 2016, Vol. 36
2016, Vol. 36文章信息
- 王小菲, 高文强, 刘建锋, 倪妍妍, 屈璐, 赵秀莲, 杨文娟, 邓云鹏, 江泽平
- WANG Xiaofei, GAO Wenqiang, LIU Jianfeng, NI Yanyan, QU Lu, ZHAO Xiulian, YANG Wenjuan, DENG Yunpeng, JIANG Zeping.
- 不同生境对栓皮栎幼苗光合生理特性的影响
- Effects of habitat change on the photosynthetic characteristics of Quercus variabilis seedlings
- 生态学报[J]. 2016, 36(24): 8062-8070
- Acta Ecologica Sinica[J]. 2016, 36(24): 8062-8070
- http://dx.doi.org/10.5846/stxb201505110960
-
文章历史
- 收稿日期: 2015-05-11
- 网络出版日期: 2016-04-12

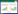




2. 湖北省秭归县林业局磨坪林业站, 秭归 443600
2. Zigui County Forestry Bureau, Moping Forestry Station, Zigui 443600, China
植物与环境之间的关系一直是植物生理生态学研究的热点问题[1-2]。植物的生长不仅受到自身遗传的调控,而且还受环境因素的影响。植物的光合特性可反映植物对光和CO2的利用特点及植物对环境的生长适应性。净光合速率,叶绿素含量以及叶绿素荧光参数作为评价植物生长的主要生理指标,对揭示植物在不同生境下的适应性有着重要的作用[3-6]。交互移植试验是探讨植物局域适应特征,揭示环境与(或)遗传因素对植物性状作用机制的重要手段;也是预测植物在气候变化背景下分布范围变迁的重要研究方法之一[7-8]。
栓皮栎(Quercus variabilis)是东南亚地区天然林分布最广的树种之一,具有较强的适应性,在我国其分布区可北起辽宁,南至云南、广东等地,纬度跨度从25°—40°,气候梯度依次横跨了中温带、暖温带、北亚热带、中亚热带和南亚热带,是我国亚热带、暖温带落叶阔叶林中具有代表性的树种之一[9],具有较大的经济和生态价值。对于栓皮栎种群动态、群落特征等相关研究较多[10-11],包括栓皮栎光合生理的研究也有报道[12-14],而通过对幼苗移栽来分析不同生境下不同种源幼苗其生长及光合特性的研究尚未见报道。本文通过收集3个不同纬度栓皮栎天然群体的种子,育苗后进行南北交互移植,通过对苗期生长及光合特性展开观察和测定,旨在分析栓皮栎不同纬度种源幼苗移栽后的生长状态及光合生理特性,比较各种源对不同生境下的适应性差异,从生理生态学角度进一步探讨这种差异的来源,为将来栓皮栎的引种驯化以及不同环境下的响应方式和适应策略研究提供理论参考。
1 材料与方法 1.1 研究材料2013年5月将盆栽于中国林业科学研究院科研温室的1年生不同种源栓皮栎(云南安宁、陕西黄龙和北京平谷)实生苗移栽至我国南、北界(分别为云南省昆明市西山林场和北京中国林业科学研究院绿化苗圃内空旷地),田间试验采用随机区组设计,每组(即每个种源)20株,3次重复,即每个种源为60株,总计180株,相邻两株之间距离为50 cm。常规管理及自然环境下生长,至当年生长旺季(8月份),对两个不同生境的各种源进行光合生理的测定(包括光合及叶绿素荧光、叶绿素含量的测定),每个种源选择6株生长健康的幼苗进行测定。
北京试验地设在中国林业科学研究院绿化苗圃内空旷地(40°00′9.59″N,116°14′37.66″E,海拔为61 m),该区属于典型的暖温带半湿润大陆性季风气候,冬季寒冷干燥,夏季则炎热多雨,春、秋季较短。试验当年(2013年)年平均气温为12.5 ℃,全年中最高气温为38.2 ℃,最低气温为-14.1 ℃,年降水总量为592 mm,生长季(6—9月)降水量566 mm,年日照时数约2778.7 h,土壤类型为黄棕壤[15]。
昆明试验地设在昆明市西山林场空旷地(25°02′24.67″N,102°37′34.28″E,海拔为2051 m),本区属低纬度高原山地季风气候,雨量适中,气候温和,试验当年(2013年)年平均气温为15.2℃,全年中最高气温为31.0 ℃,最低气温为-6.0 ℃,年降水总量为1001 mm,6—9月降水量506 mm,年日照时数约2200.5 h,土壤类型主要为红壤[15]。
1.2 研究方法 1.2.1 光响应曲线的测定应用便携式光合测定系统(Li-6400,Li-Cor Inc.,Lincoln,USA)进行光响应曲线测定,测定时用 CO2钢瓶对参比室的CO2浓度稳定在400 μmol/mol,叶室温度控制在(25±0.5)℃,相对湿度控制在(40±5)%,光强梯度设定为2000、1500、1200、1000、800、600、400、200、150、100、75、50、25、10、0 μmol m-2s-1,测定前对叶片进行15 min光诱导 (1000 μmol m-2·s-1),每个不同光强值设定数据采集时间3min,测定不同光强下叶片相应的净光合速率 (Pn)。各种源幼苗每组选择生长良好的2株,即每个种源共计6株(6个重复)。于2013年8月下旬晴天的8:30—11:30期间分别在南北试验地实地测定。
1.2.2 CO2响应曲线的测定测定时通过光合仪CO2注入系统控制CO2浓度,使用红蓝光源叶室测定,光强设定为1000 μmol m-2 s-1,温度设定为(25±0.5)℃,CO2浓度梯度为400、200、100、0、50、100、150、200、250、300、400、500、800、1200、1400、1600 μmol/mol,测定不同二氧化碳浓度下栓皮栎的净光合速率(Pn)。测定时间和重复数同上。
1.2.3 叶绿素荧光的测定叶绿素荧光参数采用Mini调制式叶绿素荧光仪(Fluropen-FP100,Photon Systems Instrument,Brno,Czech Republic)进行测定[16],选用的叶片与重复同光响应的测定。测定采用仪器内置NPQ程序并做部分修正:暗适应30 min后,用< 0.1 μmol m-2 s-1的弱光照射测定初始荧光(F0),再照射3000 μmol m-2 s-1(1 s)饱和脉冲光测得最大荧光(Fm),然后活化光(300 μmol m-2 s-1)下进行5次饱和脉冲光(每次间隔12 s)及暗恢复阶段3次饱和脉冲照射(每次间隔26 s)(http://www.psi.cz/)。统计指标主要包括可变荧光(Fv=Fm-F0)、PSⅡ潜在活性(Fv/Fo)、光系统原初光能转换效率(Fv/Fm),光化学猝灭系数PQ=(Fm′-Fs) / (Fm′-F0′)和非光化学猝灭系数NPQ=Fm /Fm′-1[17]。
1.2.4 叶绿素含量的测定使用SPAD-502 Plus叶绿素计(Konica Minolta Optics,Japan)进行测定叶绿素,测定数据为相对叶绿素值,用SPAD表示。对每个测定样株选定两片成熟叶片进行测定,每片叶子选择4—5个点测定计算出一个平均值,每个种源测定6株,共12个重复。
1.2.5 相对生长速率自2013年6月15至9月中旬,对每个种源的所有试验幼苗(60株)进行生长动态的观测,每隔30d用游标卡尺(精确到0.01 mm)和卷尺(精确到0.1 cm)测定基径(mm)和株高(cm),按下式计算相对生长速率[18]:
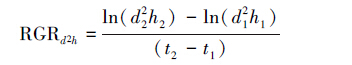
|
(1) |
式中,d1、h1为第1次测定时的基径和株高,d2、h2为第2次测定时的基径和株高,t2-t1为两次测定的时间差(月)。
1.3 数据处理植物光合作用对光和CO2响应测定结果,本文选用直角双曲线修正模型[19-20]进行估算,该模型不仅可以直接计算植物的饱和光强和最大净光合速率,与实测值最为接近,而且解决了低光强条件下和光抑制条件下的光响应问题[19-22]。光响应参数估算的表达式为:
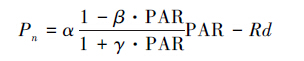
|
(2) |
式中,Pn为净光合速率(μmolCO2 m-2·s-1),α是光响应曲线的初始斜率,也称为表观量子效率AQY(μmol CO2/μmol photons),β和γ为系数(单位为 μmol m-2 s-1),PAR为光合有效辐射(μmol m-2 s-1),Rd为暗呼吸(μmol CO2 m-2 s-1)。当系数β= 0且令γ= α/Pmax,则(2)式将退化为直角双曲线模型[19]。植物的最大净光合速率和光饱和点可通过直角双曲线修正模型直接计算,如下:
最大净光合速率(Pmax)计算方法:
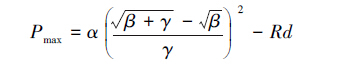
|
(3) |
光饱和点计算方法:
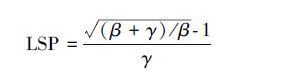
|
(4) |
其次,CO2响应参数估算的表达式[19]为:
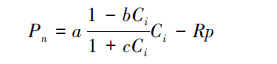
|
(5) |

|
| 图 1 不同种源栓皮栎幼苗在2种生境下的相对生长速率 Fig. 1 Relative growth rate of Quercus variabilis seedlings with various provenances under two habitats 数据为平均值±标准偏差,重复3次(data=Mean±SD,n=3);同一生境下不同大写字母表示不同种源差异达0.05显著水平(LSD法);同一种源不同小写字母表示两种生境下差异达0.05显著水平(T检验) |
式中,Pn为净光合速率,Ci为胞间CO2浓度,a为植物光合作用对CO2响应曲线在Ci=0时的斜率,即CO2响应曲线的初始斜率,也称为初始羧化效率,b和c为系数(mol/μmol),Rp为光呼吸速率(由于光下暗呼吸很小,可以近似将光下叶片向空气中释放CO2的速率看作光呼吸速率)[23],此外,当系数b=0,且c=a/Pmax时,(5)式将退化为直角双曲线模型。通过各系数可计算出植物饱和胞间CO2浓度所对应的光合能力,即最大羧化速率(Amax):
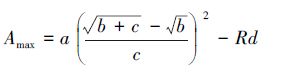
|
(6) |
对上述各统计指标,使用R软件(version 3.0.3 http://www.r-project.org/)进行显著性检验及作图;
采用two-wayANOVA分析生境变化、种源及其交互作用对栓皮栎光合参数的影响。
2 结果与分析 2.1 相对生长速率各种源幼苗南北交互移植3个月后,生长差异明显。如图 1所示,北方生境下3个种源月均相对生长速率显著高于南方生境(表 1)(P<0.05),尤以云南安宁种源生长优势更为突出,相对生长速率最大(0.87),其次是黄龙种源(0.50),北京平谷最小(0.39);而在南方生境下,仅分别为0.15、0.07和0.06。同时,南方种源(安宁和黄龙)移植至北界后,其生长势优于北方本地种源(平谷);而南界种源(安宁)在本地和北界的相对生长速率均显著高于北方种源(黄龙和平谷)(P<0.05)。
| 种源Provenance | 指标Parameters | 北京 Mean±SD | 昆明 Mean±SD |
| 云南安宁 | AQY/(μmol CO2/μmol photons) | 0.07±0.04Aa | 0.08±0.03Aa |
| Pmax/(μmol CO2 m-2 s-1) | 13.99±4.87Aa | 8.23±2.83 Ab | |
| LSP(μmol m-2 s-1) | 1542.13±185.68Ba | 858.65±88.54 Ab | |
| LCP/(μmol m-2 s-1) | 20.66±5.61 Aa | 17.09±10.04Aa | |
| Rd/(μmol CO2 m-2 s-1) | 1.30±0.44Aa | 1.14±0.52Aa | |
| R2 | 0.99 | 0.99 | |
| 陕西黄龙 | AQY/(μmol CO2/μmol photons) | 0.05±0.01Aa | 0.06±0.01Aa |
| Pmax/(μmol CO2 m-2 s-1) | 11.11±1.43Aa | 9.87±10.70 Ab | |
| LSP(μmol m-2 s-1) | 1826.19±114.4Aa | 1077.61±110.7Ab | |
| LCP/(μmol m-2 s-1) | 25.96±1.43 Aa | 10.24±3.8Ab | |
| Rd/(μmol CO2 m-2 s-1) | 1.26±1.43 Aa | 0.83±0.28Aa | |
| R2 | 0.99 | 0.99 | |
| 北京平谷 | AQY/(μmol CO2/μmol photons) | 0.04±0.01 Aa | 0.06±0.02Aa |
| Pmax/(μmol CO2 m-2 s-1) | 11.31±2.80Aa | 7.05±1.22bAa | |
| LSP/(μmol m-2 s-1) | 1794.79±67.63ABa | 1214.45±151.63Ab | |
| LCP/(μmol m-2 s-1) | 24.66±5.32 Aa | 14.35±12.83Ab | |
| Rd/(μmol CO2 m-2 s-1) | 0.99±0.22Aa | 0.64±0.21Aa | |
| R2 | 0.99 | 0.84 | |
| AQY:表观量子效率Apparent quantum yield; Pmax:最大净光合速率Maximumnet photosynthetic rate; LSP:光饱和点Light saturation point; LCP:光补偿点Light compensation point; Rd:暗呼吸速率Dark respiration rate; R2:决定系数Decisive coefficients; 数据为平均值±标准偏差,重复6次;同一行数值带有不同小写字母表示两种生境下差异达0.05显著水平(T检验),同一列同一指标不同大写字母表示同一生境下不同种源差异达0.05显著水平(LSD法) | |||
图 2为南北2种生境下不同栓皮栎种源叶片的光响应曲线(A-PAR)。由图可见,在低光照强度范围内(PAR<400 μmol m-2s-1),不同生境下各种源栓皮栎其净光合速率对光合有效辐射的响应都较为敏感,即净光合速率随着光强的增加而迅速增加,且具有较高的重合度;当光合有效辐射超过400μmol m-2s-1以后,北界(北京)和南界(云南昆明)两个生境下栓皮栎种源表现出明显差别。生长在北界的各种源栓皮栎响应曲线均高于南界,其中来自云南安宁种源,净光合速率在光强为1200μmol m-2s-1期间达到饱和,之后随着光强的增加而降低,表现出明显的光抑制现象(图 2);北京平谷种源在移栽云南以后也表现出光抑制现象(图 2),而陕西黄龙种源其净光合速率随光强(PAR>400 μmol m-2s-1)的增加表现出相对平缓的趋势(图 2)。

|
| 图 2 不同生境下栓皮栎叶片的光响应曲线 Fig. 2 Light response curve of photosynthesis in Quercus variabilis leaves under different habitats 图中,A为云南安宁种源,B为陕西黄龙种源,C北京平谷种源 |
光响应曲线参数拟合结果(表 1)表明,在北界生长的不同种源幼苗,其最大净光合速率(Pmax)和光饱和点(LSP)均显著高于移植至南界的幼苗(P<0.05);除了云南安宁种源外,生长在北界的栓皮栎幼苗其光补偿点(LCP)显著高于南界幼苗;而表观量子效率(AQY)在不同生境不同种源间无显著差异(P>0.05),但生长在南界的各种源其表观量子效率(0.06—0.08)略高于北界的幼苗(0.04—0.07)。
2.3 不同生境下各种源栓皮栎对CO2响应参数的比较图 3为不同生境下各种源栓皮栎叶片净光合速率对CO2响应的拟合曲线(A-Ci)。由图可见,各种源栓皮栎净光合速率随胞间CO2浓度的增加而升高。南、北两个生境下各种源栓皮栎幼苗表现出差异,在北界(北京)生长的3个栓皮栎种源对CO2的响应曲线均高于南界。

|
| 图 3 不同生境下栓皮栎叶片的CO2响应曲线 Fig. 3 Pn-Ci curves in Quercus variabilis leaves under different habitats |
不同生境栓皮栎幼苗Pn-Ci曲线拟合结果表明(表 2),在北界生长的各种源,其羧化效率(CE)和光呼吸速率(Rp)显著高于在南界生长的幼苗(P<0.05),其中又以安宁种源最高,分别达到0.14 mol m-2 s-1和6.39 μmol m-2 s-1。另外,不同生境下各种源CO2补偿点(CCP)的差异均不显著(P>0.05)。
| 种源Provenance | 指标Parameters | 北京 Mean±SD | 昆明 Mean±SD |
| 云南安宁 | CE (mol m-2 s-1) | 0.14±0.06Aa | 0.09±0.02 Ab |
| Amax/(μmol m-2 s-1) | 23.73±1.52Aa | 22.08±7.98Aa | |
| CCP/(μmol mol-1) | 54.55±1.17 Ba | 43.92±14.19Aa | |
| Rp/(μmol m-2 s-1) | 6.39±2.25Aa | 3.87±1.84Ab | |
| R2 | 0.99 | 0.96 | |
| 陕西黄龙 | CE (mol m-2 s-1) | 0.09±0.02ABa | 0.04±0.01Bb |
| Amax/(μmol m-2 s-1) | 23.28±2.04Aa | 18.78±7.62Aa | |
| CCP/(μmol mol-1) | 68.53±4.62 Aa | 65.59±8.45Aa | |
| Rp/(μmol m-2 s-1) | 5.36±0.63Aa | 2.53±0.31ABb | |
| R2 | 0.97 | 0.97 | |
| 北京平谷 | CE (mol m-2 s-1) | 0.06±0.02Ba | 0.03±0.00Bb |
| Amax/(μmol m-2 s-1) | 31.43±7.31 Aa | 6.98±0.84Ab | |
| CCP/(μmol mol-1) | 66.25±5.72 Aa | 52.16±11.17 Aa | |
| Rp/(μmol m-2 s-1) | 3.49±1.05Ba | 1.39±0.34Bb | |
| R2 | 0.99 | 0.73 | |
| CE:羧化效率Carboxylation efficiency; Amax:最大羧化速率Maximumnet photosynthetic rate; CCP:CO2补偿点CO2 compensation point; Rp:光呼吸速率Photo respiration rate; R2:决定系数Decisive coefficients; 数据为平均值±标准偏差,重复6次;同一行数值带有不同小写字母表示两种生境下差异达0.05显著水平(T检验),同一列同一指标不同大写字母表示同一生境下不同种源差异达0.05显著水平(LSD法) | |||
通过对不同生境下3个栓皮栎种源的叶绿素荧光各参数进行分析发现,PSⅡ原初光能转换效率(Fv/Fm)、PSⅡ潜在活性(Fv/Fo)、非光化学猝灭系数(NPQ)等参数在不同生境下无显著差异(P>0.05),而生长在北界的3个栓皮栎种源其叶片光化学猝灭系数(PQ)高于在南界生长的幼苗(P<0.05);其次,在北缘生境下各种源幼苗其荧光差数差异不显著;而在南缘生境下陕西黄龙种源其Fv/Fm显著高于北京平谷和云南安宁种源;北京平谷种源其NPQ最高,Fv/Fo最低,各种源PQ无显著差异。叶绿素相对含量(SPAD值)方面,在北界黄龙种源高于南界(P<0.05),而安宁和平谷种源无显著差异(P>0.05)(表 3)。
| 种源 Provenance | 参数 Parameters | 北京 Mean±SD | 昆明 Mean±SD |
| 云南安宁 | SPAD值 | 40.52±4.94Aa | 40.50±3.85Ba |
| Fv/Fm | 0.80 ±0.02Aa | 0.81 ±0.02Aa | |
| Fv/Fo | 3.99 ±0.54Aa | 4.21 ±0.53Aa | |
| NPQ | 0.86 ±0.22Aa | 0.80 ±0.11Ba | |
| PQ | 0.13 ±0.02Aa | 0.03 ±0.02Ab | |
| 陕西黄龙 | SPAD值 | 39.93±5.87Ab | 48.36±4.92Aa |
| Fv/Fm | 0.78 ±0.03Aa | 0.78 ±0.03ABa | |
| Fv/Fo | 3.67 ±0.69Aa | 3.62 ±0.68ABa | |
| NPQ | 1.02 ±0.27Aa | 0.85 ±0.14Ba | |
| PQ | 0.12 ±0.03Aa | 0.06 ±0.03Ab | |
| 北京平谷 | SPAD值 | 38.29±5.06Aa | 39.04±3.06Ba |
| Fv/Fm | 0.78 ±0.03Aa | 0.72 ±0.08Ba | |
| Fv/Fo | 3.63 ±0.51Aa | 2.79 ±0.92Ba | |
| NPQ | 1.08 ±0.18Aa | 1.13 ±0.21Aa | |
| PQ | 0.12 ±0.02Aa | 0.09 ±0.06Ab | |
| SPAD值:相对叶绿素含量; Fv/Fm:PSⅡ原初光能转换效率primary conversion of light energy of photosystem Ⅱ; Fv/Fo:PSⅡ潜在活性potential activation of photosystem Ⅱ; NPQ:非光化学猝灭系数the non-photochemical quenching coefficient; PQ:光化学猝灭系数photochemical quenching coefficient; 数据为平均值±标准偏差,重复6次;同一行数值带有不同小写字母表示两种生境下差异达0.05显著水平(T检验),同一列同一指标不同大写字母表示同一生境下不同种源差异达0.05显著水平(LSD法) | |||
表 4为生境、种源及其交互作用对栓皮栎光响应参数、CO2响应参数和叶绿素荧光参数影响的双因素方差分析结果。由表可知,生境变化对栓皮栎幼苗的Pmax、Amax和LCP影响显著(P<0.05),以及对LSP、CE、Rp和PQ影响则达到极显著水平(P<0.01);各种源间CCP、Rp、Fv/Fm、NPQ差异显著(P<0.05),CE、LSP和Fv/Fo达极显著差异(P<0.01);同时,生境变化和种源的交互作用对Amax和LSP分别产生了显著和极显著的影响,而AQY、Rd不受生境变化及其与种源的交互影响(P>0.05)。
| 类别 Type | 参数 Parameter | 生境 Habitat | 种源 Provenance | 生境×种源 Habitat×Provenance |
| 光响应参数 | AQY | 0.69 | 3.78 | 0.48 |
| Light response parameters | Pmax | 8.14* | 1.14 | 1.66 |
| LSP | 35.04** | 11.56** | 7.13** | |
| LCP | 4.89* | 0.35 | 0.73 | |
| Rd | 1.65 | 1.66 | 0.04 | |
| CO2响应参数 | CE | 11.52** | 9.05** | 0.40 |
| CO2 response parameters | Amax | 7.74* | 0.36 | 4.16* |
| CCP | 3.51 | 4.40* | 0.46 | |
| Rp | 15.67** | 5.78* | 0.19 | |
| 叶绿素荧光参数 | Fv/Fm | 1.92 | 5.29* | 0.89 |
| Chlorophyll fluorescence parameters | Fv/Fo | 1.13 | 6.19** | 0.69 |
| NPQ | 1.33 | 5.03* | 0.73 | |
| PQ | 26.79** | 1.16 | 1.01 | |
| 相对生长速率Relative growth rate | RGR | 186.94** | 12.86** | 4.68* |
| *,** 分别表示差异达0.05和0.01显著水平; 数据为F值 | ||||
植物的光合作用是一个复杂的生理过程,植物整体的净光合速率或光合潜力与自身生理遗传和环境因子具有密切的关系[24-25]。本研究发现,栽植于北界的3个栓皮栎种源幼苗的Pmax显著高于南界,且以南界种源(安宁)最高,可以认为栓皮栎南方种源较之北方种源具有更高的光合潜能。这与刘建锋等[3]对不同纬度栓皮栎天然实生苗的实地研究结果一致,即南界种源(安宁)具有更高的Pmax。北方(高纬度地区)通常具有更高的日照时数,且试验当年生长季(6—9月)北京降水量高出昆明约60mm,这更有利于自身具有高光合潜能的南方种源(本文中安宁和黄龙种源)进行更多的光合物质生产与积累。这与本次试验中不同种源生长状况(相对生长速率)的分析结果相吻合。
本文发现不同生境下各种源栓皮栎幼苗其表观量子效率在0.04—0.08 μmol CO2/μmol photons之间,与一般植物在适宜生长条件下的表观量子效率(0.03—0.05)基本一致或略有偏高[26-28]。Fv/Fm指PSⅡ原初光能转化效率,在适宜条件下处于0. 75—0. 85之间[29]。本研究中,不同生境下各种源的Fv/Fm 介于或接近上述范围,结合表观量子效率值,可以认为各种源栓皮栎幼苗在南、北生境下均可正常生长。另外,与在南界生境生长相比,在北界生长的幼苗其光饱和点(LSP)和光补偿点(LCP)相对较高,这可能是由于北京地区生长季气温高于昆明地区所致,一般认为温度升高,光补偿点和光饱和点均升高[26]。
荧光淬灭(分为光化学淬灭PQ和非光化学淬灭NPQ)可以作为分析衡量植物在不同生境下PSⅡ的电子传递活性和耗散光能的生理指标。其中,PQ表征PSⅡ天线色素吸收的光能用于光化学传递的份额,PQ值愈大,表明PSⅡ的电子传递活性愈大;而NPQ表征PSⅡ天线色素吸收的光能不能用于光合电子传递而以热的形式耗散掉的光能部分,即植物耗散过剩光能转为热的能力[30]。因此,本文研究结果表明生长在北界的3个栓皮栎种源其叶片光化学猝灭系数(PQ)高于在南界生长的幼苗(P<0.05),表明各种源幼苗在北方生境下具有较大的PSⅡ电子传递活性,即具有较大的光合活性,这与Pmax的结果一致;而NPQ无显著差异,说明在南北不同生境下不同种源幼苗的热耗散或光保护能力相当。
本研究中Pmax、LCP和Amax、CE、Rp及PQ受到生境的显著影响,说明上述生理指标对环境依赖性高于其它生理参数。AQY、CCP、Rd和Fv/Fm、Fv/Fo及NPQ则在不同生境下无显著差异,其中AQY和Rd不受生境变化及其与种源的交互影响,这可能与栓皮栎自身的遗传特征有关。由本文所获结果,可以初步推断南方种源可以较好地适应北方气候(后续观测表明,南方种源在北方能顺利过冬且依然保持生长优势);而北方种源对南方气候适应能力略显不足。按全球暖化导致气候带北移的趋势而言,如果不考虑适应性进化的话,目前栓皮栎将可能在未来气候情景下面临种群缩小的趋势。因此,人工协助栓皮栎南方种源向北定殖与迁移将可能是应对气候暖化,保存优良种质资源的措施之一。
| [1] | Damesin C. Respiration and photosynthesis characteristics of current-year stems of Fagus sylvatica: from the seasonal pattern to an annual balance. New Phytologist , 2003, 158 (3) : 465–475. DOI:10.1046/j.1469-8137.2003.00756.x |
| [2] | Moreno-sotomayor A, Weiss A, Paparozzi E T, Arkebauer T J. Stability of leaf anatomy and light response curves of field grown maize as a function of age and nitrogen status. Journal of Plant Physiology , 2002, 159 (8) : 819–826. DOI:10.1078/0176-1617-00809 |
| [3] | 刘建锋, 肖文发, 熊定鹏, 雷静品, 王鹏程. 不同纬度栓皮栎幼苗生理生态特征的比较. 植物研究 , 2011, 31 (4) : 467–471. |
| [4] | 李永华, 张开明, 于红芳. 10个秋菊品种的光合特性及净光合速率与部分生理生态因子的相关性分析. 植物资源与环境学报 , 2012, 21 (1) : 70–76. |
| [5] | 王建华, 任士福, 史宝胜, 刘炳响, 周玉丽. 遮荫对连翘光合特性和叶绿素荧光参数的影响. 生态学报 , 2011, 31 (7) : 1811–1817. |
| [6] | 黄红英, 窦新永, 孙蓓育, 邓斌, 吴国江, 彭长连. 两种不同生态型麻疯树夏季光合特性的比较. 生态学报 , 2009, 29 (6) : 2861–2867. |
| [7] | ÅgrenJ, SchemskeD W. Reciprocal transplants demonstrate strong adaptive differentiation of the model organism Arabidopsis thaliana in its native range. New Phytologist , 2012, 194 (4) : 1112–1122. |
| [8] | McLean E H, Prober S M, Stock W D, Steane D A, Potts B M, Vaillancourt R E, Byrne M. Plasticity of functional traits varies clinally along a rainfall gradient in Eucalyptus tricarpa. Plant, Cell & Environment , 2014, 37 (6) : 1440–1451. |
| [9] | 张文辉, 卢志军. 栓皮栎种群的生物学生态学特性和地理分布研究. 西北植物学报 , 2002, 22 (5) : 1093–1101. |
| [10] | 吴明作, 刘玉萃. 栓皮栎种群数量动态的谱分析与稳定性. 生态学杂志 , 2000, 19 (4) : 23–26. |
| [11] | 程瑞梅, 肖文发. 河南宝天曼栓皮栎林群落特征及物种多样性. 植物资源与环境 , 1998, 7 (4) : 8–13. |
| [12] | 熊定鹏. 栓皮栎幼苗生理生态特性对环境的响应研究[D]. 武汉: 华中农业大学, 2011. http://cdmd.cnki.com.cn/Article/CDMD-10504-1011405036.htm |
| [13] | 谢会成, 姜志林, 李际红. 栓皮栎林光合特性的研究. 南京林业大学学报:自然科学版 , 2004, 28 (5) : 83–85. |
| [14] | 谢会成, 宋金斗, 姜志林. 栓皮栎林内的光照分布及植物的光合特性研究. 福建林学院学报 , 2004, 24 (1) : 21–24. |
| [15] | 黄昌勇. 土壤学. 北京: 中国农业出版社, 2000235–237. |
| [16] | Samoilova O P, Ptushenko V V, Kuvykin I V, Kiselev S A, Ptushenko O S, Tikhonov A N. Effects of light environment on the induction of chlorophyll fluorescence in Leaves: a comparative study of Tradescantia species of different ecotypes. Bio Systems , 2011, 105 (1) : 41–48. DOI:10.1016/j.biosystems.2011.03.003 |
| [17] | Maxwell K, Johnson G N. Chlorophyll fluorescence-A practical guide. Journal of Experimental Botany , 2000, 51 (345) : 659–668. DOI:10.1093/jexbot/51.345.659 |
| [18] | Kohyama T, Hotta M. Significance of allometry in tropical saplings. Functional Ecology , 1990, 4 (4) : 515–521. DOI:10.2307/2389319 |
| [19] | 叶子飘. 光合作用对光和CO2响应模型的研究进展. 植物生态学报 , 2010, 34 (6) : 727–740. |
| [20] | 叶子飘, 于强. 一个光合作用光响应新模型与传统模型的比较. 沈阳农业大学学报 , 2007, 38 (6) : 771–775. |
| [21] | 吴芹, 张光灿, 裴斌, 徐志强, 赵瑜, 方立东. 不同土壤水分下山杏光合作用CO2响应过程及其模拟. 应用生态学报 , 2013, 24 (6) : 1517–1524. |
| [22] | 陈志成, 王荣荣, 王志伟, 杨吉华, 王华田, 耿兵, 张永涛. 不同土壤水分条件下栾树光合作用的光响应. 中国水土保持科学 , 2012, 10 (3) : 105–110. |
| [23] | Cai S Q, Xu D Q. Relationship between the CO2 compensation point and photorespiration in soybean leaves. Acta Phytophysiologica Sinica , 2000, 26 (6) : 545–550. |
| [24] | 林栋, 吕世海, 冯朝阳, 马晖玲, 刘立成, 赵小强. 华北山地阳坡中生灌草植被对CO2浓度和温度变化的光合响应. 草业科学 , 2008, 25 (4) : 135–140. |
| [25] | 蒋高明, 林光辉, MarinoB D V. 美国生物圈二号内生长在高CO2浓度下的10种植物气孔导度、蒸腾速率及水分利用效率的变化. 植物学报 , 1997, 39 (6) : 546–553. |
| [26] | Sharp R E, Matthews M A, Boyer J S. Kok effect and the Quantum yield of photosynthesis: light partially inhibits dark respiration. Plant Physiology , 1984, 75 (1) : 95–101. DOI:10.1104/pp.75.1.95 |
| [27] | 张淑勇, 周泽福, 夏江宝, 张光灿. 不同土壤水分条件下小叶扶芳藤叶片光合作用对光的响应. 西北植物学报 , 2007, 27 (12) : 2514–2521. |
| [28] | 陈建, 张光灿, 张淑勇, 王梦军. 辽东楤木光合和蒸腾作用对光照和土壤水分的响应过程. 应用生态学报 , 2008, 19 (6) : 1185–1190. |
| [29] | Granda E, Scoffoni C, Rubio-Casal A E, Sack L, Valladares F. Leaf and stem physiological responses to summer and winter extremes of woody species across temperate ecosystems. Oikos , 2014, 123 (11) : 1281–1290. DOI:10.1111/more.2014.123.issue-11 |
| [30] | 郭春芳, 孙云, 唐玉海, 张木清. 水分胁迫对茶树叶片叶绿素荧光特性的影响. 中国生态农业学报 , 2009, 17 (3) : 560–564. |