具特色,究竟哪一种方法是最合适的,至今仍没有定论。现将使用较多的方法作归纳。
2.1 多物种种间联结显著性检验1984年,Schluter[33]提出方差比率(VR)法测定多物种间的联结性,并用统计量w来检验关联程度,其计算公式如下:
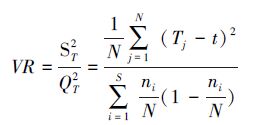
|
(1) |
式中,s为待测种群总数,N为总样方数,ni为种群i出现的样方数,Tj为样方j内出现的种群数,t为样方中种群的平均数,t=(t1+t2+…+tN)/N。
VR值作为全部物种的关联指数,在独立性零假设条件下,期望值为1。若VR>1,则表明种间表现出净的正联结;若 VR<1,则表明种间表现出净的负联结;若 VR=1,则符合所有种间无联结的零假设。对测定结果还需采用统计量 w=N×VR来检验VR值偏离1的显著程度,若物种不显著相关联,则 w 落入由下面x2分布给出的界限的概率为90%:χ0.952(N)<w<χ0.052(N),否则,总体相关。该方法用于判定群落中各主要种群是否存在显著的联结性,可以同时测定多个物种间的关联程度。
2.2 成对物种种间联结测度指标2×2联列表是种间联结测定的基础,通常是先把要测定的成对物种在取样中的存在与不存在数据排列成2 × 2联列表,再将联列表内的数据代入有关数学公式进行计算,最后根据计算结果来分析确定成对物种间的联结程度。
2.2.1 x2检验统计量该方法以定性数据为基础,从表层角度揭示出种对联结的性质和显著程度,检验统计量按下式计算[34]:
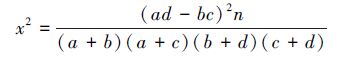
|
(2) |
式中,a为两个物种都出现的样方数,b、c为只出现一个物种的样方数,d为两个物种都不出现的样方数。
x2值在以下情况下被认为有偏差:(1)2×2列联表中任一小格期望值小于1;(2)多于两个小格的期望值小于5。这种偏差可用Yates连续校正系数来纠正[35]:
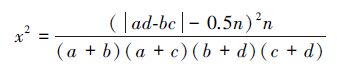
|
(3) |
若x2<3.841(P>0.05),表示种对间无联结性,两种间基本独立;若3.841≤x2<6.635(0.01<P≤0.05),表示种对间联结性显著;若x2≥6.635(P≤0.01),表示种对间联结性极显著。
x2检验以二元数据作为联结性判断的标准,这样会丧失种间相关的多度信息,且x2检验只是对种间联结性作出了一个概括性的结论,只能作定性判断,不能区分联结强度的大小。但因其分析较简单,有明确的指标,能比较准确客观地反映出种间联结性。
2.2.2 Fisher精确检验统计量首先将原始数据矩阵转化为(0,1)形式的二元数据矩阵,然后分别构造物种的2 × 2列联表,对这些列联表分别进行 Fisher精确检验[36],其表达式如下:
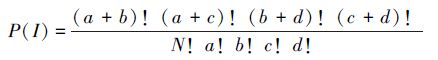
|
(4) |
对于任一2 × 2列表,精确检验首先按公式求出p(1),然后在保持行、列总数(a+b,c+d,a+c,b+d)不变的前提下,将表中最小的值逐个降低直到为0,每降低1次,按公式求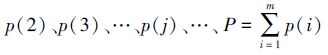
Fisher精确检验也有其自身的局限性。在植物群落当中,如果某一物种在所有样方中都存在,这时就无法进行Fisher精确检验以了解该物种与其它物种之间的相互关系,但这并不表明该物种与其它物种之间不存在关联。因此,在精确检验的基础上,结合相关系数和秩相关系数来检验种对间的相关性,可以揭示种间的关联关系。
2.2.3 相关系数和秩相关系数相关系数的测定是定性、定量数据相结合的方法。Pearson相关系数[37]如下:
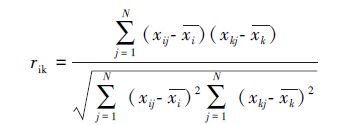
|
(5) |
式中,N为样方数目;xij和xkj分别是种i和种k在样方j中的多度值,它们分别组成两个向量xi和xk;xi和xk分别是种i和种k在所有样方中多度的平均值。rik值域为[-1,1],正值表示正相关,负值表示负相关。
秩相关系数是将多度向量变换为秩化向量,然后将秩化向量代入相关系数公式得到的。Spearman秩相关系数[37]如下:

|
(6) |
式中,N为样方总数;xij和xkj分别是种i和种k在样方j中的秩。
x2检验只能作定性描述,不能很好地反映两个变量在数量上的变化趋势,而相关系数和秩相关系数基于数量数据,相关系数检验对x2检验可以进行有效地补充和完善。Pearson相关分析要求物种服从正态分布,但自然界大多数物种服从集群分布,就使Pearson相关分析有一定的局限性;而Spearman秩相关分析属于非参数检验,对物种的分布形式不做要求,而且比Pearson相关系数检验灵敏度高,因此,可以弥补相关系数检验的不足。
2.2.4 联结系数和匹配系数(1) 联结系数(AC)[38]
若ad≥bc,则
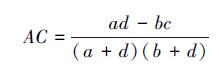
|
(7) |
若bc>ad且d≥a,则
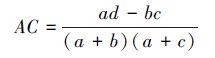
|
(8) |
若bc>ad且d<a,则
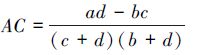
|
(9) |
由于AC计算公式中分子为 (ad - bc),当所研究群落中物种数目较多而调查样方面积较小时,仅能体现较少的物种数,这样一方面会人为地减小了b或c;另一方面也可能增加d,从而导致AC正值偏高。当ad > bc时为正联结,ad < bc时为负联结,ad = bc时无联结。
(2) 匹配系数[39]
Ⅰ Jaccard系数(JI):
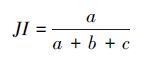
|
(10) |
Ⅱ Dice系数(DI):
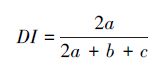
|
(11) |
Ⅲ Ochiai系数(OI):
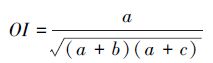
|
(12) |
JI、DI、OI 三个指数都表示种对相伴随出现的机率和联结性程度,当a=0时,取值为0,表明种间完全相异,不同时出现在同一样方中;当a=S(总种数)时,取值1,表示同时出现在样方中。JI、DI、OI实质上是等效的,在种数较多和取样数目较大时,OI、DI、JI能避免d值高所导致的AC偏高的缺点,更好的反映出关联性不明显的物种间共同出现的机率,但OI、DI、JI仅能较准确地反映物种间的正联结性的强弱,在a较小时,会忽略d的存在,就夸大了a、b、c的作用,从而夸大了负联结性的强度。
2.2.5 点相关系数 (PCC)[40]

|
(13) |
PCC计算结果与AC值相接近,一般小于AC。AC、JI、DI、OI能体现出那些由x2检验证明不显著的联结性,且能反映种间联结性的相对强弱,但对强弱的等级划分缺少统一的标准,并且在物种出现次数少,特别是当a较小时,会忽略d,从而夸大a、b、c在种间联结测定中的作用,甚至会得出不同结论。而PCC能降低上述情况对联结性分析的影响。
2.3 用样方中出现的个体数测算联结值[41]
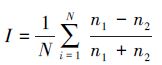
|
(14) |
式中,I为联结值,N为样方数,n1和n2分别代表两物种在某一样方中出现的个体数。
I值取值区间为[0,1],I值越大,联结关系越小;反之,联结关系越大。此方法有助于解释一些特例,但它没有什么分级标准,且受取样面积的影响较大。
3 种间联结的影响因素群落物种间的联结性受多方面因素影响,既有人为取样尺度的影响,又有环境与生物自身的因素。生物本身特性(如竞争、化感作用)和特殊事件(如虫灾、灾难性天气)都会对种间联结性产生影响。
3.1 人为取样因素尺度对植物群落种间关系格局的形成具有决定性作用,在较大尺度内,环境异质性对种间关联起主要作用;而在较小尺度内,种间竞争则起主导作用[42]。目前计算种间联结的方法大多基于2 × 2列联表,所以其测定的精确性在很大程度上取决于样方的面积和数目。取样过大,种间关系多为正联结;取样过小则不能包含个体较大的两个物种,容易形成负联结。对南亚热带常绿阔叶林种间测定研究表明,在南亚热带取样面积以100 m2为宜,取样数目以30—50个为宜[38]。如果在研究取样过程中种群内不同种对间的正负联结数相差不大,计算得到的正联结种对数占总种对的比例与负联结种对所占的比例的比值为1,或趋近于1,这就说明取样尺度合理有效,此次取样能够准确反映调查地的植被分布状况。
另外,在决定样方大小时,首先要考虑研究的群落类型、优势种的生活型及植被的均匀性等。从统计学上讲,使用面积小而数目多或者面积大数目少的样方可以达到同样的精确度,但样方小,取样工作量增加,计算也麻烦,同时许多样方的观测值可能很接近,给数量分析带来一定困难。所以,样方大小要适当,一般用群落的最小面积作为样方的大小。表 1是一些植被类型群落最小面积的经验值。
| 群落类型 Community types | 群落最小面积/m2 Minimal area of community | 群落类型 Community types | 群落最小面积/m2 Minimal area of community |
| 地衣群落Lichen community | 0.1—0.4 | 苔藓群落Moss community | 1—4 |
| 沙丘草原Sand dunes grassland | 1—10 | 干草原Steppe | 1—25 |
| 草甸Meadow | 1—50 | 高草地Prairie land | 5—50 |
| 灌丛Bushwood | 10—50 | 温带森林Temperate forest | 200—500 |
| 热带雨林Tropical rainforest | 500—4000 |
(1) 种间竞争
自然群落中植物竞争是决定群落结构、物种丰富度、物种多样性和群落动态的动力之一。Grime的 C-S-R 生活史模型和Tilman的资源比例模型虽然被广泛验证,但是并没有完全揭示自然群落中植物竞争的机制[43]。在计算指标上,以百分数形式表示的相对生长速率和竞争强度被广泛用来反映和衡量植物对种间关系的响应和植物之间相互作用的强度,即当竞争强度大于0,植物之间存在竞争;当竞争强度小于0,植物与其它物种共存,生长得到促进[44]。李莉等[45]采用去除试验研究了混播草地群落的种间竞争与共存关系,结果表明永久性混播草地群落中,豆科牧草与禾草之间竞争不对等而实现竞争共存,豆科、禾草之间的不对称竞争是白三叶长期维持合理比例和混播组分之间动态平衡的重要机制。可见,植物竞争特性对种间联结性的影响可能与物种对群落的选择和群落对生存物种的选择与淘汰有密切关系。
(2) 化感作用
植物之间的化学相互作用关系——化感作用,是化学生态学研究的重要方面。近年来,对植物化感作用的研究越来越多,成为当今植物保护领域研究的热点。任元丁等[46]综合了我国主要草地类型化感作用的现有研究成果,对已报道的化感植物、化感物质和化感物质释放途径的研究进行了综述。然而,我国草地植物化感作用的研究由于起步较晚、时间不足等因素,取得的成果仍然很少。多数研究的焦点还集中在对化感植物的分类以及化感物质的提取、分离和鉴定等方面,对于植物化感作用的机制与应用以及化感作用下的种间联结性等方面的研究还不是很多[47-51]。因此,在今后的研究中,应加强对草地上具有化感潜势的植物种的调查评价和利用研究,深入探索化感植物在草地退化进程中的种间联结性和演变规律。
3.3 特殊环境因素虫害等一些特殊事件可对植物种间关系产生影响。虫害爆发属于一种自然干扰,虫灾的爆发对群落的结构和组成变化产生很大影响。植被的变化引起昆虫种群数量的动态变化,而昆虫种群数量的动态变化又影响植被的结构组成,它们之间互为反馈[52]。另外,一些极端气候事件(如南方灾害性天气)造成的第一层高大乔木死亡亦对群落结构造成影响。当灾害性天气(如雷暴)造成第一层高大乔木死亡导致了林窗的产生,这也促进群落其它阳生性树种的发展。但随着林窗的恢复,一部分阳生性树种被淘汰,中生性树种又开始发展。同时,台风也加速了老弱病残树种的死亡,在一定程度上加速了群落的循环演替过程,造成种间关系变化及群落结构波动[52]。因此,特殊环境在植物种间联结中所起的作用也是不容忽视的。
4 种间联结存在的问题及今后的研究方向 4.1 存在的问题自Forbes首次通过两物种间出现的频度定量开展种间联结测定以来,种间联结的测定与分析一直为生态学家尤其是群落生态学家所青睐。但Forbes仅仅提供了一个松散的种间联结定义,而且其后诸多的种间联结研究结论互相矛盾,以致有众多的修正公式出现。因此,种间联结有其自身的局限性:
(1) 测定结果受样方面积大小和样方数目多少的影响很大;
(2) 仅能测定群落中两个物种间的相互关系;
(3) 是对种群之间关系的外在现象的描述,反映的是种间竞争的结果或群落的现状,不能揭示其过程。
4.2 今后的研究方向今后可在以下几个方面加深对种间联结性的研究,探讨其形成机制。
(1) 利用生态位理论研究种间联结,揭示生态位与种间联结的关系。
物种的联结性与其生态位有着密切联系:植物种对的正联结体现了植物利用资源的相似性和生态位的重叠性;负联结体现了植物间的排斥性,也是生态位分离的反映。生态位是一个复杂、抽象的概念,目前大家普遍接受 Hutchinson提出的n维生态位[41]。通常人们只选择几个甚至一个重要因子进行生态位分析,根据所选择的因子不同,得到的生态位结果也不一致,而种间联结的结果仅可能与“空间分布”生态位结果存在相关性。为合理解释种间联结,李丹雄等[9]认为,在固定样地中监测生态因子,并根据生态因子计算生态位,然后讨论生态位与种间联结的关系,有助于揭示种间联结的内在机制。
(2) 研究密度主导的种间联结的变化,阐释生境异质性与种间联结的关系。
到目前为止,有关群落中种间联结关系发生的具体机制是不清晰的。种群内部物种间的联结关系机制可能有密度决定的因素,而不同种群的种间联结关系则包括对生态资源的竞争和不同物种通过改变生境造成的相互影响,这些机制可否通过定量化来确定,如何确定,如何设计实验或取样进行定量研究,这些都是值得探讨的问题。此外,种群内密度主导的种间联结关系变化定量研究,在边缘生态系统中用数量模型模拟种间联结关系变化,以及对生境异质性与种间联结关系变化相互作用进行探讨等方面[53],也都是种间联结研究中应予以特别关注的问题。
(3) 从植物生理、分子遗传的角度研究种间联结,探讨种间联结产生的机制。
种间联结的进一步深入研究,仅用样方调查数据进行数学分析是远远不够的。在运用常规的群落调查方法对植物个体或种群本身进行长期定位研究的基础上,可以结合定量的实验生态学方法,如应用植物生理和分子生物学手段开展光合作用的种间差异[54-55]及种间基因序列比较的研究[56-57]。在种间基因序列比较研究方面,可以利用SSR(Simple Sequence Repeats)标记技术和RAPD(Random Amplified Polymorphic DNA)等技术探究植物种群各物种的亲缘关系[58-62],从遗传学角度确定植物的种间联结关系。综合应用多种检测方法进行遗传分析,全面准确地评价遗传差异,才能进一步揭示物种间联结关系的产生机制。
致谢: 感谢中山大学储诚进教授对工作的帮助。| [1] | Benard R B, Toft C A. Fine-scale spatial heterogeneity and seed size determine early seedling survival in a desert perennial shrub (Ericameria nauseosa: Asteraceae). Plant Ecology , 2008, 194 (2) : 195–205. |
| [2] | Jing J Y, Bezemer T M, van der Putten W H. Interspecific competition of early successional plant species in ex-arable fields as influenced by plant-soil feedback. Basic and Applied Ecology , 2015, 16 (2) : 112–119. DOI:10.1016/j.baae.2015.01.001 |
| [3] | Su S J, Liu J F, He Z S, Zheng S Q, Hong W, Xu D W. Ecological species groups and interspecific association of dominant tree species in Daiyun Mountain national nature reserve. Journal of Mountain Science , 2015, 12 (3) : 637–646. DOI:10.1007/s11629-013-2935-7 |
| [4] | Álvarez-YépizJ C, BúrquezA, DovčiakM. Ontogenetic shifts in plant-plant interactions in a rare cycad within angiosperm communities. Oecologia , 2014, 175 (2) : 725–735. |
| [5] | 张悦, 郭利平, 易雪梅, 曹伟, 王远遐, 吴培莉, 姬兰柱. 长白山北坡3个森林群落主要树种种间联结性. 生态学报 , 2015, 35 (1) : 106–115. |
| [6] | 黄祥童, 王绍先, 黄炳军, 尹航, 崔凯峰, 赵伟, 范宇光, 顾德峰. 珍稀植物对开蕨与其伴生物种的联结性及群落稳定性. 生态学报 , 2015, 35 (1) : 80–90. |
| [7] | 谢开云, 赵云, 李向林, 何峰, 万里强, 王丹, 韩冬梅. 豆-禾混播草地种间关系研究进展. 草业学报 , 2013, 22 (3) : 284–296. |
| [8] | 程雷星, 陈克龙, 杨仕兵, 苏旭, 王记明. 青海湖流域小泊湖湿地植物种间关系研究. 干旱区地理 , 2014, 37 (5) : 1005–1011. |
| [9] | 李丹雄, 杨建英, 史常青, 周进, 王兴宇, 王新, 伏凯, 赵廷宁. 汶川地震滑坡迹地植被恢复中优势种的种间关联性. 应用与环境生物学报 , 2014, 20 (5) : 938–943. |
| [10] | 张雪妮, 吕光辉, 杨晓东, 贡璐, 秦璐, 何学敏, 刘昊奇. 基于盐分梯度的荒漠植物多样性与群落、种间联接响应. 生态学报 , 2013, 33 (18) : 5714–5722. |
| [11] | Haugaasen T, Peres C A. Interspecific primate associations in Amazonian flooded and unflooded forests. Primates , 2009, 50 (3) : 239–251. DOI:10.1007/s10329-009-0135-4 |
| [12] | Guo L J, Zhang W H, Liu G B. Species diversity and interspecific association in development sequence of Hippophae rhamnoides plantations in the Loess Hilly Region, China. Frontiers of Biology in China , 2008, 3 (4) : 489–495. DOI:10.1007/s11515-008-0070-6 |
| [13] | Gutiérrez-Girón A, Gavilán R G. Spatial patterns and interspecific relations analysis help to better understand species distribution patterns in a Mediterranean high mountain grassland. Plant Ecology , 2010, 210 (1) : 137–151. DOI:10.1007/s11258-010-9745-6 |
| [14] | Li Y D, Xu H, Chen D X, Luo T S, Mo J H, Luo W, Chen H Q, Jiang Z L. Division of ecological species groups and functional groups based on interspecific association-a case study of the tree layer in the tropical lowland rainforest of Jianfenling in Hainan Island, China. Frontiers of Forestry in China , 2008, 3 (4) : 407–415. DOI:10.1007/s11461-008-0049-0 |
| [15] | Danieli-Silva A, Uhlmann A, Vicente-Silva J, Stürmer S L. How mycorrhizal associations and plant density influence intra- and inter-specific competition in two tropical tree species: Cabralea canjerana (Vell.) Mart. and Lafoensia pacari A. St. -Hil. Plant and Soil , 2010, 330 (1/2) : 185–193. |
| [16] | Wang Y S, Ellwood M D F, Maestre F T, Yang Z Y, Wang G, Chu C J. Positive interactions can produce species-rich communities and increase species turnover through time. Journal of Plant Ecology , 2012, 5 (4) : 417–421. DOI:10.1093/jpe/rts005 |
| [17] | Chu C J, Weiner J, Maestre F, Xiao S, Wang Y S, Li Q, Yuan J L, Zhao L Q, Ren Z W, Wang G. Positive interactions can increase size inequality in plant populations. Journal of Ecology , 2009, 97 (6) : 1401–1407. DOI:10.1111/jec.2009.97.issue-6 |
| [18] | Chu C J, Maestre F, Xiao S, Weiner J, Wang Y S, Duan Z H, Wang G. Balance between facilitation and resource competition determines biomass-density relationships in plant populations. Ecology Letters , 2008, 11 (11) : 1189–1197. |
| [19] | 李鸣光, 鲁尔贝, 郭强, 昝启杰, 韦萍萍, 蒋露, 徐华林, 钟填奎. 入侵种薇甘菊防治措施及策略评估. 生态学报 , 2012, 32 (10) : 3240–3251. |
| [20] | 张丽坤, 王朔, 冯玉龙. 紫茎泽兰种子形态特征和萌发特性与其入侵性的关系. 生态学报 , 2014, 34 (13) : 3584–3591. |
| [21] | 周晴, 潘晓云. 中国南部基塘区农业模式的变迁与凤眼蓝的入侵. 植物生态学报 , 2014, 38 (10) : 1093–1098. |
| [22] | 褚延梅, 杨健, 李景吉, 彭培好. 三种增温情景对入侵植物空心莲子草形态可塑性的影响. 生态学报 , 2014, 34 (6) : 1411–1417. |
| [23] | 周兵, 闫小红, 肖宜安, 王宁, 旷志强. 不同生境下入侵植物胜红蓟种群构件生物量分配特性. 生态学报 , 2015, 35 (8) : 2602–2608. |
| [24] | 杨兆静, 张钦弟, 李豪, 白玉宏, 毕润成. 山西五鹿山自然保护区暴马丁香群落木本植物种间联结性分析. 植物科学学报 , 2013, 31 (1) : 42–48. |
| [25] | Liang J, Hua S S, Zeng G M, Yuan Y J, Lai X, Li X D, Li F, Wu H P, Huang L, Yu X. Application of weight method based on canonical correspondence analysis for assessment of Anatidae habitat suitability: A case study in East Dongting Lake, Middle China. Ecological Engineering , 2015, 77 : 119–126. DOI:10.1016/j.ecoleng.2015.01.016 |
| [26] | Jin Z J, Li Z Y, Li Q, Hu Q J, Yang R M, Tang H F, Li M, Huang B F, Zhang J Y, Li G W. Canonical correspondence analysis of soil heavy metal pollution, microflora and enzyme activities in the Pb-Zn mine tailing dam collapse area of Sidi village, SW China. Environmental Earth Sciences , 2015, 73 (1) : 267–274. DOI:10.1007/s12665-014-3421-4 |
| [27] | 黄世能, 李意德, 骆土寿, 王伯荪. 海南岛尖峰岭次生热带山地雨林树种间的联结动态. 植物生态学报 , 2000, 24 (5) : 569–574. |
| [28] | 张明霞, 王得祥, 康冰, 张岗岗, 刘璞, 杜焰玲, 于飞. 秦岭华山松天然次生林优势种群的种间联结性. 林业科学 , 2015, 51 (1) : 12–21. |
| [29] | Watt A S. Pattern and process in the plant community. Journal of Ecology , 1947, 35 (1/2) : 1–22. DOI:10.2307/2256497 |
| [30] | Hendriks M, Ravenek J M, Smit-Tiekstra A E, van der Paauw J W, de Caluwe H, van der Putten W H, de Kroon H, Mommer L. Spatial heterogeneity of plant-soil feedback affects root interactions and interspecific competition. New Phytologist , 2015, 207 (3) : 830–840. DOI:10.1111/nph.13394 |
| [31] | García-Cervigón A I, Gazol A, Sanz V, Camarero J J, Olano J M. Intraspecific competition replaces interspecific facilitation as abiotic stress decreases: The shifting nature of plant-plant interactions. Perspectives in Plant Ecology, Evolution and Systematics , 2013, 15 (4) : 226–236. DOI:10.1016/j.ppees.2013.04.001 |
| [32] | Ver Hoef J M, Glenn-Lewin D C. Multiscale ordination: a method for detecting pattern at several seales. Vegetatio , 1989, 82 (1) : 59–67. |
| [33] | Schluter D. A variance test for detecting species associations, with some example application. Ecology , 1984, 65 (3) : 998–1005. DOI:10.2307/1938071 |
| [34] | Kichenin E, Wardle D A, Peltzer D A, Morse C W, Freschet G T. Contrasting effects of plant inter- and intraspecific variation on community-level trait measures along an environmental gradient. Functional Ecology , 2013, 27 (5) : 1254–1261. DOI:10.1111/fec.2013.27.issue-5 |
| [35] | Li J, Zhang X Y, Tian C M. Spatial pattern of paulownia witches' broom and its effects on the growth of Paulownia elongata in Shaanxi Province, China. Forestry Studies in China , 2009, 11 (1) : 34–38. |
| [36] | 吴仲贤. 生物统计学. 北京: 中国农业大学出版社, 1992393–395. |
| [37] | Bishara A J, Hittner J B. Testing the significance of a correlation with nonnormal data: comparison of Pearson, Spearman, transformation, and resampling approaches. Psychological Methods , 2012, 17 (3) : 399–417. DOI:10.1037/a0028087 |
| [38] | 王伯荪, 彭少麟. 南亚热带常绿阔叶林种间联结测定技术研究I. 种间联结测式的探讨与修正. 植物生态学与地植物学丛刊 , 1985, 9 (4) : 274–285. |
| [39] | Cárdenas R E, Valencia R, Kraft N J B, Argoti A, Dangles O. Plant traits predict inter-and intraspecific variation in susceptibility to herbivory in a hyperdiverse neotropical rain forest tree community. Journal of Ecology , 2014, 102 (4) : 939–952. DOI:10.1111/1365-2745.12255 |
| [40] | 吴大荣. 一种新的植物种间联结测定公式的探讨. 东北林业大学学报 , 1997, 25 (5) : 74–80. |
| [41] | Gehring C, Flores-Rentería D, Sthultz C M, Leonard T M, Flores-Rentería L, Whipple A V, Whitham T G. Plant genetics and interspecific competitive interactions determine ectomycorrhizal fungal community responses to climate change. Molecular Ecology , 2014, 23 (6) : 1379–1391. DOI:10.1111/mec.12503 |
| [42] | Müller B, Bartelheimer M. Interspecific competition in Arabidopsis thaliana: root hairs are important for competitive effect, but not for competitive response. Plant and Soil , 2013, 371 (1/2) : 167–177. |
| [43] | Connolly J, Wayne P, Bazzaz F A. Interspecific competition in plants: how well do current methods answer fundamental questions?. The American Naturalist , 2001, 157 (2) : 107–125. DOI:10.1086/318631 |
| [44] | Kim T N, Underwood N, Inouye B D. Insect herbivores change the outcome of plant competition through both inter- and intraspecific processes. Ecology , 2013, 94 (8) : 1753–1763. DOI:10.1890/12-1261.1 |
| [45] | 李莉, 王元素, 王堃. 喀斯特地区永久性禾草+白三叶混播草地群落种间竞争与共存. 草业科学 , 2014, 31 (10) : 1943–1950. |
| [46] | 任元丁, 尚占环, 龙瑞军. 中国草地生态系统中的化感作用研究进展. 草业科学 , 2014, 31 (5) : 993–1002. |
| [47] | 闫志强, 宋本如, 刘黾, 刘燕, 吴小业, 李洁珊, 林燕珍, 冯翊果, 陈章和, 赵建刚. 5种沉水植物对斜生栅藻的化感作用. 应用与环境生物学报 , 2015, 21 (1) : 75–79. |
| [48] | 马丹炜, 王亚男, 王煜, 张红, 廖颖, 何兵. 化感胁迫诱导植物细胞损伤研究进展. 生态学报 , 2015, 35 (5) : 1640–1645. |
| [49] | 谢星光, 陈晏, 卜元卿, 戴传超. 酚酸类物质的化感作用研究进展. 生态学报 , 2014, 34 (22) : 6417–6428. |
| [50] | 陈娟, 白尚斌, 周国模, 王懿祥, 梁倩倩, 程艳艳, 沈蕊. 毛竹浸提液对苦槠幼苗生长的化感效应. 生态学报 , 2014, 34 (16) : 4499–4507. |
| [51] | 侯玉平, 柳林, 王信, 闫晓宇, 门航, 李伟杰, 徐维明. 外来植物火炬树水浸液对土壤微生态系统的化感作用. 生态学报 , 2013, 33 (13) : 4041–4049. |
| [52] | 彭李菁. 鼎湖山气候顶极群落种间联结变化. 生态学报 , 2006, 26 (11) : 3732–3739. |
| [53] | Dissanayaka D M S B, Maruyama H, Masuda G, Wasaki J. Interspecific facilitation of P acquisition in intercropping of maize with white lupin in two contrasting soils as influenced by different rates and forms of P supply. Plant and Soil , 2015, 390 (1/2) : 223–236. |
| [54] | Hikosaka K. Mechanisms underlying interspecific variation in photosynthetic capacity across wild plant species. Plant Biotechnology , 2010, 27 (3) : 223–229. DOI:10.5511/plantbiotechnology.27.223 |
| [55] | Sang Y R, Wang C K, Huo H. Inter-specific and seasonal variations in photosynthetic capacity and water use efficiency of five temperate tree species in Northeastern China. Scandinavian Journal of Forest Research , 2011, 26 (1) : 21–29. DOI:10.1080/02827581.2010.534499 |
| [56] | Zhang Y S, Hou B W, Zhang W M, Ding X Y. Isolation and characterization of novel microsatellite markers for Bletilla striata and inter-specific amplification in 2 congeneric species. Conservation Genetics Resources , 2015, 7 (2) : 483–485. DOI:10.1007/s12686-014-0401-7 |
| [57] | González V M, Aventín N, Centeno E, Puigdomènech P. Interspecific and intraspecific gene variability in a 1-Mb region containing the highest density of NBS-LRR genes found in the melon genome. BMC Genomics , 2014, 15 : 1131–1131. DOI:10.1186/1471-2164-15-1131 |
| [58] | 左力辉, 张文林, 邱彤, 张军, 杨敏生. 新疆野苹果叶形性状变异及其与SSR标记关联分析. 园艺学报 , 2015, 42 (4) : 759–768. |
| [59] | 陆欢, 张丹, 章炉军, 王瑞娟, 尚晓冬, 谭琦. 金针菇种质资源5个农艺性状与SSR标记的关联分析. 农业生物技术学报 , 2015, 23 (1) : 96–106. |
| [60] | 郭传友, 王贝贝, 吕岩桢, 张兴旺. 基于RAPD标记的皖南山核桃野生居群遗传多样性分析. 生态学杂志 , 2015, 34 (5) : 1301–1306. |
| [61] | Fajardo C G, de Almeida Vieira F, Molina W F. Interspecific genetic analysis of orchids in Brazil using molecular markers. Plant Systematics and Evolution , 2014, 300 (8) : 1825–1832. DOI:10.1007/s00606-014-1009-9 |
| [62] | Xia W, Xiao Y, Liu Z, Luo Y, Mason A S, Fan H K, Yang Y D, Zhao S L, Peng M. Development of gene-based simple sequence repeat markers for association analysis in Cocos nucifera. Molecular Breeding , 2014, 34 (2) : 525–535. DOI:10.1007/s11032-014-0055-x |