 2016, Vol. 36
2016, Vol. 36文章信息
- 刘宝贵, 刘霞, 吴瑶, 钟正, 陈宇炜
- LIU Baogui, LIU Xia, WU Yao, ZHONG Zheng, CHEN Yuwei.
- 鄱阳湖浮游甲壳动物群落结构特征
- Spatial and temporal distribution characteristics of planktonic crustaceans in Lake Poyang
- 生态学报[J]. 2016, 36(24): 8205-8213
- Acta Ecologica Sinica[J]. 2016, 36(24): 8205-8213
- http://dx.doi.org/10.5846/stxb201505070941
-
文章历史
- 收稿日期: 2015-05-07
- 网络出版日期: 2016-04-12

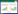




2. 江西省鄱阳湖水利枢纽建设办公室, 南昌 330046;
3. 中国科学院大学, 北京 100049
2. Office of Poyang Lake Water Resources hub Construction, Nanchang 330046, China;
3. University of Chinese Academy of Science, Beijing 100049, China
生境类型对浮游甲壳动物丰度、生物量及群落结构具有十分重要的影响。对富营养化湖泊的研究发现,随着营养水平的改变,湖泊浮游甲壳群落结构和现存量都会随之改变,如浮游甲壳动物丰度变大,剑水蚤增多等[1-2]。同样,大量有关水库或河流的研究表明,水文因素(如流速、流量、含沙量等)对浮游甲壳动物的群落结构亦有十分重要的影响[3-5]。在流速较快的水体中,桡足类的数量高于枝角类的数量并相对保持稳定;而枝角类以小型浮游动物占优势(如象鼻溞、微型裸腹溞等),且丰度随季节波动较大[6-7]。Baranyi[8]等的研究发现,在水体滞留时间短的河流区,象鼻溞是浮游甲壳动物的优势种,其丰度可达总浮游甲壳动物(不包括无节幼体和桡足幼体)的70%以上。
我国最大的淡水湖—鄱阳湖具有“丰水为湖,枯水成河”的特点。即使在同一时期,也同时包含有静水区和流水区。因此,在分析环境因子对鄱阳湖浮游甲壳动物群落结构时空分布影响时,水文因素的影响不容忽略。本文分析了鄱阳湖浮游甲壳动物主要优势类群的季节演替趋势和群落构成特征,旨在阐明水文因素对浮游甲壳动物现存量及群落结构分布的影响。
1 研究方法 1.1 研究区域介绍鄱阳湖位于江西省北部,长江南岸,是中国目前最大的淡水湖。它汇聚赣江、抚河、信江、饶河、修水等五河的来水,经湖口汇入长江。流域面积16.2×105km2,多年平均入江径流量1457×108m3,占长江多年平均径流量的15.6%(大通站)[9-10]。鄱阳湖“五河”的来水于每年的4—6月份达到峰值,7—9月份锐减;每年的10月至次年的2月,流域降水最少[11]。长江的枯水期与鄱阳湖相遇,湖口水位下降,使得鄱阳湖湖水发生了排空,达到最低水位;而长江洪峰则比鄱阳湖稍晚,7月湖口水位的升高,阻塞了鄱阳湖湖水的外泄,使得鄱阳湖达到最高水位。由于其丰、枯水期湖泊水位变动较大,自古就有“丰水一片,枯水一线”和“丰水为湖,枯水为河”等说法![10]。
1.2 采样点设置于2009年1月(枯水期)、4月(涨水期)、7月(丰水期)和10月(退水期)对鄱阳湖浮游甲壳动物现存量及群落结构进行调查分析。调查点(图 1)覆盖了主要湖区及“五河”入湖河口区域。按照丰枯水期地理与水文的变化将研究区域分为三部分。一是常年呈狭窄航道形态的北部湖区,包括L1—L6等6个调查点位;二是丰、枯水期水面大小发生显著变化的南部湖区,主要含L7—L12等6个点位;三是“五河”的7个入湖河口及蚌湖口,包括R1—R8等8个点位。调查点均设置于全年淹水的敞水区(图 1)以保证丰、枯水季均能采集到样品。

|
| 图 1 调查点位(L-lake;R-river) Fig. 1 Location of the sites studied |
浮游甲壳动物的样品采集和分析参照《淡水浮游生物研究方法》进行[12]。采用5L有机玻璃采水器采集表层水样共计10L,再用25#浮游生物网(孔径64μm)过滤后装入50mL塑料瓶中,并用最终体积份数为5%的甲醛固定。带回实验室后,在解剖镜下全部计数鉴定。浮游甲壳动物干重生物量,根据体重-体长回归方程“lgW=blgL+a”计算而得。(式中W为体重,L为体长;不同浮游动物的参数a和b可由参考文献12中查获[12])。
温度、溶解氧、电导率、PH、浊度、矿化度、盐度等指标用Hydrolab(美国)公司生产的DS5X型水质多功能探头于现场进行测定。透明度通过塞氏盘测定。总氮采用过硫酸钾氧化紫外分光光度法测定,总磷采用钼酸铵分光光度法测定。叶绿素a采用热乙醇-分光光度法测定。
1.4 数据分析浮游甲壳动物季节差异性分析,采用单因素方差分析(ANOVA)方法,通过IBM spss20.0 软件包实现。为满足统计分析对数据正态性的需求,部分数据在统计分析时进行了标准化(Y=lg(X+1))处理。生物与非生物因子间相关性大小排序,则采用Canoco for Windows 4. 5进行典范对应分析(CCA:canonical correspondence analysis)。浮游甲壳动物群落结构与丰度的空间分布特征,则借Arcgis10.2软件直接展现在鄱阳湖底图中。
2 分析结果 2.1 现存量季节变化鄱阳湖浮游甲壳动物总丰度呈现出夏季>秋季>春季>冬季的趋势,两两季节之间变化的差异性均达到极显著水平(ANOVA,P<0.001)。夏季浮游甲壳动物的总丰度远远高于其他季节,达到44.59个/L;冬季仅有0.89个/L;春季和秋季的平均丰度分别为1.83个/L和2.90个/L。 除哲水蚤外,其余浮游甲壳动物的数量均以夏季最高,冬季最低。剑水蚤、无节幼体和象鼻溞三者属于终年出现类,且由冬季到秋季其数量先升高后降低,在夏季达到峰值(图 2)。

|
| 图 2 浮游甲壳动物丰度与生物量的季节变化 Fig. 2 Variation of crustaceans abundance and biomass in different seasons |
鄱阳湖枝角类的年均丰度约占浮游甲壳动物总丰度的81%:其中基合溞38%、象鼻溞27%、裸腹溞12%、秀体溞4%。而枝角类的数量,只在夏季高于桡足类,其余季节均以桡足类的数量占优势。冬季,桡足类密度是枝角类密度的6.3倍;而夏季恰恰相反,枝角类的密度是桡足类密度的6.3倍。春季和秋季,桡足类密度分别是枝角类的1.7倍和1.6倍。
鄱阳湖浮游甲壳动物丰度百分比超过10%的优势类群在夏季为象鼻溞、基合溞、裸腹溞和哲水蚤;其余季节皆为无节幼体、象鼻溞、剑水蚤和哲水蚤(图 3)。除夏季外,枝角类均以象鼻溞所占比例最高,分别是冬季95.45%、春季87.31%和秋季91.93%;夏季,象鼻溞属的数量仅占全部枝角类的30.28%,而基合溞、裸腹溞和秀体溞的比例分别为49.96%、14.93%和4.73%。

|
| 图 3 浮游甲壳动物相对丰度与相对生物量的季节变化 Fig. 3 Variation of crustaceans relative abundance and relative biomass in different seasons |
鄱阳湖浮游甲壳动物的生物量的季度变化与密度的季度变化趋势基本一致(图 2)。四季的平均生物量分别为冬季4.72μg/L、春季7.22μg/L、夏季82.07μg/L和秋季9.96μg/L。由于个体之间的大小差异,生物量的构成与密度的构成有巨大的不同(图 3)。枝角类年均生物量约占总生物量的54%,其余46%为桡足类。其中以剑水溞的相对生物量最高占34%,其次依次为象鼻溞21%、基合溞13% 、哲水蚤11%、裸腹溞10%、秀体溞8%、溞1%和无节幼体1%。冬季,桡足类生物量占总生物量的94%,其中哲水蚤占70%,剑水蚤占22%;春季这一比例变为了70%,其中哲水蚤41%、剑水蚤27%;秋季这一比例是80%,其中哲水蚤28%、剑水蚤50%。夏季是枝角类生物量超过桡足类生物量的唯一季节,其群落构成为剑水蚤33%、象鼻溞24%、基合溞17%、裸腹溞13%、秀体溞10%,其余均低于10%。
2.2 空间分布鄱阳湖浮游甲壳动物年均丰度最高的位置在抚河河口(R6),达到85.2个/L。除此之外,湖口(L1)、龙口(R4)、康山(R8)3个点的年均密度也很高分别为31.93个/L、23.53个/L、16.93个/L。年均丰度最低的位置依次为蚌湖口(R3)0.6个/L、修水河河口(R2)0.7个/L、赣江北支河口(R1)1.1个/L。年均密度最高与最低的点均出现在河口或湖口地区,且差别巨大。与此类似,鄱阳湖南北湖区浮游甲壳动物的丰度差异十分显著,年均丰度北部湖区是南部湖区的2.12倍(图 4)。

|
| 图 4 鄱阳湖浮游甲壳动物空间分布特征 Fig. 4 Spatial distribution of planktonic crustaceans in Poyang Lake |
尽管整体看来鄱阳湖浮游甲壳动物的构成均以象鼻溞、基合溞和剑水蚤为主,但不同点位之间的群落构成差异亦十分明显。从图 4可以看出,大部分河口的点和北部入江航道地区(所有以R标注的点及L1—L5)无节幼体、剑水蚤和象鼻溞(灰色表示)所占的比例明显较高;而湖区中部和南部地区(L7—L10)溞和哲水蚤(红色表示)的比例构成明显高于其他地点。主湖区自南向北,溞和哲水蚤的比例有下降趋势;无节幼体、剑水蚤和象鼻溞的比例有增加的趋势;基合溞、裸腹溞和秀体溞(绿色表示)的相对比例则有有先增后减的规律。
2.3 典范对应分析对群落构成与环境因子的CCA分析(图 5)结果表明,前两轴累计解释变化的39.97%。第一轴主要是受温度、叶绿素和营养盐等因素控制;而第二轴代表的是浊度、透明度等物理因素。根据物种间的卡方距离,分析将浮游甲壳动物分为3种不同的类型,可以与不同的生态系统很好的契合。第Ⅰ类是各种生境下均可形成优势类群的无节幼体、象鼻溞、剑水蚤。第Ⅱ类是河流生态系统中经常占优势的裸腹溞、秀体溞和基合溞,他们受温度的影响较大。第Ⅲ类是可以在湖泊静水生态系统中占优势的溞和哲水蚤;他们通常喜好富营养的水体。在鄱阳湖中,Ⅰ类型中的无节幼体,象鼻溞和剑水蚤在一年四季均可检出;而Ⅱ型中基合溞、秀体溞和裸腹溞只在高温、高水位、缓流速的季节出现。从CCA排序图上还可以看出,象鼻溞和剑水蚤离坐标原点较近,丰度与营养盐的关系较密切。说明这两种类群属于广布种,在不同的生境类型下均可形成优势,并喜好富营养的环境,这与表 1的结论一致。

|
| 图 5 环境变量与浮游甲壳动物之间的典范对应分析 Fig. 5 Canonical correspondence analysis between crustaceans and environmental variables TN:总氮 total nitrogen;TP:总磷 total phosphate;TE:温度 temperature;SD:透明度 secchi depth;NTU:浊度 turbidity;Chla:叶绿素-a chlorophyll-a |
| 生境类型 Habitat type | 研究对象 Object | 采样时间与频次 Time & frequency | 优势类群 Dominant taxa | 参考文献 References |
| 河流 River | 多瑙河 | 2006-10—2007-11 每两周一次 | 长额象鼻溞Bosmina longirostris 温剑水蚤Thermocyclops. sp 微型裸腹溞 Moina micrura* | [31] |
| 伊利诺伊河 | 1997—1998年的3月—9月 每两周1次 | 剑水蚤Cyclopoids 无节幼体Nauplii 象鼻溞Bosmina. spp | [32] | |
| 北运河 | 2009—2010 季度采样 | 长额象鼻溞B. longirostris 、 颈沟基合溞Bosminopsis deitersi* 微型裸腹溞 M.micrura* 广布中剑水溞 Mesocyclops leuckarti | [33] | |
| 俄亥俄河 | 2003—2004 5月—10月 | 象鼻溞Bosmina. spp 网纹溞属 Ceriodaphni spp** 秀体溞属 Diaphanosoma.spp* 刺剑水蚤属Diacyclops. spp 中剑水蚤属Mesocyclops.spp | [34] | |
| 乌斯河 | 1989—1993 | 无节幼体Nauplii 象鼻溞Bosmina.spp | [6] | |
| 洛东江 | 1995—1997 每两月1次 | 长额象鼻溞B. longirostris 剑水蚤 Cyclops spp | [35] | |
| 湖泊Lake | Hanebjerg湖 | 剑水蚤目 Cyclopoids | [36] | |
| 武汉东湖 | 1991年 每月1次 | 溞Daphnia ** 秀体溞Diaphanosoma* 广布中剑水蚤 Mesocyclops leuckarti 汤匙华哲水蚤 Sinocalanus dorrii** | [37-38] | |
| 太湖 | 2003.10—2004.9 每月 | (春)透明溞D.hyalina** 哲水蚤**; (夏)简弧象鼻溞B.coregoni 网纹溞Ceriodaphni sp** 剑水蚤Cyclops spp; (秋)简弧象鼻溞B.coregoni; (冬)无节幼体Nauplii | [39] | |
| 梁子湖 | 2006.8—2008.3 月度分析 | (3—4月)透明溞 D.hyalina ** (6—8月)小栉溞D. cristata** (9月)弧象鼻溞B.coregoni (11月)长额象鼻溞B.longirostris | [40] | |
| 南印度湖 | 未提及 | 角突刺剑水蚤Cyclops bicuspidatus thornasi Biaptomus ashlandi 圆形盘肠溞Chydorus sphaericus | [41] | |
| 温尼伯湖 | 1969 6—10月 | 刺剑水蚤 Diacyclops bicuspidatus thomasi Diapmus ashlandi 矮小刺剑水蚤 Acanthocyclops vernalis | [42] | |
| 水库Reservoir | 卡里巴湖 | 1986—1988 | 长额象鼻溞B. longirostris 无节幼体Nauplii 剑水蚤幼体Copepodite cyclopoids | [22] |
| Broa 水库 | 1975 | 哲水蚤Calanoida (most Nauplii) ** 象鼻溞Bosmina | [4] | |
| Arancio湖 | 1990—1991/ 1993 每周1次 | 近剑水蚤Tropocyclops prasinus 长额象鼻溞B.longirostris 短形裸腹溞Moina brachiata* 透明溞D.hyalina** | [29] | |
| 三峡水库 | 2004.4—2005.1 季度采样,仅桡足类 | 北碚中剑水蚤M. pehpeiensis 广布中剑水蚤M.leuckarti 汤匙华哲水蚤Sinocalanus dorrii** | [43] | |
| 格伦大水库 | 1999—2001, 3—6月每周采样 | 透明溞D.pulicaria** 溞属 Daphnia.sp ** 长额象鼻溞B. longirostris | [30] | |
| Saidenbach 水库 | 1976—1988 每周1次 | 盔形溞D. galeata** 冰川真镖水蚤Eudiaptomus gracilis ** 近邻剑水蚤Cyclops vicinus | [44] | |
| *河流生态系统中常见种属;**湖泊生态系统中常见种属 | ||||
温度的升高可以使浮游甲壳动物的繁殖加快,发育期缩短[12-13],进而可使浮游甲壳动物的数量在温暖期达到峰值。与很多研究结果相似[14-16],鄱阳湖浮游甲壳动物的年均密度以夏季最高,冬季最低,秋季和春季居中。但也有研究发现,浮游甲壳动物的数量峰值出现在春季[17]或秋季[18]。王金秋等[19]在1999年对鄱阳湖浮游动物的调查结果显示:枝角类的数量春季(6月)大于秋季(9月),桡足类的数量秋季大于春季。可能与王金秋等[19]调查月份的不同,本文的调查无论是枝角类还是桡足类,秋季(10月)的数量都高于春季(4月)。该结果与谢钦铭等[20] 1988年对鄱阳湖桡足类浮游动物进行了1月1次的调查结果类似:无论是9月还是10月,桡足类的密度都比6月或4月高。
CCA分析(图 5)表明,第一排序轴上温度、叶绿素a、营养盐对大多数浮游甲壳动物季节演替有重要影响。而优势类群与营养盐之间却并未呈现显著的正相关,这与一般的研究结果不同[21-23]。这主要是由于鄱阳湖的特殊水文条件所决定的。枯水期水量小,水体营养盐的浓度最高,而此时正值冬季,温度低、浮游生物的生长发育受到温度限制;除此之外此时的鄱阳湖正好处于河流的形态,流速大、透明度低、固体悬浮物浓度高等条件也不利于浮游动物生长。与之相反,丰水期水量大、流域输入的营养盐被稀释,营养盐的浓度为全年最低;加以此时正值夏季,温度高,生物生长迅速对营养盐吸收作用也较明显;另外,此时的鄱阳湖呈现宽水面的湖泊状态,沿岸区流速慢、透明度高等条件也使得此时的浮游动物丰度达到最高。因此,鄱阳湖浮游甲壳动物与温度、叶绿素a及水位的变化呈现一致,而与营养盐的变化趋势相反。综合以上分析,温度和营养盐对浮游甲壳动物丰度有重要的影响;浮游动物丰度与水体滞留时间呈正相关关系[7-8]。
大量研究表明,流速快的水体中,轮虫和小型甲壳动物如微型裸腹溞、长额象鼻溞,无节幼体或桡足幼体等占优势[6, 14, 24],缓流水体中大型浮游动物,如溞属和哲水蚤的相对生物量会相应增加甚至占优势[14, 25-27];象鼻溞和剑水蚤在缓流水体和激流水体中均可占优势。选取国内外典型的河流、湖泊和水库为对象,对比和总结了3种不同生境下,浮游甲壳动物优势类群的变化(表 1)。结果发现:生境类型而不是营养水平,对浮游甲壳动物群落构成有决定性的作用。河流生态系统中,桡足类的丰度相对稳定,且比例一般高于枝角类,枝角类的数量只偶尔在夏季高于桡足类[6-7];且大型浮游甲壳动物如盘肠溞[28]、僧帽溞、哲水蚤[8]的数量很少,以小型的无节幼体和长额象鼻溞为主。湖泊生态系统中,大型浮游动物如溞属的透明溞、小栉溞、网纹溞,及剑水蚤的部分种类,如中剑水蚤和刺剑水蚤等通常占优势,哲水蚤偶尔占优势。由于水库生态系统同时具有湖泊区和河流区,其浮游甲壳动物群落结构则变化多样,有的水库浮游甲壳动物群落构成具有河流的特征[22],而有的具有湖泊的特征[29],又有的同时具有两种特征[30]。本文的研究对象鄱阳湖,在枯水期呈现河流形态,丰水期呈现湖泊形态;受水位影响,其生境类型在一年中发生显著的转变。
鄱阳湖敞水区浮游甲壳动物优势类群与河流生态系统[8]或水库生态系统的河流区[30]的优势类群具有极大的相似性。比如,除丰水期外,桡足类的数量均显著高于枝角类;桡足类中,无节幼体和桡足幼体的数量通常占优势;枝角类以小型个体占优势等。
3.2 空间分布影响浮游甲壳动物空间分布的因素,概括的可以分为生物因素[10, 45-47]和非生物因素[26, 48]。淡水水域,食物的质量与数量是影响浮游甲壳动物空间分布的最直接因素[12, 49],除此之外流速、悬浮沉积物、藻类的种类与数量等因素对浮游甲壳生物的空间分布也有重要影响[15, 27, 48, 50]。鄱阳湖浮游甲壳动物的年均密度较高的位置位于抚河河口、湖口、饶河河口(龙口)以及康山(抚河、信江、赣江南支在此处交汇,共同汇入鄱阳湖)。密度较低的点则是修水河河口、赣江北支河口以及二者汇流之后的蚌湖湖口。丰度最高与最低的点都出现在河口或湖口区域。因此我们推测,五河来水的水质、水文条件同样是导致河口区域,浮游甲壳动物的生物量相差巨大的主要原因。
鄱阳湖浮游甲壳动物的空间分布规律,再次证明了生境类型对群落构成的重要影响。具有河流生态系统特征的区位包括五河入湖河口和北部入江航道;具有湖泊生态系统特征的区域则主要以鄱阳湖南部开阔湖区为主。与CCA排序分析(图 5)及表 1的结论一致:鄱阳湖浮游甲壳动物群落空间分布显著受到生境类型的控制。
4 结语鄱阳湖浮游甲壳动物优势类群与河流生境下的优势类群有很大的相似性;群落构成及季节演替规律也具有河流生境的特征。在温度、水文等因素共同主导下,鄱阳湖浮游甲壳动物群落发生季节演替,丰、枯水期群落构成和现存量的变化极其显著。同时,不同水系水文、水质条件的不同导致鄱阳湖浮游甲壳动物空间上具有显著的异质性。
| [1] | Pace M L. An empirical analysis of zooplankton community size structure across lake trophic gradients. Limnology and Oceanography , 1986, 31 (1) : 45–55. DOI:10.4319/lo.1986.31.1.0045 |
| [2] | Brett M T, Müller-Navarra D C, Sang-Kyu P. Empirical analysis of the effect of phosphorus limitation on algal food quality for freshwater zooplankton. Limnology and Oceanography , 2000, 45 (7) : 1564–1575. DOI:10.4319/lo.2000.45.7.1564 |
| [3] | Marques S C, Azeiteiro U M, Marques J C, Neto J M, Pardal M Â. Zooplankton and ichthyoplankton communities in a temperate estuary: spatial and temporal patterns. Journal of Plankton Research , 2006, 28 (3) : 297–312. |
| [4] | Bini L M, Tundisi J G, Matsumura-Tundisi T, Matheus C E. Spatial variation of zooplankton groups in a tropical reservoir (Broa Reservoir, São Paulo State-Brazil). Hydrobiologia , 1997, 357 (1/3) : 89–98. DOI:10.1023/A:1003178618304 |
| [5] | Horn W. Long-term development of the crustacean plankton in the Saidenbach Reservoir (Germany) -changes, causes, consequences. Hydrobiologia , 2003, 504 (1/3) : 185–192. |
| [6] | Bass J A B, Pinder L C V, Leach D V. Temporal and spatial variation in zooplankton populations in the River Great Ouse: an ephemeral food resource for larval and juvenile fish. Regulated Rivers: Research & Management , 1997, 13 (3) : 245–258. |
| [7] | Basu B K, Pick F R. Phytoplankton and zooplankton development in a lowland, temperate river. Journal of Plankton Research , 1997, 19 (2) : 237–253. DOI:10.1093/plankt/19.2.237 |
| [8] | Baranyi C, Hein T, Holarek C, Keckeis S, Schiemer F. Zooplankton biomass and community structure in a Danube River floodplain system: effects of hydrology. Freshwater Biology , 2002, 47 (3) : 473–482. DOI:10.1046/j.1365-2427.2002.00822.x |
| [9] | Ye X C, Zhang Q, Liu J, Li X H, Xu C Y. Distinguishing the relative impacts of climate change and human activities on variation of streamflow in the Poyang Lake catchment, China. Journal of Hydrology , 2013, 494 : 83–95. DOI:10.1016/j.jhydrol.2013.04.036 |
| [10] | 朱海虹, 张本. 鄱阳湖: 水文-生物-沉积-湿地-开发整治. 合肥: 中国科学技术大学出版社, 199745–67. |
| [11] | Gao J H, Jia J J, Kettner A J, Xing F, Wang Y P, Xu X N, Yang Y, Zou X Q, Gao S, Qi S H, Liao F Q. Changes in water and sediment exchange between the Changjiang River and Poyang Lake under natural and anthropogenic conditions, China. Science of the Total Environment , 2014, 481 : 542–553. DOI:10.1016/j.scitotenv.2014.02.087 |
| [12] | 章宗涉, 黄祥飞. 淡水浮游生物研究方法. 北京: 科学出版社, 1995147–158. |
| [13] | 郑重. 淡水枝角类的生殖. 动物学杂志 , 1959 (1) : 22–28. |
| [14] | Thorp J H, Black A R, Haag K H, Wehr J D. Zooplankton assemblages in the Ohio River: seasonal, tributary, and navigation dam effects. Canadian Journal of Fisheries and Aquatic Sciences , 1994, 51 (7) : 1634–1643. DOI:10.1139/f94-164 |
| [15] | Gaughan D J, Potter I C. Composition, distribution and seasonal abundance of zooplankton in a shallow, seasonally closed estuary in temperate Australia. Estuarine, Coastal and Shelf Science , 1995, 41 (2) : 117–135. DOI:10.1006/ecss.1995.0057 |
| [16] | Froneman P W. Seasonal changes in zooplankton biomass and grazing in a temperate estuary, South Africa. Estuarine, Coastal and Shelf Science , 2001, 52 (5) : 543–553. DOI:10.1006/ecss.2001.0776 |
| [17] | 冯坤, 万成炎, 彭建华, 张志永, 郑志伟, 胡莲, 邹曦. 三峡库区26条支流浮游甲壳动物的群落结构. 水生态学杂志 , 2012, 33 (4) : 40–48. |
| [18] | 冯坤, 彭建华, 万成炎, 胡莲, 邹曦, 张志永. 三道河水库浮游甲壳动物群落结构特征的初步研究. 水生态学杂志 , 2010, 3 (3) : 18–22. |
| [19] | 王金秋, 吴建平, 於燕斌, 王天宇, 成功. 春秋两季鄱阳湖浮游动物的编目、数量分布与变动. 湖泊科学 , 2003, 15 (4) : 345–352. |
| [20] | 谢钦铭, 李长春. 鄱阳湖桡足类的群落组成与现存量季节变动的初步研究. 江西科学 , 1998, 16 (3) : 180–187. |
| [21] | Bothár A, Kiss K T. Phytoplankton and zooplankton (Cladocera, Copepoda) relationship in the eutrophicated River Danube (Danubialia Hungarica, CXI). Hydrobiologia , 1990, 191 (1) : 165–171. DOI:10.1007/BF00026050 |
| [22] | Masundire H M. Spatial and temporal variations in the composition and density of crustacean plankton in the five basins of Lake Kariba, Zambia-Zimbabwe. Journal of Plankton Research , 1997, 19 (1) : 43–62. DOI:10.1093/plankt/19.1.43 |
| [23] | Chen F Z, Tang H J, Shu T T, Wang W X, Zhou L B. Effects of Microcystis blooms on the crustacean plankton community: enclosure experiments in a subtropical lake. Hydrobiologia , 2013, 711 (1) : 175–185. DOI:10.1007/s10750-013-1480-2 |
| [24] | Bøhn T, Amundsen P A. Effects of invading vendace (Coregonus albula L.) on species composition and body size in two zooplankton communities of the Pasvik River System, northern Norway. Journal of Plankton Research , 1998, 20 (2) : 243–256. DOI:10.1093/plankt/20.2.243 |
| [25] | 林秋奇. 流溪河水库后生浮游动物多样性与群落结构的时空异质性[D]. 广州: 暨南大学, 2007. http://cdmd.cnki.com.cn/Article/CDMD-10559-2007192507.htm |
| [26] | Betsill R K, van den Avyle M J. Spatial heterogeneity of reservoir zooplankton: a matter of timing?. Hydrobiologia , 1994, 277 (1) : 63–70. DOI:10.1007/BF00023986 |
| [27] | Branco C W, Rocha M I A, Pinto G F S, Gômara G A, Filippo R D. Limnological features of Funil Reservoir (R.J., Brazil) and indicator properties of rotifers and cladocerans of the zooplankton community. Lakes & Reservoirs: Research & Management , 2002, 7 (2) : 87–92. |
| [28] | Vijverberg J, Boersma M. Long-term dynamics of small-bodied and large-bodied cladocerans during the eutrophication of a shallow reservoir, with special attention for Chydorus sphaericus. Hydrobiologia , 1997, 360 (1/3) : 233–242. DOI:10.1023/A:1003148600983 |
| [29] | Naselli-Flores L, Barone R. Importance of water-level fluctuation on population dynamics of cladocerans in a hypertrophic reservoir (Lake Arancio, south-west Sicily, Italy). Hydrobiologia , 1997, 360 (1) : 223–232. |
| [30] | Bernot R J, Dodds W K, Quist M C, Guy C S. Spatial and temporal variability of zooplankton in a great plains reservoir. Hydrobiologia , 2004, 525 (1/3) : 101–112. |
| [31] | Vadadi-Fülöp C, Hufnagel L, Jablonszky G, Zsuga K. Crustacean plankton abundance in the Danube River and in its side arms in Hungary. Biologia , 2009, 64 (6) : 1184–1195. |
| [32] | Wahl D H, Goodrich J, Nannini M A, Dettmers J M, Soluk D A. Exploring riverine zooplankton in three habitats of the Illinois River ecosystem: Where do they come from?. Limnology and Oceanography , 2008, 53 (6) : 2583–2593. DOI:10.4319/lo.2008.53.6.2583 |
| [33] | 李学军, 鲍战猛, 高彩凤, 毛战坡. 北运河浮游动物调查及水质评价. 生态学杂志 , 2014, 33 (6) : 1559–1564. |
| [34] | Thorp J H, Mantovani S. Zooplankton of turbid and hydrologically dynamic prairie rivers. Freshwater Biology , 2005, 50 (9) : 1474–1491. DOI:10.1111/fwb.2005.50.issue-9 |
| [35] | Kim H W, Joo G J. The longitudinal distribution and community dynamics of zooplankton in a regulated large river: a case study of the Nakdong River (Korea). Hydrobiologia , 2000, 438 (1/3) : 171–184. DOI:10.1023/A:1004185216043 |
| [36] | Romare P, Berg S, Lauridsen T, Jeppesen E. Spatial and temporal distribution of fish and zooplankton in a shallow lake. Freshwater Biology , 2003, 48 (8) : 1353–1362. DOI:10.1046/j.1365-2427.2003.01081.x |
| [37] | Yang Y F, Huang X F, Liu J K, Jiao N Z. Effects of fish stocking on the zooplankton community structure in a shallow lake in China. Fisheries Management and Ecology , 2005, 12 (2) : 81–89. DOI:10.1111/fme.2005.12.issue-2 |
| [38] | 杨宇峰, 黄祥飞. 武汉东湖浮游动物群落结构的研究. 应用生态学报 , 1994, 5 (3) : 319–324. |
| [39] | 杨桂军, 潘宏凯, 刘正文, 王卫民, 秦伯强. 太湖不同湖区浮游甲壳动物季节变化的比较. 中国环境科学 , 2008, 28 (1) : 27–32. |
| [40] | 许隆君, 陆鑫歆, 王忠锁. 梁子湖浮游甲壳动物的生物多样性. 生态学报 , 2009, 29 (12) : 6419–6428. |
| [41] | Patalas K, Salki A. Effects of impoundment and diversion on the crustacean plankton of Southern Indian Lake. Canadian Journal of Fisheries and Aquatic Sciences , 1984, 41 (4) : 613–637. DOI:10.1139/f84-074 |
| [42] | Patalas K, Salki A. Crustacean Plankton in Lake Winnipeg-variation in space and time as a function of lake morphology, geology, and climate. Canadian Journal of Fisheries and Aquatic Sciences , 1992, 49 (5) : 1035–1059. DOI:10.1139/f92-116 |
| [43] | 姚建良, 薛俊增, 王登元, 蔡庆华, 黄祥飞, 刘建康. 三峡水库初次蓄水后干流库区桡足类的纵向分布与季节变化. 生物多样性 , 2007, 15 (3) : 300–305. |
| [44] | Horn W. The influence of biomass and structure of the crustacean plankton on the water transparency in the Saidenbach storage reservoir. Hydrobiologia , 1991, 225 (1) : 115–120. DOI:10.1007/BF00028390 |
| [45] | 潘继征, 熊飞, 李文朝, 李爱权. 抚仙湖浮游甲壳动物群落结构与空间分布. 湖泊科学 , 2009, 21 (3) : 408–414. |
| [46] | 杨宇峰, 王庆, 陈菊芳, 庞世勋. 河口浮游动物生态学研究进展. 生态学报 , 2006, 26 (2) : 576–585. |
| [47] | 杨宇峰, 黄祥飞. 鲢鳙对浮游动物群落结构的影响. 湖泊科学 , 1992, 4 (3) : 78–86. |
| [48] | Matsumura-Tundisi T, Tundisi J G. Calanoida (Copepoda) species composition changes in the reservoirs of São Paulo State (Brazil) in the last twenty years. Hydrobiologia , 2003, 504 (1/3) : 215–222. |
| [49] | 蒋燮治, 堵南山. 中国动物志-节肢动物门 甲壳纲 淡水枝角类. 北京: 科学出版社, 1979. |
| [50] | Davidson Jr N L, Kelso W E, Rutherford D A. Relationships between environmental variables and the abundance of cladocerans and copepods in the Atchafalaya River Basin. Hydrobiologia , 1998, 379 (1/3) : 175–181. DOI:10.1023/A:1003488332055 |