 2016, Vol. 36
2016, Vol. 36文章信息
- 李雯雯, 李丽, 吴巩胜, 张宇, 代云川, 周跃
- LI Wenwen, LI Li, WU Gongsheng, ZHANG Yu, DAI Yunchuan, ZHOU Yue.
- 评估不同尺度下农牧地对滇金丝猴景观连接度的影响
- Impact evaluation on the Yunnan snub-nosed monkey's habitat in terms of landscape connectivity by agricultural land with consideration at different scale scenarios
- 生态学报[J]. 2016, 36(24): 8136-8144
- Acta Ecologica Sinica[J]. 2016, 36(24): 8136-8144
- http://dx.doi.org/10.5846/stxb201505211030
-
文章历史
- 收稿日期: 2015-05-21
- 网络出版日期: 2016-04-13





2. 云南财经大学野生动植物管理与生态系统健康研究中心, 昆明 650221;
3. 昆明理工大学环境科学与工程学院, 昆明 650093;
4. 云南师范大学旅游与地理科学学院, 昆明 650500
2. Wildlife Management and Ecosystem Health center, Yunnan University of Finance and Economics, Kunming 650221, China;
3. Faculty of Environmental Science and Engineering, Kunming University of Science and Technology, Kunming 650093, China;
4. School of Tourism and Geographical Science, Yunnan Normal University, Kunming 650500, China
人类活动导致野生动物生境的丧失和破碎化已成为当前生物多样性保护面临的最大威胁[1-3]。尤其是近年来土地利用中农牧地的开垦造成大量的热带原始森林被砍伐和破坏,使得灵长类动物由于生境破碎化,被限制在互不相连的保护区内,即使在同一个保护区内,多数物种之间亦缺乏生境的延续性而呈孤岛状分布[4]。生境破碎化是指原来成片的生境,被分割、破碎,形成分散、孤立的岛状生境或生境破碎的现象。破碎化的增加致使生境斑块间连接度丧失,导致种群间基因交流困难、连接网络中断,种群只能存活于面积小且隔离的生境斑块中,长期以往物种将面临基因多样性丧失和高灭绝的风险[5-6]。许多学者在基于景观连接度评估生境破碎化对区域生物多样性带来的影响中发现,只要保持较高的景观连接度该种群仍可能在区域中续存[7]。景观连接度研究也由此成为区域生物多样性保护的主要途径之一。
景观连接度描述了景观要素在功能和生态学过程上的有机联系,这种联系可能是生物群体间的物种流,也可能是景观要素间直接的物质、能量与信息流[8],它为探索景观空间异质性和揭示景观空间格局与生态过程间的关系提供了理论和技术方法[9]。在自然界中的灵长类动物是没有固定家域的哺乳类动物(free-ranging mammals),其景观完整性与连通性直接影响灵长类动物的分布与生存[10],而影响景观完整性的主要障碍在于人类土地利用,尤其是农牧地的变化直接影响了景观要素在功能和生态学过程中的联系[11]。因此,基于斑块(patch)-景观(landscape)尺度上农牧地对景观连接度的影响研究有助于灵长类动物廊道的恢复[11-13],有助于灵长类动物在破碎生境中去识别适宜生境而维持生存[14-15]。然而,在过去20年前,灵长类学家才开始去关注灵长类动物对景观变化的反应[16-19],但是关于土地利用,尤其是农牧地对景观连接度的影响却很少被关注。
在景观连接度的研究中,图论法Graph theory(也称为网络分析network analysis)借助GIS软件平台,利用直观的空间生境数据并结合物种的扩散行为,能很好的预测景观潜在功能的连接度,被认为是高性价比的方法[20],该方法中阈值的确定是图论法应用的关键。
本文选取滇金丝猴这一珍稀濒危物种,综合最小费用距离和猴群活动的最小家域,建立潜在廊道及廊道缓冲区,在不同尺度上选择缓冲区内的农牧地斑块,通过不同的情景假设,运用图论法原理,定量分析景观尺度与斑块尺度上农牧地对滇金丝猴栖息地景观连接度的影响。旨在探讨以下两个问题:(1)在景观尺度(landscape)上,整体分析农牧地对滇金丝猴栖息地连接度的影响,定量评价滇金丝猴栖息地景观连接度的变化;(2)在斑块尺度(patch)上,基于未来不同农牧地的变化情景,分析15个滇金丝猴栖息地斑块尺度上的组分中景观连接度的变化以及组分数量发生变化对于景观连接度的影响。
1 研究区域与种群滇金丝猴是我国特有的珍稀濒危物种之一,长年生活在人迹罕至的高山地带,是海拔分布最高的非人灵长类。滇金丝猴的活动范围从海拔1800m至4500m[21],活动量大,活动范围较广。滇金丝猴自然种群仅存15个(12个在云南境内,3个在西藏地区),约2500只(表 1)[22]。该物种栖息地位于三江并流区域,分布范围东西界为金沙江和澜沧江,分布最北的猴群纬度为29°20′N,最南的一个猴群分布纬度为26°14′N,即分布于云南的德钦、维西、兰坪、丽江和西藏的芒康5县境内,其栖息地相互处于分离状态,成岛屿状分布[23-25]。
| 猴群 Group | 猴群栖息地 Monkey Habitat | 种群数量/只 Number | 活动范围/km2 Group range | 管理单元[26] Management unit |
| G1 | 执都 | 50 | 24.2 | S1 |
| G2 | 小昌都 | >200 | 47.3 | S1 |
| G3 | 米粒卡 | 100 | 10.7 | S1 |
| G4 | 巴美 | 80 | 117.6 | S1 |
| G5 | 吾牙普牙 | >300 | 180.4 | S2 |
| G6 | 茨卡通 | 50 | 61.9 | S3 |
| G7 | 弄资河头、巴迪河头、归龙 | 100 | 208.4 | S3 |
| G8 | 同洛河头、施坝 | 200 | 131.6 | S3 |
| G9 | 格花箐、响古箐 | >900 | 82.9 | S3 |
| G10 | 大草坝各玛、巴倮河头、仙人洞 | 30 | 36.4 | S3 |
| G11 | 金丝厂 | 250 | 41.5 | S4 |
| G12 | 大坪子 | <50 | 41 | S4 |
| G13 | 长岩山 | 120 | 22.7 | S5 |
| G14 | 拉沙山 | 100 | 16.5 | S5 |
| G15 | 龙马山 | >100 | 14.8 | S5 |
| G1—G3为西藏种群,G4—G15为云南种群 | ||||
研究区植被图由云南省林业调查规划设计院根据2012年SPORT5卫星遥感影像解译获得,以1∶50000比例尺水系图和数字高程模型(DEM)作为参考的控制影像,在ERDAS9.2中对整景影像进行了几何精校正,均方根误差(RMS)<1,植被图满足研究精度要求。滇金丝猴活动范围来自龙勇诚2004—2009年调查数据(未发表数据),遗传距离来源于刘志瑾2009年发表的研究论文[26],本研究仅采用云南地区的遗传距离。
2.2 研究方法 2.2.1 绘制滇金丝猴栖息地潜在景观廊道由于研究区域内植被种类较多,为了分析的准确性,本研究结合《云南植被》分类体系和滇金丝猴生境相关文献[22, 27-28]将研究区植被重新划分为5类(表 2),本文潜在景观廊道的绘制,结合景观遗传学,根据各生境类型的费用赋值[29]得到最小费用路径即滇金丝猴的潜在扩散廊道。
连接度与廊道有着密切关系。当连接度较好时,物种在廊道中迁徙、扩散受到的阻力较小,其觅食、繁殖和生存也会比较容易;当连接度较差时,物种在廊道中迁徙、扩散和觅食将受到更多的限制,运动的阻力较大,生存困难。道路、沟渠、堤坝等人工设施,通常是影响生物廊道连接度的重要因素,而生物廊道上退化或受到破坏的片段也是导致连接度降低的因素[30]。
研究区域中农牧地斑块数量非常多,滇金丝猴的活动范围与研究区域的总面积相差甚小,因此本文仅将廊道缓冲区内的农牧地作为评估对滇金丝猴栖息地连接度的影响,参考专家知识以潜在扩散廊道为中心,以猴群的最小家域14.7km2[21]为阈值去计算适宜的廊道缓冲区。
2.2.3 确定最佳距离阈值及评估不同管理单元间猴群的可能扩散在同一个管理单元内,滇金丝猴交流的可能性极大,其连接度也大,然而相邻管理单元之间滇金丝猴交流可能性很小,其连接度也非常小,管理单元之间的连接度概率也较低。根据已有的研究,将云南境内的12个滇金丝猴栖息地分成5个管理单元[26],通过对不同函数拟合分析滇金丝猴栖息地遗传距离与最小费用距离的相关性,分别求出相邻管理单元之间的距离,将其管理单元之间最小的费用距离作为评估两两相邻管理单元之间的猴群可能进行扩散的最佳距离阈值。
2.2.4 改善农牧地斑块对滇金丝猴栖息地连接度的影响影响景观连接度的因素有多个方面,不仅和景观的空间结构有着密切的关系,而且与生态过程对象有关。其中廊道的组成物质、宽度、形状、长度都将影响景观连接度水平[8]。本文通过假设4种潜在农牧地斑块的改善情景,C1:用不适宜生境(费用赋值为90)优化为廊道缓冲区内的农牧地(生境类型为阻碍,费用赋值为100);C2:用适宜生境(费用赋值为70)优化为廊道缓冲区内的农牧地;C3:用次适宜生境(费用赋值为10)优化为廊道缓冲区内的农牧地;C4:用最适宜生境(费用赋值为1)优化为廊道缓冲区内的农牧地。在不同的情景中,根据猴群可能扩散的最佳距离阈值,运用图论法中的可能连通性指数(PC)的计算来评估景观尺度与斑块尺度上农牧地对滇金丝猴栖息地连接度的影响。
在景观尺度上,PC是用来度量整体连接度的变化,指数的计算需要指定景观中生境斑块连通的距离阈值。当斑块之间的距离大于阈值,认为这两个斑块不连通;小于或等于阈值,则认为它们是连通的。
PC指数在Global水平上的表达式为:
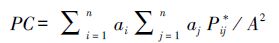
|
(1) |
式中,ai和aj分别为斑块i和j的面积,A是整个景观的面积(包括研究区域的面积,比如栖息地和非栖息地)。这些变化可能也涉及到不同于斑块面积的属性,例如栖息地的质量或者一些其它斑块的特性。在分析中,这些特性可能被考虑是相关的(衡量面积的质量,承载能力,聚集死亡率,人口大小,栖息地适宜性,核心区域等)。Pij*代表在斑块i和斑块j之间潜在的路径中,连接度最大的概率。如果斑块i和j距离很近,最大可能的路径将类似于斑块i和j之间直接运动(Pij*=Pij),如果斑块i和j距离很远,则Pij*>Pij[31]。
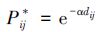
|
(2) |
式中,dij指最小费用距离阈值,α是随着p和d变化的。式中可见,随着不同的最小费用距离阈值d,其斑块间连接的可能性大小也发生变化。
在斑块尺度上,PC指数计算与斑块水平的公式相同,此时计算得到的PC指数是代表不同的组分的连接度,距离阈值不仅是作为两个斑块是否连接的条件也是作为划分组分的判定,通过确定出的最佳距离阈值,将整个研究区域分成不同的组分,小于等于阈值两两斑块在同一个组分中,大于阈值则不在一个组分。
3 结果与分析 3.1 廊道及廊道缓冲区的建立图 1表明了研究区域和管理单元以及依据已有的最小费用的赋值,所获得猴群间潜在的扩散廊道,以潜在扩散廊道为中心,以确保不超过猴群的最小14.7km2家域为阈值,计算获得缓冲区的距离为2.15km,在2.15km廊道缓冲区范围内,确定用以研究的农牧地斑块为768块。

|
| 图 1 研究区域及廊道和廊道缓冲区 Fig. 1 Study area、corridor and corridor buffer |
表 3表明了对遗传距离和最小费用距离进行的4种函数类型的拟合,幂函数(R2=0.408)为拟合遗传距离与最小费用距离的相关性的最优函数。
| 函数类型Function type | 表达式Expression | R2 | 样本数量NSample size |
| 线性函数Linear function | y=2×10-6x+0.069 | 0.359 | 32 |
| 对数函数Log function | y=0.024lnx-0.103 | 0.396 | 32 |
| 指数函数Exponential function | y=0.059e2×10-5x | 0.361 | 32 |
| *幂函数Power function | y=0.009x0.258 | 0.408 | 32 |
| *为最优函数 | |||
图 2表明,遗传距离与最小费用距离成正相关,即最小费用距离越大,其遗传距离也越大,意味着遗传的差异性也越大。在图 1所示的5个管理单元内,如果相邻管理单元之间的遗传距离较大,认为不会连接,由此分别计算出相邻管理单元距离,取其最小值为最佳阈值,超过这个阈值,滇金丝猴交流的可能性极小。通过计算,得到两两相邻管理单元之间的最小费用距离为1600,因此取1600为最佳距离阈值。除此之外,通过观察拟合函数(图 2),估测出当最小费用路径距离为0时,遗传距离为0.059,说明在同一个滇金丝猴活动范围内猴群间也存在差异性;根据拟合函数可以看出,一开始函数斜率增长得较快,即遗传距离对最小费用距离的增加较敏感,随后函数斜率增长缓慢,即遗传距离对最小费用距离的增加不敏感,由此估测出其斜率变化的转折点,与相邻管理单元最小费用距离十分相近,以此来确定不同管理单元间猴群可能扩散的最小费用距离阈值。当最佳距离阈值为1600时,在两个斑块之间的费用距离小于或等于1600时,这两个斑块是连接的,猴群可能在这两个斑块进行扩散,超过费用距离1600,两个斑块不再连接,即猴群在这两个斑块间不会扩散。

|
| 图 2 遗传距离与路径距离相关性 Fig. 2 Collection between genetic distance and path distance 1: 代表管理单元间最小费用距离;2: 代表管理单元之间平均最小费用距离;3: 代表管理单元之间最大费用距离 |
图 3表明了用以研究的农牧地斑块。表 4表明了通过对基于距离阈值1600所计算出的PC指数及其它指数发现,在景观尺度下,在四种不同的情景中,PC指数是逐渐增加的,增加的幅度越来越大。表 4中显示了不同指数在不同情景中的变化情况。在假设情景C1中,PC值增加了6.83%,即情景C1可以恢复6.83%的连接度,此时连接数也增加了2个,节点没有发生变化;在假设情景C2中,此时PC指数增加了7.28%,即情景C2可以恢复7.28%的连接度,此时连接数也增加5个,节点没有变化;在情景C3中,此时PC值增加了10.09%,即情景C3可以恢复10.09%的连接度,此时连接数增加了9个,节点没有变化;在情景C4中,此时PC指数增加了30.43%,连接度显著增加,连接数增加了53个,节点也增加了43个。

|
| 图 3 缓冲区内农牧地分析 Fig. 3 Agricultural land in the corridor buffer |
| 情景 | PC指数 | 连接数 | 节点数 | PC变化率/% | 连接数变化 | 节点数变化 |
| Scenario | PC index | Link number | Node number | The rate of PC | The change of link number | The change of node number |
| C1 | 9.92×10-4 | 2447 | 1815 | 6.83 | 2 | 0 |
| C2 | 9.96×10-4 | 2450 | 1815 | 7.28 | 5 | 0 |
| C3 | 1.02×10-3 | 2454 | 1815 | 10.09 | 9 | 0 |
| C4 | 1.21×10-3 | 2498 | 1854 | 30.43 | 53 | 43 |
基于距离阈值1600将整个研究区域划分了80个组分(图 4),其中猴群分布在3个组分中,分别为图 4中的a,b,c组分。在表 5中,分析了这3个组分在不同情景中连接度的变化,其中组分b的PC指数都比原先的PC指数小,减小的比率越来越小,在情景C2,C3,C4情景中,组分a,c的PC指数增加,且增加的比率越来越大,说明了在不同的情景假设中,景观连接度在逐渐的发生变化,在组分a和c中,景观连接度有所恢复。

|
| 图 4 原始组分及组分变化 Fig. 4 The beginning component and the changing component |
| 情景Scenario | C1 | C2 | C3 | C4 |
| 组分a(PC) Component a | 1.83×10-9 | 9.90×10-4 | 1.02×10-3 | 1.20×10-3 |
| 组分b(PC) Component b | 5.28×10-8 | 1.56×10-6 | 1.84×10-6 | 7.26×10-6 |
| 组分c(PC) Component c | 3.92×10-7 | 5.28×10-8 | 5.28×10-8 | 5.60×10-8 |
| 组分a PC变化率 Rate of change of PC of component a/% | -100.00 | 7.33 | 10.12 | 30.26 |
| 组分b PC变化率 Rate of change of PC of component b/% | -99.96 | -98.93 | -98.74 | -95.03 |
| 组分c PC变化率 Rate of change of PC of component c/% | 645.25 | 0.47 | 0.38 | 6.46 |
在情景C1,C2中,组分数量没有发生变化;在情景C3和C4中,组分数量发生了变化,其中情景C3组分为78个,减少了2个组分,情景C4中组分为76 个,减少了4个组分,说明了减少的组分距离阈值小于等于1600与别的相邻的组分融为一个组分,在图 4中,在有猴群的组分中,只有组分b发生了变化,在情景C3和C4中,原来的组分b与右边的组分融为一个组分,虽然组分b的景观连接度降低,但是与右边的组分融为一个组分,说明猴子的活动范围变广,有利于猴群的基因交流。
4 讨论由于人类干扰,尤其是土地利用变化所造成的生境破碎化问题越来越引起人们的注意,生境的消失和破碎化已成为全球生物多样性的最大威胁。景观连接度的研究对于人们了解物种的扩散制定保护战略具有较大的参考价值。
本文正是基于土地利用变化中,受人为干扰影响最大的农牧地作为研究对象,在不同尺度上定量分析农牧地斑块对滇金丝猴栖息地景观连接度的影响,具有一定的理论价值与实际应用价值。在景观尺度和斑块尺度上,首次使用图论法,运用Graphab1.2软件,计算获得景观连接指数PC定量分析滇金丝猴栖息地景观连接度和组分的连接度,确定了景观功能连接的最佳阈值为1600,在该阈值下,不同的改善情景对于景观连接度均有不同变化。在景观尺度上,PC指数逐渐增加,意味着未来4种不同的农牧地改善情景(C1,C2,C3,C4)都将促进景观连接度的增加;在斑块尺度下,基于距离阈值1600,将整个研究区域分成不同的组分,研究区域中的5个不同管理单元间的猴群被划分在3个不同的组分中,情景C1,C2中组分数量没有变化,意味着连接度没有发生改变,但在情景C3,C4中,组分数量减少,并与相邻的组分融为一个组分,意味着连接度发生改变,滇金丝猴的活动范围扩大。通过不同的情景假设,发现在廊道缓冲区内,优先选择南部的农牧地进行退耕还林,并且优化为次适宜生境,在有限的资源配置下,能减少整个分布区域内的组分数量提高滇金丝猴栖息地的景观连接度有利于滇金丝猴的扩散与基因交流。
本文的研究意义在于(1)根据滇金丝猴的最小家域14.7km2划分了景观连接廊道的缓冲区距离为2.15km,相比其他廊道缓冲区的计算更为客观,也更具有实际应用价值。(2)首次以滇金丝猴栖息地的遗传距离和最小费用距离作为评估相邻两个管理单元间滇金丝猴可能扩散的最小费用距离阈值为1600,该阈值能应用在未来栖息地的规划和保护中。(3)本文分析是基于图论的Graphab1.2上的研究,Graphab1.2是一款应用景观地图进行生态网络建模的软件,具有创建景观连接地图、计算连接度指数、分析连接度指数对物种分布的影响等功能,并拥有制图接口。在此软件中计算PC指数,不同于整体连通性指数(integral index of connectivity,IIC),IIC是基于二位连接模型,即景观中的两个斑块只有连接或不连接两种情况。PC是基于可能性模型,可能性指生境斑块之间连通的可能性,这种可能性与斑块之间的距离呈负相关关系,既可反映整体景观的连通性,又可以计算各组分的景观连接度和各斑块对景观连通性的重要值。相对于国内大多对Confor Sensinode2.2的应用,丰富了景观连接度研究中软件的多元化。
研究区域包含云南境内和西藏地区15个猴群自然保护区,生物种类极为丰富。本文探究廊道缓冲区内的农牧地对滇金丝猴栖息地连接度的影响,由于时间和数据的限制,环境变量选取较少,不同程度的改善情景中许多现实因素还尚未考虑,比如生境质量、改善农牧地的可行性以及每种斑块特点等,结论的验证仍需长期的监测数据。
| [1] | 傅伯杰, 陈利顶. 景观多样性的类型及其生态意义. 地理学报 , 1996, 51 (5) : 454–462. |
| [2] | 万冬梅, 高玮, 王秋雨, 王海涛, 刘明玉. 生境破碎化对丹顶鹤巢位选择的影响. 应用生态学报 , 2002, 13 (5) : 581–584. |
| [3] | Herkert J R, Reinking D L, Wiedenfeld D A, Winter M, Zimmerman J L, Finck Er J, Koford R R, Wolfe D H, Sherrod S K, Alan Jenkins M, Faaborg J, Robinson S K. Effects of Prairie Fragmentation on the Nest Success of Breeding Birds in the Midcontinental United States. Conservation Biology , 2003, 17 (2) : 587–593. DOI:10.1046/j.1523-1739.2003.01418.x |
| [4] | 王应祥, 蒋学龙, 冯庆. 中国叶猴类的分类、现状与保护. 动物学研究 , 1999, 20 (4) : 306–315. |
| [5] | Tischendorf L, Fahrig L. On the usage and measurement of landscape connectivity. Oikos , 2000, 90 (1) : 7–19. DOI:10.1034/j.1600-0706.2000.900102.x |
| [6] | Fahrig L. Effects of habitat fragmentation on biodiversity. Annual Review of Ecology, Evolution,and Systematics , 2003, 34 : 487–515. DOI:10.1146/annurev.ecolsys.34.011802.132419 |
| [7] | Urban D, Keitt T. Landscape connectivity:a graph-theoretic respective. Ecology , 2001, 82 (5) : 1205–1218. DOI:10.1890/0012-9658(2001)082[1205:LCAGTP]2.0.CO;2 |
| [8] | 陈利顶, 傅俊杰. 景观连接度的生态学意义及其应用. 生态学杂志 , 1996, 15 (4) : 37–42. |
| [9] | Minor E S, Urban D L. A graph-theory framework for evaluating landscape connectivity and conservation planning. Conservation Biology , 2008, 22 (2) : 297–307. DOI:10.1111/j.1523-1739.2007.00871.x |
| [10] | Arroyo-Rodriguez-V, Fahrig L. Why is a landscape Perspective Important in studies of Primates?. American Journal of Primatology , 2014, 76 (10) : 901–909. DOI:10.1002/ajp.v76.10 |
| [11] | Anzures-Dadda A, Manson R H. Patch- and landscape-scale effects on howler monkey distribution and abundance in rainforest fragments. Animal Conservation , 2007, 10 (1) : 69–76. DOI:10.1111/acv.2007.10.issue-1 |
| [12] | Arroyo-Rodríguez V, Gonz-á-lez-Perez I M, Garmendia A, Sol-à-M, Estrada A. The relative impact of forest patch and landscape attributes on black howler monkey populations in the fragmented Lacandona rainforest,Mexico. Landscape Ecology , 2013, 28 (9) : 1717–1727. DOI:10.1007/s10980-013-9929-2 |
| [13] | Thornton D H, Branch L C, Sunquist M E. Passive sampling effects and landscape location alter associations between species traits and response to fragmentation. Ecological Applications , 2011, 21 (3) : 817–829. DOI:10.1890/10-0549.1 |
| [14] | Chapman C A, Lawes M J, Naughton-Treves L. Primate survival in community-owned forest fragments: are metapopulation models useful amidst intensive use?//Marsh L K, ed. Primates in Fragments , 2003 : 63–78. |
| [15] | Swart J, Lawes M J. The effect of habitat patch connectivity on samango monkey (Cercopithecus mitis) metapopulation persistence. Ecological Modelling , 1996, 93 (1/3) : 57–74. |
| [16] | Cowlishaw G, Dunbar R. Primate Conservation Biology. Chicago: University of Chicago Press, 2000498. |
| [17] | Estrada A, Garber P, Pavelka M S M, Luecke L. New Perspectives in the Study of Mesoamerican Primates: Distribution, Ecology, Behavior and Conservation. International Journal of Primatology , 2007, 28 (2) : 493–495. DOI:10.1007/s10764-007-9130-z |
| [18] | Marsh L K. Primates in Fragments: Ecology and Conservation. New York: Kluwer Academic/Plenum Press, 2003191–209. |
| [19] | Marsh L, Chapman C A. Primates in Fragments: Complexity and Resilience. New York: Springer-Verlag Inc, 2013537. |
| [20] | 吴昌广, 周志翔, 王鹏程, 肖文发, 滕明君. 景观连接度的概念、度量及其应用. 生态学报 , 2010, 30 (7) : 1903–1910. |
| [21] | 龙勇诚, 钟泰, 肖李. 滇金丝猴地理分布、种群数量与相关生态学的研究. 动物学研究 , 1996, 17 (4) : 437–441. |
| [22] | Wong M H G, Li R Q, Xu M, Long Y C. An integrative approach to assessing the potential impacts of climate change on the Yunnan snub-nosed monkey. Biological Conservation , 2013, 158 : 401–409. DOI:10.1016/j.biocon.2012.08.030 |
| [23] | 薛亚东, 李丽, 李迪强, 吴巩胜, 周跃, 吕玺喜. 基于景观遗传学的滇金丝猴栖息地连接度分析. 生态学报 , 2011, 31 (20) : 5886–5893. |
| [24] | 王亚明, 薛亚东, 夏友福. 滇西北滇金丝猴栖息地景观格局分析及其破碎化评价. 林业调查规划 , 2011, 36 (2) : 34–37. |
| [25] | 黎国强, 杨宇明, 肖文. 滇金丝猴栖息地生物多样性及其威胁因素研究. 云南环境科学 , 2006, 25 (4) : 15–18. |
| [26] | Liu Z J, Ben B P, Wu R D, Hao Y L, Wang B S, Wei F W, Long Y C, Li M. The effect of landscape features on population genetic structure in Yunnan snub-nosed monkeys (Rhinopithecus bieti) implies an anthropogenic genetic discontinuity. Molecular Ecology , 2009, 18 (18) : 3831–3846. DOI:10.1111/mec.2009.18.issue-18 |
| [27] | 武瑞东, 周汝良, 龙勇诚, 杜勇, 叶江霞, 魏晓燕. 滇金丝猴适宜栖息地的遥感分析. 遥感信息 , 2005 (6) : 24–28. |
| [28] | 黎国强, 杨宇明, 肖文. 滇金丝猴栖息地植被类型研究. 西部林业科学 , 2007, 36 (1) : 95–98. |
| [29] | Li L, Xue Y D, Wu G S, Li D Q, Giraudoux P. Potential habitat corridors and restoration areas for the black-and-white snub-nosed monkey Rhinopithecus bieti in Yunnan, China. Oryx , 2015, 49 (4) : 719–726. DOI:10.1017/S0030605313001397 |
| [30] | 董全. 线性人工设施、景观破碎化及其生态效应//现代生态学讲座.IV.理论与实践. 北京: 高等教育出版社, 2009270–289. |
| [31] | Saura S, Pascual-Hortal L. A new habitat availability index to integrate connectivity in landscape conservation planning:Comparison with existing indices and application to a case study. Landscape and Urban Planning , 2007, 83 (2/3) : 91–103. |