 2016, Vol. 36
2016, Vol. 36文章信息
- 陈文年, 王辉, 肖小君, 陈发军, 张志勇, 齐泽民, 黄作喜
- CHEN Wennian, WANG Hui, XIAO Xiaojun, CHEN Fajun, ZHANG Zhiyong, QI Zemin, HUANG Zuoxi.
- 坡向对暗紫贝母生长和繁殖特征的影响
- Effects of slope aspect on growth and reproduction of Fritillaria unibracteata (Liliaceae)
- 生态学报[J]. 2016, 36(24): 8174-8182
- Acta Ecologica Sinica[J]. 2016, 36(24): 8174-8182
- http://dx.doi.org/10.5846/stxb201505251041
-
文章历史
- 收稿日期: 2015-05-25
- 网络出版日期: 2016-04-12





2. 四川省高校特色农业资源研究与利用重点实验室, 内江 641112
2. Key Laboratory of Characteristic Agricultural Resources, Department of Education, Neijiang 641112, China
在青藏高原东部,存在着十分明显的阴阳坡现象。坡向作为主要的地形因子之一,影响着坡面上太阳辐射的吸收量,并会进一步影响诸如气温、土壤温度、土壤水分等生态因子,从而影响不同坡面植物的生长、繁殖等个体特征,甚至影响各坡面上的群落及植被类型[1-3]。
暗紫贝母(Fritillaria unibracteata Hsiao et K.C. Hsia)是青藏高原东部重要的药用植物,隶属于百合科贝母属,其野生资源主要生长于3200—4500 m的青藏高原高山环境中,分布区域比较狭窄[4-6]。在自然生境中,暗紫贝母种子的萌发率十分低下[7],种群密度较低,数量很有限。另外,暗紫贝母赖以生存的青藏高原东部地区属于生态环境脆弱带[8],再加上人们受经济利益的驱动,长期大量采挖,致使其生境不断恶化,野生资源存量已是十分低下[9-10]。因此,如何扩大暗紫贝母种群的数量、提高其产量显得十分必要,而筛选出暗紫贝母野生资源的最佳生长环境正是扩大其种群密度和资源产量的重要途径之一。那么,暗紫贝母在不同坡向的环境中生长和繁殖特征是否相同?如果不同,会表现出怎样的差异?以前的研究,很多是关于暗紫贝母的药用成份的提取和分析[11-12],还有些是关于器官(鳞茎)的组织培养[13-14],也还有部分是关于药理[15]和内生真菌[16]方面的。在宏观生态学领域,陈士林等用数学方法对暗紫贝母群落进行了分类[17],并研究了群落类型与其品质之间的关系,认为产于松潘等地的“松贝”为最佳商品规格[18]。随着研究的不断深入,各方面的工作也逐渐展开,如对暗紫贝母生物量的分配特征进行的研究[19],以及对暗紫贝母鳞茎和物候在海拔梯度上的变化趋势展开的讨论[20]。但对野生暗紫贝母个体生长和繁殖特性在同一山体不同坡面间的比较研究还少有报道。
据在研究区域内的长期观察,暗紫贝母的分布主要是受海拔高度的影响,在适宜的海拔高度上,不同朝向的坡面上都能有所生长。为此,以不同的坡向作为环境因子,揭示其对暗紫贝母生长和繁殖特性的影响,从而更好地比较不同坡向上暗紫贝母生长的优劣,为暗紫贝母人工种植中最佳生境的选择提供理论依据。
1 材料和方法 1.1 研究地概况研究地位于青藏高原东部的四川省松潘县水晶乡牛牛山(32°47′ N,103°37′ E)和章腊乡卡卡山(32°59′ N,103°40′ E)[21-22],两山相距大约45 km,研究区域年均温为2.8℃,1月均温-7.6℃,7月均温9.7 ℃,年降雨量为634.8 mm,其中的72%发生在6—8月的夏季,土壤类型为黑毡土[23],两山海拔高度均为4000 m左右。
1.2 样方设置及试验设计在牛牛山和卡卡山的东、南、西、北4个方向,各设置一个70 m×15 m(长×宽)的样地,两山的各个样地海拔高度都设置为3800 m左右,并于2010年7月开始在各个样方周围设置铁丝围栏以防人畜干扰。
暗紫贝母是多年生植物,据笔者在研究区域的育苗实践及长期野外观察,暗紫贝母在野生自然条件下,其种子在萌发的当年,地上部分仅能长出一片很细的针形叶,俗称“一根针”;到第2年,地上部分可以长出一片稍长稍宽一些的披针形叶子,俗称“一片草”;到第3年,地上部分才开始长出茎,茎上长出几片披针形的叶子,俗称“树儿子”;到第4年,地上茎上可以开花,开始有性繁殖阶段,俗称“灯笼花”。以此为标准,于2011年7月,在两山的每一个样地中随机选择30株野生的“一片草”植株(即二年生个体,2—3a后可开花结果)作为研究对象,所有样方中被选择的植株都作了标记。
1.3 测定指标 1.3.1 各样地生态因子的测量在2013、2014年的3—5月,测量两山各样地中各标记植株从积雪中融化出来的时间(以1月1日开始的序日表示);同时,在每个样地中放置5个自动温度记录仪(型号:ZDR-01)以测定3—5月的气温的变化,记录仪离地高度为50 cm,设定为每1h自动记录1次;在5月中旬(此时积雪已融化完,冻土已经解冻)到8月底,用便携式水份测量仪(型号:Mpkit-B)和pH计(型号:PHT-810)测量每个样地中的土壤含水量及土壤酸碱度,每个样地取中心和四周测量点5个,每周1次。
1.3.2 物候期的测定2013年的4—9月,测定两山各样地中选定植株的下列物候期,每3d观察1次。1)开始生长期:以萌芽冒出地面1cm为标准,记录每个萌芽达到1cm的日期,并转化为从1月1日开始计算的序日[24](以下物候期采用相同的转换方法);2)始花期:以每株的花蕾刚开始开放为标准,记录每株的花蕾刚开始开放的日期;3)盛花期:以每株的花蕾完全开放为标准,记录每株的花蕾完全开放的日期;4)植株枯黄期:以每个个体叶片枯黄一半时的日期为标准,记录每株叶片枯黄达到一半的日期。
1.3.3 叶特征的测定2014年7月中下旬,正值暗紫贝母的生长期,把各样地中每株被选中的植株上最大的一片叶子摘下,用叶面积测量仪(美国CID公司,型号CI-203)测量其叶面积的大小。待测定完叶面积后,把每片叶作好编号,装入信封袋中在恒温箱中在60℃下烘48h[25],用万分之一电子天平(型号:FA1004)称量其各自的重量,并计算比叶面积(specific leaf area),比叶面积(SLA)的计算公式为[25]:
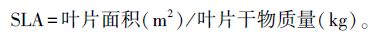
|
2014年8月下旬,在果实和种子的成熟期,测量各样地中所有标记30株植株的高度。同时,用收割法将各样地中各标记植株平地割起,分别装在不同的信封袋中,作好编号。将装有植株的信封放入恒温箱中60℃烘48h,也用万分之一电子天平将各植株的地上部分进行称量、记数,将测定叶面积后的那片叶的干重加入各植株的地上部分中,以计算各植株地上部分的总生物量。然后再挖起地下鳞茎,小心清洗,也装入编了号的信封袋中,用和测量地上部分生物量相同的方法测定这些鳞茎各自的生物量。
1.3.5 果实和种子的测定同样在2014年8月下旬,在测定完植株的地上部分的生物量之后,从这些植株上轻轻剪下果实,称量其干重,并从果实中取出种子,记录种子的个数,同时称量每个果实中的种子干重,并进一步推算各自千粒重。
1.4 统计方法用SPSS17.0对不同坡向间生态因子、叶特征、植株和鳞茎特征、果实和种子特征等方面进行方差分析(one-way ANOVA)和多重比较,所得数据以“平均数±标准误”表述。
2 结果 2.1 各部位生态因子的比较如表 1所示,不管是在牛牛山还是在卡卡山,积雪融化完的时间以北坡最迟而南坡最早,南北坡向之间差异最大,达到两个星期左右,东面坡向和西面坡向的融雪时间介于南坡和北坡的融雪时间之间。从3—5月的平均气温来看,南坡最高,而北坡最低,二者相差达3.4℃(牛牛山)和2.6℃(卡卡山),东坡和西坡的气温也是介于南坡和北坡之间。土壤含水量(%)则在南坡最低,而在北坡最高,二者相差6.7%(牛牛山)和5.2%(卡卡山)。土壤pH在各坡面之间没有大的差异,都表现出微酸性。
| 山体及坡向 Mounts and Slopes | 融雪时间 Snow melting time/d | 气温 Air temperature/℃ | 土壤含水量 Soil water content/% | pH |
| 牛牛山Mount Niuniu: | ||||
| 东坡 East-facing slope | 102.8±8.5b | 10.3±1.6b | 46.8±6.1ab | 6.4±0.8a |
| 南坡South-facing slope | 93.8±8.6c | 12.5±2.2a | 41.7±5.1b | 6.6±0.7a |
| 西坡West-facing slope | 99.2±11.3b | 11.0±2.3ab | 43.9±5.9ab | 6.1±0.5a |
| 北坡North-facing slope | 107.6±9.7a | 9.1±1.9b | 48.4±6.2a | 6.2±0.5a |
| 卡卡山 Mount Kaka: | ||||
| 东坡 East-facing slope | 100.3±8.5b | 8.9±2.0ab | 45.4±5.7ab | 6.0±0.6a |
| 南坡South-facing slope | 94.2±8.9c | 10.8±1.9a | 42.5±6.6b | 6.2±0.5a |
| 西坡West-facing slope | 98.9±11.3b | 9.3±1.8ab | 44.8±6.8ab | 6.3±0.6a |
| 北坡North-facing slope | 108.8±9.2a | 8.2±1.6b | 47.7±7.1a | 5.9±0.4a |
| 不同的字母代表差异显著,P<0.05 | ||||
如表 2所示,暗紫贝母的前3个物候期(开始生长期、始花期和盛花期)都是在北坡最迟,在南坡最早,东坡和西坡的物候期介于南、北坡之间。以开始生长期来说,南、北坡之间相差达到13.8 d(牛牛山)和14.6 d(卡卡山);始花期在南北坡之间相差11.2 d(牛牛山)和10.9 d(卡卡山);盛花期,南北坡之间相差达到6.7 d(牛牛山)和6.3 d(卡卡山)。植株枯黄期的情况有所不同,虽然在各坡向之间有一定的差异,但差异都未达到显著的程度,即暗紫贝母在各坡面的枯萎死亡时间具有较大的一致性。另外,两山尽管所处的地理位置相差较远(大约45 km),但在相同坡面相同海拔处,暗紫贝母的同一物候期相差不大。
| 山体及坡向 Mounts and Slopes | 开始生长期/d Phase of plant emergence | 始花期/d Phase of first flowering | 盛花期/d Phase of peak flowering | 植株枯黄期/d Phase of consenescence |
| 牛牛山Mount Niuniu: | ||||
| 东坡 East-facing slope | 117.4±11.5b | 155.6±11.8b | 178.2±9.6ab | 243.0±13.7a |
| 南坡South-facing slope | 108.5±9.2c | 149.3±12.1c | 174.6±13.6b | 241.7±14.8a |
| 西坡West-facing slope | 115.8±7.8b | 154.8±13.0b | 177.6±11.9ab | 242.6±11.8a |
| 北坡North-facing slope | 122.3±10.5a | 160.5±12.5a | 181.3±14.2a | 244.5±15.1a |
| 卡卡山 Mount Kaka: | ||||
| 东坡 East-facing slope | 115.1±10.8b | 156.7±13.4b | 177.2±12.7ab | 244.3±11.8a |
| 南坡South-facing slope | 106.2±8.9c | 151.8±11.2c | 173.8±14.1b | 242.6±15.3a |
| 西坡West-facing slope | 114.7±11.3b | 158.2±9.6b | 176.8±12.5ab | 243.8±14.8a |
| 北坡North-facing slope | 120.8±9.5a | 162.7±10.7a | 180.1±16.3a | 246.1±16.2a |
从单叶面积来看,不管是牛牛山还是卡卡山都是北坡最大,南坡最小,而东坡与西坡的值则介于南坡与北坡之间。和南坡相比,北坡叶面积值要大29.5%(牛牛山)和31.9%(卡卡山)。南、北坡面之间的差异显著,而其他坡面之间的差异并不显著(表 3)。比叶面积和叶面积表现出了同样的变化规律,也是北坡最大,南坡最小。在牛牛山北坡比叶面积达到(12.3±1.0) m2/kg,比南坡大24.2%;而卡卡山北坡达到(13.7±1.1)m2/kg,比南坡大29.2%。无论哪一山,南、北坡面间比叶面积差异都达到显著,而其他坡面之间的差异并不显著(表 4)。
| 坡向 Slopes | 牛牛山/(10-4m2) Mount Niuniu | 卡卡山/(10-4m2) Mount Kaka | 坡向 Slopes | 牛牛山/(10-4m2) Mount Niuniu | 卡卡山/(10-4m2) Mount Kaka |
| 东坡East-facing slope | 1.82±0.37ab | 1.89±0.21ab | 南坡South-facing slope | 1.59±0.34b | 1.66±0.27b |
| 西坡West-facing slope | 1.91±0.22ab | 1.95±0.38ab | 北坡North-facing slope | 2.06±0.28a | 2.19±0.35a |
| 坡向 Slopes | 牛牛山/(m2/kg) Mount Niuniu | 卡卡山/(m2/kg) Mount Kaka | 坡向 Slopes | 牛牛山/(m2/kg) Mount Niuniu | 卡卡山/(m2/kg) Mount Kaka |
| 东坡East-facing slope | 11.2±0.9ab | 12.3±1.2ab | 南坡South-facing slope | 9.9±0.8b | 10.6±0.9b |
| 西坡West-facing slope | 10.9±0.7ab | 11.8±1.5ab | 北坡North-facing slope | 12.3±1.0a | 13.7±1.1a |
如表 5所示,暗紫贝母的植株株高以北坡最高而南坡最小。以卡卡山为例,北坡的株高比南坡大21.4%;而在牛牛山,北坡的株高比南坡大30.3%。东、西坡的株高介于南北坡之间。就鳞茎生物量而言,其所变化的趋势与株高相反,即南坡最大,北坡最小,南坡比北坡大11.7%(牛牛山)和10.2%(卡卡山),东、西坡面的值也是介于南坡和北坡之间。
| 山体及坡向 Mounts and slopes | 牛牛山Mount Niuniu | 卡卡山Mount Kaka | ||||||
| 东坡 East- facing slope | 南坡 South- facing slope | 西坡 West- facing slope | 北坡 North- facing slope | 东坡 East- facing slope | 南坡 South- facing slope | 西坡 West- facing slope | 北坡 North- facing slope | |
| 植株株高 Height of stems/cm | 18.2±2.1ab | 15.8±1.6b | 17.9±2.0ab | 20.6±1.7a | 18.0±1.5ab | 16.3±1.9b | 17.7±2.1ab | 19.8±2.8a |
| 鳞茎生物量 Biomass of bulbs/mg | 230.9±20.5ab | 238.4±15.9a | 224.6±14.6b | 213.5±10.2c | 229.7±17.1ab | 241.2±18.7a | 231.5±13.6ab | 218.8±12.5b |
无论牛牛山还是卡卡山,单粒果实的生物量在各坡面之间无明显的差异(图 1),这一点与株高和鳞茎生物量呈现出的趋势不同。从果实生物量在地上部分中所占的比例(%)来看,在南坡最大,达到45.7%(卡卡山)和48.0%(牛牛山),而北坡则最小,只有39.3%(卡卡山)和40.32%(牛牛山)。南北坡面之间的差异显著,其他坡面之间差异不显著(图 2)。

|
| 图 1 各坡面单个果实生物量比较 Fig. 1 Comparison of single fruit biomass amnog four slopes of Mount Kaka and Mount Niuniu |

|
| 图 2 各坡面单个果实生物量与地上部分生物量之比 Fig. 2 Ration of single fruit biomass to aboveground biomass |
单粒果实平均种子数以北坡最多,达到85.8粒(卡卡山)和80.4粒(牛牛山);相比较而言,南坡最少,仅有67.6粒(卡卡山)和70.6粒(牛牛山),东坡和西坡的数值介于南、北坡向之间。与果实平均种子数不同,种子千粒重则以南坡最重,北坡最轻。以卡卡山为例,南坡千粒重比北坡大319.1 mg,大32.3%;在牛牛山,南坡千粒重比北坡大271.1 mg,大28.1%(图 3,图 4)。在南、北坡向之间差异显著,而其他坡向之间差异不显著。

|
| 图 3 各坡面上的单果种子数 Fig. 3 Number of seeds per fruit at all slopes |

|
| 图 4 各坡面种子千粒重比较 Fig. 4 Comparison of 1000 seeds weight amont four slopes |
植物物候期的早迟,不仅可以影响同一物种不同种群间传粉方面的差异[26],而且还可以影响到果实和种子的大小[27],对植物完成生活史有着十分重要的意义。在卡卡山和牛牛山各坡面中,南坡融雪时间最早,且早春(3—5月)气温相对较高(表 1,表 2),植株萌动(开始生长)较早(表 3);北坡融雪时间最迟,早春气温也最低,植株萌动(开始生长)也最晚。这表明,暗紫贝母物候期在各坡面的差异,要受各坡面融雪时间和气温的影响。有研究发现,在积雪融化相对较早的地方植物开始生长也较早,而积雪融化迟的区域植物开始生长也较迟[28];Kudo研究了5种雪生植物的物候,发现融雪时间的早迟与这些植物开花和结果的先后有很大的一致性[29];Galen等的研究也认为,积雪消融的时间对生长于雪下的植物有相当大的影响,其开花时间在早融雪部位可以比晚融雪部位提前20多天[24];还有学者比较了一些雪生植物的开花物候与积雪消融之间的关系,发现同一种雪生植物在积雪晚融的年份比积雪早融的年份开花时间要明显地迟一些[30]。显然,这些研究结果和本研究的结果是很相似的。只是,本研究地点由于处在低纬度高海拔区域(青藏高原东部),各坡向积雪消融和物候期的时间差异没有上述研究中的那么大。另外,Suzuki等利用人工OTC模拟增温对物候的影响进行研究发现,增温确实可以使展叶和开花等物候期提前[31],本研究中的南向坡面因气温较高,物候期较早出现,本文的结果与之也是相符合的。可见,积雪的早融和较高的气温会导致植物物候期的提前,这具有相当的普遍性。
3.2 植株及鳞茎在坡面间的差异在青藏高原东部的高山生境,不管哪个坡面都存在着一年一度的雪积累、雪融化、植物生长以及休眠(一年生植物为死亡)这样一种周期性循环。这种季节性积雪融化时间的早晚可能通过生态因子的级联变化对植物的生长产生影响。在北坡,由于融雪晚,土壤湿度相对大一些(表 1,表 2);并且,延迟的融雪会使植物的生长处于生长季中更温暖的时期。因此,北坡暗紫贝母株高更高一些。而在南坡由于土壤相对干燥,同时积雪的早融导致植株过早萌动,这反而会由于早春的夜间低温[21-22]而造成冻害,影响其生长,故而植株相对低矮一些,也就是说,植株对提前的融雪反而表现出负的生长效应,这与其他学者的研究结果[32-34]是一致的。
在北坡,植株单叶面积和比叶面积都比南坡要大一些,其原因主要是北坡是阴坡,光照强度相对较弱,且融雪时间较晚,土壤水分含量较高。在这样的环境中,单叶面积和比叶面积的增大有利于其提高光合效率和积累有机养料,这对于北坡的植株顺利完成生活史是有益的。而南坡属于阳坡,土壤水分含量相对较低,气温相对较高,光照相对较强,植株叶片的蒸藤强度较大,其单叶面积和比叶面积的缩小有利于其减少蒸藤,这对于其生存也是有利的。李明财等在东北阔叶红松林中的研究表明,随着光照强度降低,所有树种比叶面积呈现增加的趋势[35];而黄菊莹等在温带草原中的研究也显示,适量增加水分供给可显著提高羊草的比叶面积[36]。可见本研究的结果与这些结论是一致的。
暗紫贝母鳞茎在南坡生物量较大,而在北坡生物量较小,这可能跟南坡积雪融化较早、生长期相对较长有一定的关系。因为生长期越长,营养物质积累越多,再加上南坡植株地上部分相对低矮(表 5),地下的鳞茎便可获得相对更多的营养物质,于是鳞茎生物量相对就大了。在高山草甸群落中的研究也显示了类似的结果[37-39]。当然,南坡鳞茎生物量较大可能还与南坡土壤相对干燥有关。因为在干旱的环境中,植物为了应对水分的胁迫,势必减少地上部分的生长从而有效降低水分的散失[40],这样一来自然也就增加了地下部分的产量。孙儒泳等也认为,在相对潮湿的土壤中,植物生长不发达的浅根系,而在相对干燥的土壤中,植物具有发达的深根系[41]。
3.3 果实和种子特征在坡面间的差异在积雪早融的区域,植物较早地失去了雪被的保护,会增加霜冻损害的风险,导致其繁殖活性降低[34, 42];高山植物的繁殖器官对零下低温十分敏感,从雪被中过早暴露会增加霜冻的风险[43]。这些可能是暗紫贝母单个果实的种子数在早融雪的南坡明显减少(图 3)的重要原因。另外,早春时期积雪早融导致的零下低温会使访花昆虫的多样性和活动频率有所降低[44],从而影响传粉的有效性,引起胚珠的受精率下降,于是南坡种子数减少,孙守如等在研究授粉次数和授粉时间对西葫芦种子数量的影响时也得出了类似的结论[45-46]。不过,南坡的暗紫贝母还是将有限的营养资源用在了种子的生长上,虽然种子的数量减少了,但种子的大小并未受影响,反而还有所增大,这从种子的千粒重上可以反映出来(图 4)。高山地区不同部位融雪时间的差异可以影响雪生植物种子的大小[47],本研究的结果与之一致。种子的大小又会进一步影响幼苗的生长发育及形态大小[48],较大的种子可以提升幼苗的适合度或存活力[49],这对于暗紫贝母幼苗在条件相对更为恶劣的生境中顺利成长并完成其生活史显得十分必要。
3.4 全球变暖条件下高山植物可能的变化全球变暖是一个不争的事实,随着气温的升高,高山地区的积雪会相对早融[50]。气温升高以及积雪早融引起暗紫贝母的生长和繁殖特性发生相应的变化,如种子数量减少、种子和鳞茎变大,这些对于人工培育暗紫贝母(特别是其最佳生境的选择),有着较大的参考意义。同时,暗紫贝母是高山雪生植物,存在着一定的分布海拔范围[4, 6],全球气温升高后,其分布区域可能也会发生相应的改变。在青藏高原东部的高山草甸群落中,类似于暗紫贝母这样的草本植物还有很多,全球变暖使这些植物的个体生长特性、繁殖、分布都可能发生一定的变化,群落的结构和整个植被的格局也可能会随之改变。
| [1] | Bennie J, Huntley B, Wiltshire A, Hill M O, Baxter R. Slope, aspect and climate: Spatially explicit and implicit models of topographic microclimate in chalk grassland. Ecological Modelling , 2008, 216 (1) : 47–59. DOI:10.1016/j.ecolmodel.2008.04.010 |
| [2] | Carletti P, Vendramin E, Pizzeghello D, Concheri G, Zanella A, Nardi S, Squartini A. Soil humic compounds and microbial communities in six spruce forests as function of parent material, slope aspect and stand age. Plant and Soil , 2009, 315 (1/2) : 47–65. |
| [3] | Gong X Y, Brueck H, Giese K M, Zhang L M, Sattelmacher B, Lin S. Slope aspect has effects on productivity and species composition of hilly grassland in the Xilin River Basin, Inner Mongolia, China. Journal of Arid Environments , 2008, 72 (4) : 483–493. DOI:10.1016/j.jaridenv.2007.07.001 |
| [4] | 四川植物志编辑委员会. 四川植物志(第七卷). 成都: 四川民族出版社 , 1991 : 57–59. |
| [5] | 徐国钧. 中草药彩色图谱(修订本). 福州: 福建科学技术出版社 , 1997 : 210–213. |
| [6] | 青海植物志编辑委员会. 青海植物志(第四卷). 西宁: 青海人民出版社 , 1999 : 277–278. |
| [7] | 于婧, 魏建和, 陈士林, 代勇, 杨成民. 川贝母种子休眠及萌发特性的研究. 中草药 , 2008, 39 (7) : 1081–1084. |
| [8] | 刘燕华, 李秀彬. 脆弱生态环境与可持续发展. 北京: 商务印书馆 , 2001 : 22–28. |
| [9] | 桂镜生, 杨树德. 川贝母与平贝母的资源状况调查及市场供求分析. 云南中医学院学报 , 2008, 31 (6) : 36–39. |
| [10] | 刘辉, 陈士林, 姚辉, 李西文, 张艺. 川贝母的资源学研究进展. 中国中药杂志 , 2008, 33 (14) : 1645–1648. |
| [11] | 朱丹妮, 蒋莹, 陈婷, 高山林. 组织培养川贝母化学成分和药理作用的研究. 中国药科大学学报 , 1992, 23 (2) : 118–121. |
| [12] | 韵海霞, 陈志. 暗紫贝母乙醚萃取物的GC-MS分析. 青海医学院学报 , 2009, 30 (2) : 133–135. |
| [13] | 李隆云, 周裕书, 代敏, 罗庆明. 暗紫贝母鳞茎再生组织培养技术研究. 中国中药杂志 , 1995, 20 (2) : 78–80. |
| [14] | 蔡朝晖, 高山林, 徐德然, 朱丹妮. 不同培养条件及方法对组培暗紫贝母生长的影响. 中国药科大学学报 , 1992, 23 (6) : 367–369. |
| [15] | 高山林, 夏艳, 谭丰苹. 组织培养暗紫贝母的药理作用. 植物资源与环境学报 , 2000, 9 (1) : 4–8. |
| [16] | 张辉东, 曾雪丽, 陈鹊, 吴卫. 暗紫贝母内生真菌红色素的提取及稳定性研究. 湖北农业科学 , 2011, 50 (18) : 3815–3818. |
| [17] | 陈士林, 肖小河, 陈善墉. 暗紫贝母植被分布格局的数值分析. 西南师范大学学报: 自然科学版 , 1997, 22 (4) : 416–420. |
| [18] | 陈士林, 肖小河, 陈善墉. 暗紫贝母的群落生态研究. 中药材 , 1989, 12 (11) : 5–8. |
| [19] | 徐波, 王金牛, 石福孙, 高景, 吴宁. 青藏高原东缘野生暗紫贝母生物量分配格局对高山生态环境的适应. 植物生态学报 , 2013, 37 (3) : 187–196. |
| [20] | 陈文年, 陈发军, 谢玉华, 王淯, 段辉国, 齐泽民. 暗紫贝母的物候和鳞茎在海拔梯度上的变化. 草业学报 , 2012, 21 (5) : 319–324. |
| [21] | Chen W N, Wu Y, Wu N, Luo P. Effect of snow-cover duration on plant species diversity of alpine meadows on the eastern Qinghai-Tibetan Plateau. Journal of Mountain Science , 2008, 5 (4) : 327–339. DOI:10.1007/s11629-008-0182-0 |
| [22] | Chen W N, Wu Y, Wu N, Wang Q. Effect of snowmelt time on growth and reproduction of Pedicularis davidii var. pentodon in the eastern Tibetan Plateau. Plant Biosystems , 2011, 145 (4) : 802–808. DOI:10.1080/11263504.2011.599989 |
| [23] | 马德隆. 松潘县志. 北京: 民族出版社, 1999101–148. |
| [24] | Galen C, Stanton M L. Consequences of emergence phenology for reproductive success in Ranunculus adoneus (Ranunculaceae). American Journal of Botany , 1991, 78 (7) : 978–988. DOI:10.2307/2445177 |
| [25] | 李玉霖, 崔建垣, 苏永中. 不同沙丘生境主要植物比叶面积和叶干物质含量的比较. 生态学报 , 2005, 25 (2) : 304–311. |
| [26] | Waser N M. Competition for hummingbird pollination and sequential flowering in two Colorado wildflowers. Ecology , 1978, 59 (5) : 934–944. DOI:10.2307/1938545 |
| [27] | Hendrix S D. Variation in seed weight and its effects on germination in Pastinaca sativa L. (Umbelliferae). American Journal of Botany , 1984, 71 (6) : 795–802. DOI:10.2307/2443470 |
| [28] | Holway J G, Ward R T. Phenology of alpine plants in northern Colorado. Ecology , 1965, 46 (1/2) : 73–83. |
| [29] | Kudo G. Performance and phenology of alpine herbs along a snow-melting gradient. Ecological Research , 1992, 7 (3) : 297–304. DOI:10.1007/BF02347098 |
| [30] | Kudo G, Hirao A S. Habitat-specific responses in the flowering phenology and seed set of alpine plants to climate variation: implications for global-change impacts. Population Ecology , 2006, 48 (1) : 49–58. DOI:10.1007/s10144-005-0242-z |
| [31] | Suzuki S, Kudo G. Short-term effects of simulated environmental change on phenology, leaf traits, and shoot growth of alpine plants on a temperate mountain, northern Japan. Global Change Biology , 1997, 3 (S1) : 108–115. DOI:10.1111/j.1365-2486.1997.gcb146.x |
| [32] | Galen C, Stanton M L. Short-term responses of alpine buttercups to experimental manipulations of growing season length. Ecology , 1993, 74 (4) : 1052–1058. DOI:10.2307/1940475 |
| [33] | Galen C, Stanton M L. Responses of snowbed plant species to changes in growing-season length. Ecology , 1995, 76 (5) : 1546–1557. DOI:10.2307/1938156 |
| [34] | Saavedra F, Inouye D W, Price M V, Harte J. Changes in flowering and abundance of Delphinium nuttallianum (Ranunculaceae) in response to a subalpine climate warming experiment. Global Change Biology , 2003, 9 (6) : 885–894. DOI:10.1046/j.1365-2486.2003.00635.x |
| [35] | 李明财, 朱教君, 孙一荣. 东北次生林主要树种比叶面积对光照强度的响应. 生态学杂志 , 2009, 28 (8) : 1437–1442. |
| [36] | 黄菊莹, 袁志友, 李凌浩. 羊草绿叶氮、磷浓度和比叶面积沿氮、磷和水分梯度的变化. 植物生态学报 , 2009, 33 (3) : 442–448. |
| [37] | 李英年. 高寒草甸植物地下生物量与气象条件的关系及周转值分析. 中国农业气象 , 1998, 19 (1) : 36–38. |
| [38] | 刘伟, 周华坤, 周立. 不同程度退化草地生物量的分布模式. 中国草地 , 2005, 27 (2) : 9–15. |
| [39] | 陈文年, 吴彦, 吴宁, 罗鹏. 高山草甸群落生物量在融雪梯度上的变化. 应用与环境生物学报 , 2009, 15 (6) : 745–749. |
| [40] | 王艳芬, 汪诗平. 不同放牧率对内蒙古典型草原地下生物量的影响. 草地学报 , 1999, 7 (3) : 198–203. |
| [41] | 孙儒泳, 李庆芬, 牛翠娟, 娄安茹. 基础生态学. 北京: 高等教育出版社, 200237–39. |
| [42] | Inouye D W, Morales M A, Dodge G J. Variation in timing and abundance of flowering by Delphinium barbeyi Huth (Ranunculaceae): the roles of snowpack, frost, and La Niña, in the context of climate change. Oecologia , 2002, 130 (4) : 543–550. DOI:10.1007/s00442-001-0835-y |
| [43] | Billings W D. Adaptations and origins of alpine plants. Arctic and Alpine Research , 1974, 6 (2) : 129–142. DOI:10.2307/1550081 |
| [44] | Bingham R A, Orthner A R. Efficient pollination of alpine plants. Nature , 1998, 391 (6664) : 238–239. DOI:10.1038/34564 |
| [45] | 孙守如, 陈艳丽, 杨子琴, 高新生, 陈红兵. 不同授粉次数对西葫芦种子数量和质量的影响. 种子 , 2007, 26 (10) : 27–28. |
| [46] | 孙守如, 陈艳丽, 宋留高, 李静. 不同授粉时间对西葫芦种子数量与质量的影响. 河南农业大学学报 , 2004, 38 (3) : 289–291. |
| [47] | Jerling L. Genetic differentiation in fitness related characters in Plantago maritima along a distributional gradient. Oikos , 1988, 53 (3) : 341–350. DOI:10.2307/3565534 |
| [48] | Lacey E P, Pace R. Effect of parental flowering and dispersal times on offspring fate in Daucus carota (Apiaceae). Oecologia , 1983, 60 (2) : 274–278. DOI:10.1007/BF00379533 |
| [49] | 陈文年, 陈发军, 谢玉华. 融雪时间对川贝母生长的影响. 湖北农业科学 , 2013, 52 (4) : 868–872. |
| [50] | Whetton P H, Haylock M R, Galoway R. Climate change and snow-cover duration in the Australian Alps. Climatic Change , 1996, 32 (4) : 447–479. DOI:10.1007/BF00140356 |