 2016, Vol. 36
2016, Vol. 36文章信息
- 龚堃, 鲍毅新, 任鹏, 方平福, 李海宏, 王华
- GONG Kun, BAO Yixin, REN Peng, FANG Pingfu, LI Haihong, WANG Hua.
- 漩门湾围垦区外滩涂不同生境大型底栖动物功能群及其变化
- Macrobenthic functional groups in different habitats in the reclaimed wetland Xuanmen Bay
- 生态学报[J]. 2016, 36(24): 8214-8223
- Acta Ecologica Sinica[J]. 2016, 36(24): 8214-8223
- http://dx.doi.org/10.5846/stxb201504260860
-
文章历史
- 收稿日期: 2015-04-26
- 网络出版日期: 2016-04-12

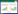




围垦能有效增加土地资源,拓展人类生产生活的范围,同时带来可观的经济效益,但围垦也将对潮间带原有的生态环境造成破坏。潮间带生态平衡被打破之后会引发一系列湿地生态问题,如潮间带底质变化、海岸气候变化、生物流失等[1-2],尤其是大型底栖动物群落的变化,越来越受到国内外学者的关注[3-4],滩涂湿地生态系统的结构和功能都会因围垦而发生改变[5]。大型底栖动物是湿地生物的重要类群,在生态系统中发挥重要作用。大型底栖动物是次级生产者,参与食物网中有机物的分解、物质循环和能量流动、水体净化等过程[6-7]。因而,大型底栖动物的生物多样性、群落结构和物种组成对该区域的生态环境具有一定的指示作用[8],但考虑到底栖动物的自由活动能力和对环境变化的敏感程度等因素,当环境发生变化时,某个或某些物种的迁徙、更替、消失有时并不能准确反映该区域生态系统的变化[9]。
底栖动物功能群是具有相似生态功能的一类底栖动物的集合[10-11],利用这种以生态功能为基础的方法进行分类,而不是纯粹依据分类学的标准对物种进行分类,可以简化种间关系,弱化单一物种的作用。通过将物种划分为简单的几个功能群来研究,使生态系统的复杂性降低[12],这些类群对环境变化的反应相较于个体或种群更为迅速[13],综合性更强,因此更有利于从整体评估生态系统的结构和功能[9, 14]。
目前,国内外学者对大型底栖动物的功能群进行了许多研究[15-19]。其中,国内的有关研究主要集中在潮间带区域[14, 20-23],而对围垦后新形成的滩涂湿地,其功能群的研究较少。本文通过对漩门湾围垦区外3种生境的滩涂湿地大型底栖动物进行连续两周年的采样调查,分析不同生境功能群的生物多样性及其季节和周年变化,为漩门湾湿地资源的合理利用与保护,尤其是围垦湿地生态系统的恢复和重建提供理论依据,亦为深入研究围垦对滩涂湿地大型底栖动物群落变化的影响机制积累资料。
1 材料与方法 1.1 研究区概况漩门湾位于浙江省台州市玉环县,总面积31.48 km2,属亚热带季风气候,年平均气温为16.9—17.6℃,年降水量为1300—1400 mm,是浙江省沿海地区典型的湿地类型。1998年10月漩门湾二期围垦工程开工建设,至2005年12月完成,此时新形成的潮间带没有明显的高、中、低潮间带区别,潮带宽度约为250—300 m,土质属粘涂土种,土体尚未分化,呈母质状,为最新浅海沉积物。表层平均有机质含量为1.38%,全氮0.090%、碱解氮45×10-6、速效磷1030×10-6、pH值8.2—8.4。调查的潮间带包括互花米草草滩(HH)、自然滩涂光滩(GT)和人工红树林泥滩(HS) 3种生境。其中互花米草(Spartina alterniflora)入侵较早,所形成的互花米草草滩大多已发育成熟,具有潮沟地貌,调查区内互花米草盖度为25%—35%;秋茄红树(Kandelia candel)的种植始于2010年,所引种的红树,由于受到当年冬季雪灾的影响,长势不良,第2年后逐渐得到恢复。
1.2 样品采集与分析2010年10月至2012年7月,每年10月(秋季)、1月(冬季)、4月(春季)、7月(夏季),分别对3种生境(HH、GT和HS)共进行8个季度的大型底栖动物调查。3种生境各选2条与海岸线垂直的带状断面,各断面布设5个样点(图 1),在最低潮时,用25 cm×25 cm×30 cm规格的定量采样器采集泥样,采样深度为20 cm;上述每个样点均选取4个样方,总计120个样方。

|
| 图 1 漩门湾围垦湿地大型底栖动物采样示意图 Fig. 1 Sketch map of sampling sites of macrobenthos in the reclaimed wetland Xuanmen Bay |
用1 mm孔径的分样筛对所采泥样进行淘洗,获得的标本用10%福尔马林固定,带回实验室后进行分类计数,而后将标本放入烘箱中,在65℃下烘干至恒量,用电子天平(精确到0.0001g)对标本进行称量,记录数据。采样时对偶见种和不易用采样器采集的物种进行定性取样,结果分析时将定量和定性调查综合考虑。调查方法和样品处理按《海洋调查规范》第六部分:海洋生物调查(GB12763.6—2007)进行。
1.3 功能群划分本文依据食性特点将大型底栖动物划分为5个功能群[14-15, 21]:浮游生物食者(Pl)、植食者(Ph)、肉食者(C)、杂食者(O)和碎屑食者(D)。
1.4 数据处理将所得数据运用Excel、SPSS20.0、PRIMER5.0软件统计,参照以下公式对取样所得的数据进行分析:
Margalef丰富度指数S=(S-1)/lnN
Shanno-Wiener多样性指数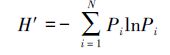
Pielou均匀度指数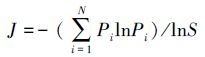
式中,S为总物种数,N为总个体数;Pi为第i个物种出现的频率。
2 结果 2.1 功能群组成5个功能群的组成和数量见表 1。第1周年,HH、GT和HS生境下属于Pl功能群的均为5种,定量取样获得的物种数分别为5、4和5,其中HH代表种为焦河蓝蛤(Potamocorbula ustulata)和彩虹明樱蛤(Moerella iridescen),而GT和HS代表种均为焦河蓝蛤和短拟沼螺(Assiminea brevicula);Ph功能群均有12种,定量取样获得的物种数分别为8、6和6,HH代表种为珠带拟蟹守螺(Cerithidea cingulata),GT代表种为珠带拟蟹守螺、伍氏厚蟹(Helice Wuana)和弧边招潮(Uca arcuata),而HS代表种为珠带拟蟹守螺和泥蟹(Ilyoplax sp.);C功能群均有14种,定量取样获得的物种数分别为9、7和7,HH和GT代表种均为婆罗囊螺(Retusa boenesis);O功能群均有11种,定量取样获得的物种数分别为3、3和4;D功能群均有7种,定量取样获得的物种数分别为4、2和2。需要指出的是,O和D功能群所有物种的个体数均少于总个体数的1%。第2周年,HH、GT和HS生境下属于Pl功能群的均为5种,定量取样获得的物种数分别为3、3和2,HH代表种为短拟沼螺,而GT和HS代表种除了短拟沼螺,还有焦河蓝蛤;Ph功能群均有12种,定量取样获得的物种数分别为9、8和8,HH代表种为珠带拟蟹守螺,GT代表种为泥蟹,HS代表种为珠带拟蟹守螺和泥蟹;C功能群均有14种,定量取样获得的物种数分别为6、9和10,HH所有物种的个体数均小于总个体数的1%,而GT代表种为半褶织纹螺(Nassarius semiplicatus),HS代表种为婆罗囊螺和习见织纹螺(Nassarius dealbatus);O功能群均有11种,定量取样获得的物种数分别为2、7和5,且所有物种的个体数均小于总个体数的1%;D功能群均有7种,定量取样获得的物种数分别为6、6和3,HH代表种为可口革囊星虫(Phascolosoma esculenda)。
| 功能群 Functional groups | 种类 Species | HH中的数量Number | GT中的数量Number | HS中的数量Number | |||
| 第1周年 1st year | 第2周年 2nd year | 第1周年 1st year | 第2周年 2nd year | 第1周年 1st year | 第2周年 2nd year | ||
| 肉食者 | 婆罗囊螺 Retusa boenesis | ++ | — | ++ | + | + | ++ |
| Camivorous group,C | 宽带梯螺 Epitonium clementinum | — | + | + | + | — | — |
| 红带织纹螺 Nassarius succinctus | + | + | + | + | + | + | |
| 习见织纹螺 N. dealbatus | + | — | + | + | + | ++ | |
| 半褶织纹螺 N. semiplicatus | + | + | + | ++ | + | + | |
| 秀丽织纹螺N. dealbatus | — | — | — | — | — | — | |
| 丽核螺 Pyrene bella | + | — | — | — | — | — | |
| 斑玉螺 Natica tigrina | + | — | + | — | — | + | |
| 微黄镰玉螺 Lunatia gilva | + | + | + | + | — | + | |
| 日本蟳 Charybdis japonica | — | — | — | + | + | + | |
| 长吻沙蚕 Glycera chirori | — | + | — | + | — | + | |
| 日本刺沙蚕 Neanthes japonica | + | — | — | — | + | + | |
| 日本沙蚕 Nereis japonica | + | + | — | + | + | + | |
| 纽虫Cerebratulina sp. | — | — | — | — | — | — | |
| 碎屑食者 | 泥蚶 Tegillarca granosa | + | + | + | + | + | + |
| Detritivorous group,D | 褐蚶 Didimacar tenebrica | + | + | + | + | + | + |
| 黑荞麦蛤 Vignadula atrata | — | + | — | + | — | — | |
| 裸盲蟹 Typhlocarcinus nudus | — | — | — | + | — | — | |
| 锥头虫 Gaudryina sp. | + | + | — | — | — | — | |
| 小头虫Capitella capitata | — | + | — | + | — | + | |
| 可口革囊星虫 Phascolosoma esculenda | + | ++ | — | + | — | — | |
| 杂食者 | 石磺 Onchidium verruculatum | — | + | — | + | — | + |
| Omnivorous group,O | 豆形拳蟹 Philyra pisum | — | — | + | — | — | — |
| 橄榄拳蟹 P. olivacea | — | — | — | + | — | — | |
| 中华长眼寄居蟹 Paguristes sinensis | — | — | — | + | — | + | |
| 长腕寄居蟹 Pagurus geminus | — | — | — | + | — | — | |
| 长指寄居蟹 P. dubius | — | — | — | + | — | — | |
| 中国毛虾 Acetes chinensis | — | — | — | + | + | — | |
| 日本鼓虾 Alpheus japonicus | + | — | — | — | + | + | |
| 安氏白虾 Exopalaemon annandalei | + | — | + | — | + | + | |
| 秀丽白虾 Exopalaemon modestus | — | — | — | — | — | — | |
| 泥螺 Bullacta exarata | + | + | + | + | + | + | |
| 浮游生物食者 | 缢蛏 Sinonovacula constricta | + | + | + | — | + | — |
| Planktophagous group, | 小刀蛏 Cultellus attenuatus | + | — | — | — | + | — |
| Pl | 焦河蓝蛤 Potamocorbula ustulata | +++ | — | +++ | +++ | +++ | ++ |
| 彩虹明樱蛤 Moerella iridescen | ++ | + | + | + | + | — | |
| 短拟沼螺 Assiminea brevicula | + | +++ | +++ | +++ | +++ | +++ | |
| 植食者 | 珠带拟蟹守螺 Cerithidea cingulata | +++ | +++ | ++ | ++ | ++ | +++ |
| Phytophagous group, | 尖锥拟蟹守螺 C. largillierti | + | — | — | — | — | — |
| Ph | 光滑狭口螺 Stenothyra glabra | — | — | — | — | — | — |
| 沈氏厚蟹 Helice sheni | + | ++ | + | ++ | — | + | |
| 中华相手蟹 Sesarma sinensis | — | + | — | — | — | — | |
| 伍氏厚蟹 H. wuana | + | ++ | ++ | ++ | + | ++ | |
| 长足长方蟹 Metaplax longipes | + | + | + | + | + | + | |
| 沈氏长方蟹 Metaplax sheni | — | + | — | + | + | + | |
| 日本大眼蟹 Macrophthalmus japonicus | + | + | + | + | + | + | |
| 弧边招潮 Uca arcuata | + | ++ | — | — | — | + | |
| 泥蟹 Ilyoplax sp. | + | ++ | ++ | +++ | ++ | +++ | |
| 绒螯近方蟹 Hemigrapsus penicillatus | — | — | — | + | — | — | |
| HH: 互花米草 Spartina alterniflora;GT: 光滩 Natural tidal flat;HS: 红树林 Mangrove forests; +++ 该物种个体数占总个体数的10%以上; ++ 该物种个体数占总个体数的; + 该物种个体数占总个体数小于1%; — 定量取样未获得 | |||||||
两周年3种生境均以浮游生物食者Pl和植食者Ph为主导功能群,且平均密度和平均生物量均是Pl>Ph(表 2)。两功能群的密度和生物量均较高,其年平均密度之和在3种生境中均是第1周年>第2周年,两周年平均密度以HS中为最高,GT次之,HH最低;年平均生物量之和在HH和GT中均是第2周年>第1周年,HS中是第1周年>第2周年,两周年平均密度以HS中为最高,HH次之,GT最低。
| 生境 Habitats | 周年 Years | 季节 Seasons | C | D | O | Ph | Pl | |||||
| 密度 Density/ (个/m2) | 生物量 Biomass/ (g/m2) | 密度 Density/ (个/m2) | 生物量 Biomass/ (g/m2) | 密度 Density/ (个/m2) | 生物量 Biomass/ (g/m2) | 密度 Density/ (个/m2) | 生物量 Biomass/ (g/m2) | 密度 Density/ (个/m2) | 生物量 Biomass/ (g/m2) | |||
| HH | 第1周年 | 春 | 37.20 | 3.75 | 2.00 | 3.00 | 3.20 | 0.13 | 74.00 | 21.67 | 609.20 | 11.62 |
| 夏 | 5.60 | 1.91 | 4.00 | 0.81 | 1.60 | 0.17 | 118.40 | 60.84 | 120.40 | 6.63 | ||
| 秋 | 65.60 | 2.45 | 4.40 | 1.59 | — | — | 128.00 | 16.19 | 460.40 | 8.24 | ||
| 冬 | 17.60 | 2.18 | 2.40 | 0.28 | 0.40 | 0.01 | 0.80 | 0.01 | 161.60 | 7.22 | ||
| 第2周年 | 春 | 2.40 | 0.28 | 20.00 | 2.95 | 2.00 | 0.11 | 22.00 | 10.85 | 121.20 | 3.80 | |
| 夏 | 4.00 | 0.55 | 22.40 | 3.79 | 1.60 | 0.09 | 78.40 | 37.37 | 124.80 | 9.49 | ||
| 秋 | 4.80 | 1.05 | 8.80 | 11.42 | — | — | 164.80 | 70.13 | 218.80 | 5.38 | ||
| 冬 | 2.00 | 0.53 | 6.80 | 3.73 | — | — | 68.80 | 20.78 | 113.60 | 2.58 | ||
| GT | 第1周年 | 春 | 44.00 | 4.22 | 2.40 | 0.72 | 1.20 | 0.20 | 19.60 | 1.23 | 3206.40 | 31.46 |
| 夏 | 5.80 | 0.12 | — | — | 0.40 | 0.02 | 101.00 | 7.32 | 316.40 | 15.18 | ||
| 秋 | 12.80 | 0.98 | — | — | — | — | 7.60 | 0.19 | 196.40 | 8.42 | ||
| 冬 | 50.40 | 5.05 | — | — | 0.40 | 0.01 | 52.00 | 12.08 | 236.40 | 9.75 | ||
| 第2周年 | 春 | 12.80 | 2.84 | 3.60 | 3.74 | 2.40 | 0.11 | 94.00 | 6.66 | 273.60 | 16.19 | |
| 夏 | 12.00 | 4.36 | 4.00 | 9.32 | 6.80 | 2.89 | 46.00 | 10.73 | 496.40 | 30.02 | ||
| 秋 | 29.2 | 3.50 | 1.20 | 0.14 | 1.20 | 0.04 | 116.40 | 7.07 | 149.60 | 11.39 | ||
| 冬 | 2.00 | 0.13 | 3.60 | 0.43 | 2.00 | 0.09 | 23.20 | 1.66 | 111.20 | 10.42 | ||
| HS | 第1周年 | 春 | 23.60 | 0.74 | — | — | 1.60 | 0.18 | 64.80 | 2.58 | 3540.40 | 21.63 |
| 夏 | 14.00 | 3.14 | 1.60 | 5.17 | 1.20 | 0.10 | 81.60 | 29.32 | 920.80 | 48.61 | ||
| 秋 | 3.20 | 0.03 | — | — | 0.80 | 0.14 | 148.40 | 51.19 | 205.20 | 6.38 | ||
| 冬 | 9.20 | 1.19 | 1.20 | 2.59 | 0.80 | 0.03 | 50.80 | 2.71 | 315.20 | 13.75 | ||
| 第2周年 | 春 | 12.00 | 6.98 | 2.40 | 8.16 | 2.80 | 0.08 | 73.20 | 24.86 | 29.20 | 0.63 | |
| 夏 | 8.40 | 1.62 | 4.00 | 2.03 | 5.60 | 0.59 | 100.40 | 23.96 | 250.80 | 13.66 | ||
| 秋 | 24.40 | 11.73 | 1.20 | 0.04 | 0.80 | 0.05 | 165.20 | 72.68 | 267.60 | 7.19 | ||
| 冬 | 32.00 | 2.57 | 3.20 | 0.16 | 0.80 | 0.14 | 26.40 | 7.17 | 62.40 | 2.39 | ||
| —数值Value<0.01 | ||||||||||||
3种生境中,不同功能群的密度和生物量在两周年间呈现不同的季节变化规律。功能群密度方面:第1周年HH生境中,C、D和Ph的密度秋季最大,O和Pl春季最大;GT生境中,D、O和Pl春季最大,C和Ph则分别在冬季和夏季最大;HS生境中,C、O和Pl春季最大,D和Ph则分别在夏季和秋季最大。第2周年HH生境中,C、Ph和Pl秋季最大,D和O则分别在夏季和春季最大;GT生境中,D、O和Pl夏季最大,C和Ph在秋季最大;HS生境中,C冬季最大,D和O夏季最大,而Ph和Pl秋季最大。生物量方面:第1周年HH生境中,C、D和Pl的生物量春季最高,O和Ph则夏季最高;GT生境中,D、O和Pl春季最高,C和Ph则夏季最高;HS生境中,C、D和Pl夏季最高,而O和Ph分别在春季和秋季最高。第2周年HH生境中,C、D和Ph秋季最高,而O和Pl则分别在春季和夏季最高;GT生境中,各功能群均在夏季最高;HS生境中,C和Ph秋季最高,D春季最高,而O和Pl秋季最高。
通过对两周年各功能群进行无重复双因素方差分析(表 3),结果表明,第一、第2周年D的密度在生境间差异显著,其余各功能群的密度在生境间均无显著性差异;第2周年Ph和Pl的生物量在生境间差异显著,其余各功能群的生物量在生境间均无显著性差异。第1周年O和Pl、第2周年O和Ph的密度在季节间差异显著,其余各功能群的密度在季节间均无显著性差异;各功能群的生物量在生境间均无显著性差异。
| 周年 Years | 参数 Parameters | 功能群 Functional groups | 生境间Different habitats | 季节间Different seasons | ||
| F2,6 | P | F3,6 | P | |||
| 第1周年1st year | 密度/(个/m2) | C | 0.94 | >0.05 | 0.85 | >0.05 |
| D | 5.29 | <0.05* | 0.14 | >0.05 | ||
| O | 1.95 | >0.05 | 4.94 | <0.05* | ||
| Ph | 1.02 | >0.05 | 1.58 | >0.05 | ||
| Pl | 1.38 | >0.05 | 5.39 | <0.05* | ||
| 生物量/(g/m2) | C | 0.75 | >0.05 | 0.72 | >0.05 | |
| D | 1.01 | >0.05 | 0.35 | >0.05 | ||
| O | 0.87 | >0.05 | 3.73 | >0.05 | ||
| Ph | 1.15 | >0.05 | 1.29 | >0.05 | ||
| Pl | 1.50 | >0.05 | 1.41 | >0.05 | ||
| 第2周年2nd year | 密度/(个/m2) | C | 3.11 | >0.05 | 0.94 | >0.05 |
| D | 10.14 | <0.05* | 1.65 | >0.05 | ||
| O | 4.10 | >0.05 | 7.98 | <0.05* | ||
| Ph | 0.46 | >0.05 | 6.49 | <0.05* | ||
| Pl | 1.22 | >0.05 | 1.66 | >0.05 | ||
| 生物量/(g/m2) | C | 3.66 | >0.05 | 1.43 | >0.05 | |
| D | 0.44 | >0.05 | 0.42 | >0.05 | ||
| O | 1.17 | >0.05 | 1.06 | >0.05 | ||
| Ph | 5.58 | <0.05* | 1.13 | >0.05 | ||
| Pl | 10.38 | <0.05* | 1.77 | >0.05 | ||
| *差异显著(Significant differences),P<0.05 | ||||||
两周年3种生境下功能群多样性指数情况见表 4,其变化的总体趋势是:第1周年3种多样性指数在各生境间波动较大,而第2周年波动相对平缓。第1周年S、H′最大值均出现在HH中,J最大值出现在HS中,而GT中3种多样性指数均较低;第2周年H′、J最大值均出现在HS中,S最大值出现在GT中,而HH中3种多样性指数均最低。
| 周年 Years | 生境 Habitat | C | D | O | Ph | Pl | ||||||||||
| S | H′ | J | S | H′ | J | S | H′ | J | S | H′ | J | S | H′ | J | ||
| 第1周年 1st year | HH | 1.24 | 1.25 | 0.57 | 0.72 | 1.14 | 0.82 | 0.61 | 0.93 | 0.84 | 0.95 | 0.54 | 0.26 | 0.45 | 0.81 | 0.50 |
| GT | 0.95 | 1.39 | 0.72 | 0.40 | 0.64 | 0.92 | 0.87 | 1.05 | 0.96 | 0.73 | 1.40 | 0.78 | 0.30 | 0.46 | 0.33 | |
| HS | 1.09 | 1.53 | 0.79 | 0.38 | 0.68 | 0.99 | 0.97 | 0.35 | 0.26 | 0.67 | 0.91 | 0.51 | 0.40 | 0.45 | 0.28 | |
| 第2周年 2nd year | HH | 1.19 | 1.68 | 0.94 | 0.88 | 0.93 | 0.52 | 0.35 | 0.53 | 0.76 | 1.08 | 1.30 | 0.59 | 0.25 | 0.10 | 0.09 |
| GT | 1.42 | 1.55 | 0.71 | 1.21 | 1.53 | 0.86 | 1.45 | 1.78 | 0.92 | 0.97 | 1.28 | 0.62 | 0.23 | 0.70 | 0.63 | |
| HS | 1.51 | 1.59 | 0.69 | 0.75 | 1.12 | 0.81 | 1.02 | 1.19 | 0.74 | 0.93 | 1.18 | 0.57 | 0.25 | 0.33 | 0.30 | |
就不同年度不同生境5个功能群的多样性指数而言,总体变化趋势是Ca的3种多样性指数均处在较高水平,Pl较低。两周年中,S最大值出现在第2周年HS的Ca中,最小值出现在第2周年GT的Pl中;H′最大值出现在第2周年GT的Om中,最小值出现在第2周年HH的Pl中;J最大值出现在第1周年HS的De中,最小值出现在第2周年HH的Pl中。通过对两周年各功能群物种多样性指数进行无重复双因素方差分析,结果表明,两周年中3种多样性指数在生境间均无显著性差异(P>0.05);两周年中S在功能群间均有极显著差异(P<0.01),第1周年H′、J在功能群间均无显著性差异(P>0.05),第2周年H′有极显著差异(F4,8=7.83,P=0.007),J有显著差异(F4,8=4.37,P=0.036)。
3 讨论 3.1 功能群的组成大型底栖动物功能群是海洋生境和环境质量的综合反映[24-26]。调查区域的生境不同,功能群的类型、优势地位及其物种组成可能也不同。漩门湾两周年调查共获得大型底栖动物49种,其中Pl有5种;Ph有5种;C有14种;O有11种;D有7种。调查期间的3种生境均以Pl和Ph为优势功能类群,其中Pl的代表种为焦河蓝蛤和短拟沼螺,Ph的代表种为珠带拟蟹守螺和泥蟹。与象山港[15]、长江口九段沙[20]、灵昆东滩围垦区[21]和胶州湾西北部潮滩[23]有共同的优势类群,但其优势种不同,如象山港以Pl为优势类群,代表种为缢蛏(Sinonovacula constricta)和菲律宾蛤仔(Ruditapes philippiuarum);灵昆东滩围垦区以Ph为优势类群,优势种为珠带拟蟹守螺和尖锥拟蟹守螺(Cerithidea largillierti)。而彭松耀等[27]的研究表明,C功能群在乳山近海占较大优势,其优势种为寡鳃齿吻沙蚕(Nephtys oligobranchia)和纽虫(Cerebratulina sp.)等;李少文等[28]的研究显示,优势功能群同样为C,占总种数的42.37%,其代表种有寡鳃齿吻沙蚕(Nephtys oligobranchia)和澳洲鳞沙蚕(Aphrodita australis);另外,廖一波等[29]的研究也显示C功能群的种类数比例最高,占总数的33.3%,代表种有长吻沙蚕(Glycera chirori)和双鳃内卷齿蚕(Aglaophamus dibranchisa)。这可能与漩门湾滩涂湿地的软泥底质有关,由于该底质主要由较细颗粒沉积物构成,沉积速率大,致使底质经常处于扰动变化之中,限制了在海水、底质交界处生活的类群(如多毛类、腔肠和棘皮动物等)的生长,那些个体小、以沉积物为食的埋栖性动物(如软体动物和甲壳动物)则易于占据优势地位[30]。
3.2 功能群的密度和生物量大型底栖动物的密度、生物量以及功能群多样性受多种因素的影响,如水温、底质、水化学条件和人为因素等[31],其中生境与大型底栖动物功能群的组成结构存在一定关系[32]。具体而言,植被的类型、覆盖度以及初级生产力的高低,将直接影响营养源的分布和食物结构,从而进一步影响底栖动物功能群的结构[33-34]。围垦外滩涂中植被的不同,使得底栖动物功能群的分布在时空上存在差异。红树林是生产力最高的生态系统类型之一[35],生态意义十分明显。红树林为周围环境输出了大量的有机物,可以逐渐改变生境内土壤的性质,如盐度、酸度以及营养物质含量等,为林下动物提供了索饵场、避难所和栖息地,进而影响底栖动物的组成和分布[36-37]。互花米草具有发达的根系和粗壮的植株,大面积分布时能作为“生物软堤坝”,大大降低了以石块或钢筋混凝土筑坝护岸的成本。此外,互花米草还具有突出的促进淤泥沉降能力,当水流裹挟泥沙进入互花米草时,水流速度受阻,泥沙被互花米草的茎叶阻挡,沉降于互花米草滩中[38]。但互花米草具有较强的入侵能力,能够大大降低大型底栖无脊椎动物的密度和物种丰富度[39],尤其在成熟的互花米草浓密区,其密集的地上枝干和发达的地下根系严重影响了底栖动物的栖息和生长[40],特别是抑制了底内潜穴型动物的生存[41],如一些软体动物门的瓣鳃纲种类和环节动物门种类。这与江苏射阳河口[42]、长江口崇明东滩湿地[43]和广西北海互花米草滩[44]的研究结果相似,可见互花米草入侵光滩后,促淤造陆的作用导致高程增加,所在生境被潮水淹没的次数和时间减少,但相对稳定的环境,为运动迟缓的动物(如软体动物门的腹足纲种类)提供了优良的生存条件,使其大量繁殖,因此互花米草入侵可能增加底埋型动物类群数量。通过对两周年各功能群进行无重复双因素方差分析表明,第一、第2周年D的密度在生境间差异显著,第1周年O和Pl、第2周年的O和Ph密度在季节间差异显著,第2周年Ph和Pl的生物量在生境间差异显著,其余各功能群的密度或生物量在生境间和季节间均无显著性差异。由此可以判断,各功能群的密度和生物量在不同年度之间所占据的比例不同。HH,GT和HS中密度最大值均出现在第1周年春季的Pl功能群,且HS(3540.40 个/m2)>GT(3206.40 个/m2)>HH(609.20 个/m2);HH和HS的生物量最大值均出现在第2周年秋季的Ph功能群,GT为第1周年的Pl功能群,且HS(72.68 g/m2)>HH(70.13 g/m2)>GT(31.46 g/m2)。主要原因是第1周年春季采获大量的焦河蓝蛤幼体,而焦河蓝蛤恰好处在繁殖期(4月份)[45],其栖息密度在HS中最高可达3402.00 个/m2;第2周年焦河蓝蛤栖息密度最大值出现在夏季的GT中,仅有118.40 个/m2,而且焦河蓝蛤在生物量上所占的比重较小,第2周年HS中在生物量所占比重较大的珠带拟蟹守螺增多,生物量在秋季高达67.53 g/m2,这就使得HS中密度和生物量最大值分别出现在第1周年春季和第2周年秋季。但此次采样区域的红树林由2010年刚引种的秋茄红树构成,由于生长年限较短,生境处在发育初期,尚未对其异质性产生大的影响;此外,采样前一年红树林遭受霜冻,生长状况不佳,导致HS和GT在第1周年Pl功能群的密度和生物量相差不大。HH生境中,Pl的密度虽然较低,但生物量较高,和HS相差不大,这是因为珠带拟蟹守螺的栖息密度在HH中较高(113.6 g/m2),而其生物量比重较大。
3.3 功能群的多样性漩门湾围垦多年,长年促淤使得围垦后新形成的潮滩高程逐年升高,各潮位之间的高程差己不明显,盐度、潮汐动力等因素的改变使底栖动物功能群的分布发生明显的变化[46]。尤其围垦后盐沼植被迅速扩张,如人工引种的红树林,使潮水流速减慢,为潮间带的植食者以及滤食生活的浮游生物食者提供了丰富的食物来源和稳定的生存环境,因而这两种功能群的丰度和多样性在围垦潮滩均处于较高水平。此次采样调查的红树林生境由2010年刚引种的秋茄红树组成,而一般认为,红树林植被增加了生境的复杂性,在发育初期有益于增加底栖动物的多样性[47],这与本研究的结果吻合。围垦之后,互花米草开始向低潮带的光滩蔓延,加之人工红树林的大面积种植,植被呈连续的斑块状出现,一些在植被中生活的植食者和浮游生物食者由于种群的扩大而迅速扩张,这必然导致其在红树林和互花米草中的生存空间不足而迁移到光滩。植食者和浮游生物食者的增加,使其余几种功能群比例减少。这种变化最终可能导致潮滩初级生产力不断下降,这与围垦将近10年的长江口横沙东滩围垦潮滩[9]调查结果相似,而初级生产力的不断下降可能不利于功能群的类型趋于多样化[48]。另外,鲍毅新等[21]的研究也表明,互花米草的入侵在一定程度上降低了研究站点的生物多样性。两周年各功能群物种多样性指数无重复双因子方差分析的结果表明,3种多样性指数在生境间均无显著性差异,说明围垦后新形成的3种生境对物种多样性的影响较小。而多样性指数在各功能群之间差异显著,表明各功能群的物种多样性差异显著,在生境中所处的地位不同。连续两周年的调查结果显示,各生境的物种数、密度、生物量、多样性均处在不断的变化之中,表明漩门湾围垦后新形成的滩涂湿地各生境仍处在演替阶段,各种环境指数尚未达到稳定。
| [1] | 彭少麟, 任海, 张倩媚. 退化湿地生态系统恢复的一些理论问题. 应用生态学报 , 2003, 14 (11) : 2026–2030. |
| [2] | 胡知渊, 李欢欢, 鲍毅新, 葛宝明. 灵昆岛围垦区内外滩涂大型底栖动物生物多样性. 生态学报 , 2008, 28 (4) : 1498–1507. |
| [3] | Sato S, Yamashita H, Kim K, Matsuo M. Human impacts on Coastal Communities: A case study of benthic faunal changes after Dike Construction in Saemangeum, South Korea. The Quaternary Research , 2007, 46 : 265–274. DOI:10.4116/jaqua.46.265 |
| [4] | Yang H Y, Chen B, Barter M Piersma T, Zhou C F, Li F S, Zhang Z W. Impacts of tidal land reclamation in Bohai Bay, China: ongoing losses of critical Yellow Sea waterbird staging and wintering sites. Bird Conservation International , 2011, 21 (3) : 241–259. DOI:10.1017/S0959270911000086 |
| [5] | 牛俊英, 衡楠楠, 张斌, 袁晓, 王天厚. 上海市南汇东滩围垦后海岸带湿地冬春季水鸟生境选择. 动物学研究 , 2011, 32 (6) : 624–630. |
| [6] | Covich A P, Palmer M A, Crowl T A. The role of benthic invertebrate species in freshwater ecosystems: zoobenthic species influence energy flows and nutrient cycling. BioScience , 1999, 49 (2) : 119–127. DOI:10.2307/1313537 |
| [7] | 胡知渊, 鲍毅新, 程宏毅, 张龙龙, 葛宝明. 中国自然湿地底栖动物生态学研究进展. 生态学杂志 , 2009, 28 (5) : 959–968. |
| [8] | Harrel R C, Smith S T. Macrobenthic community structure before, during, and after implementation of the Clean Water Act in the Neches River estuary (Texas). Hydrobiologia , 2002, 474 (1/3) : 213–222. DOI:10.1023/A:1016596123558 |
| [9] | 吕巍巍, 马长安, 余骥, 田伟, 袁晓, 赵云龙. 长江口横沙东滩围垦潮滩内外大型底栖动物功能群研究. 生态学报 , 2013, 33 (21) : 6825–6833. |
| [10] | 李妍, 李国勇, 慕军鹏, 孙书存. 消费者多样性对食物网结构和生态系统功能的影响. 生态学报 , 2008, 28 (1) : 388–398. |
| [11] | Sanders J L, Kendall M A, Hawkins A J S, Spicer J I. Can functional groups be used to indicate estuarine ecological status?. Hydrobiologia , 2007, 588 (1) : 45–58. DOI:10.1007/s10750-007-0651-4 |
| [12] | Terbourgh J, Robinson S. Guilds and their utility in ecology// Kikkawa J, Anderson D J, eds. Community ecology: Pattern and Processes. Melbourne: Blackwell Scientific Publications , 1986 : 65–99. |
| [13] | Andersen A N. A classification of Australian ant communities, based on functional groups which parallel plant life-forms in relation to stress and disturbance. Journal of Biogeography , 1995, 22 (1) : 15–29. DOI:10.2307/2846070 |
| [14] | 袁兴中, 陆健健, 刘红. 长江口底栖动物功能群分布格局及其变化. 生态学报 , 2002, 22 (12) : 2054–2062. |
| [15] | 尤仲杰, 陶磊, 焦海峰, 施慧雄, 楼丹. 象山港大型底栖动物功能群研究. 海洋与湖沼 , 2011, 42 (3) : 431–435. |
| [16] | Valença A P M C, Santos P J P. Macrobenthic community for assessment of estuarine health in tropical areas (Northeast, Brazil): Review of macrofauna classification in ecological groups and application of AZTI Marine Biotic Index. Marine Pollution Bulletin , 2012, 64 (9) : 1809–1820. DOI:10.1016/j.marpolbul.2012.06.003 |
| [17] | Conde A, Novais J M, Dominguez J. Multivariate analysis applied to agglomerated macrobenthic data from an unpolluted estuary. Marine Environmental Research , 2013, s87-88 (2) : 112–121. |
| [18] | Gogina M, Zettler M L. Diversity and distribution of benthic macrofauna in the Baltic Sea: Data inventory and its use for species distribution modelling and prediction. Journal of Sea Research , 2010, 64 (3) : 313–321. DOI:10.1016/j.seares.2010.04.005 |
| [19] | Roleta C, Spilmont N, Dewarumez J M, Luczak C. Linking macrobenthic communities structure and zonation patterns on sandy shores: mapping tool toward management and conservation perspectives in northern France. Continental Shelf Research , 2015, 99 : 12–25. DOI:10.1016/j.csr.2015.03.002 |
| [20] | 朱晓君, 陆健健. 长江口九段沙潮间带底栖动物的功能群. 动物学研究 , 2003, 24 (5) : 355–361. |
| [21] | 鲍毅新, 胡知渊, 李欢欢, 葛宝明, 程宏毅. 灵昆东滩围垦区内外大型底栖动物季节变化和功能群的比较. 动物学报 , 2008, 54 (3) : 416–427. |
| [22] | 葛宝明, 鲍毅新, 程宏毅, 李欢欢, 胡知渊. 灵昆岛东滩潮间带大型底栖动物功能群及营养等级构成. 生态学报 , 2008, 28 (10) : 4796–4804. |
| [23] | 辛俊宏, 任一平, 徐宾铎, 张崇良, 薛莹, 纪毓鹏. 胶州湾西北部潮滩湿地大型底栖动物功能群. 应用生态学报 , 2011, 22 (7) : 1885–1892. |
| [24] | Gaudêncio M J, Cabral H N. Trophic structure of macrobenthos in the Tagus estuary and adjacent coastal shelf. Hydrobiologia , 2007, 587 (1) : 241–251. DOI:10.1007/s10750-007-0686-6 |
| [25] | Carvalho S, Pereira P, Pereira F, De Pablo H, Vale C, Gaspar M B. Factors structuring temporal and spatial dynamics of macrobenthic communities in a eutrophic coastal lagoon (Óbidos lagoon, Portugal). Marine Environmental Research , 2011, 71 (2) : 97–110. DOI:10.1016/j.marenvres.2010.11.005 |
| [26] | Dimitriadis C, Evagelopoulos A, Koutsoubas D. Functional diversity and redundancy of soft bottom communities in brackish waters areas: Local vs regional effects. Journal of Experimental Marine Biology and Ecology , 2012, 426-427 : 53–59. DOI:10.1016/j.jembe.2012.05.016 |
| [27] | 彭松耀, 李新正. 乳山近海大型底栖动物功能摄食类群. 生态学报 , 2013, 33 (17) : 5274–5285. |
| [28] | 李少文, 刘元进, 李凡, 张莹, 徐宗法, 吕振波, 王田田, 张爱波. 莱州湾大型底栖动物功能群现状. 生态学杂志 , 2013, 32 (2) : 380–388. |
| [29] | 廖一波, 寿鹿, 曾江宁, 高爱根, 汤雁滨, 严小军, 陈全震. 浙江西门岛海洋特别保护区大型底栖动物功能群特征及其与环境的关系. 生物多样性 , 2013, 21 (1) : 3–10. |
| [30] | Rhoads D C, Young D K. The influence of deposit-feeding organisms on sediment stability and community trophic structure. Journal of Marine Research , 1970, 28 : 150–178. |
| [31] | 于海燕, 李新正, 李宝泉, 王金宝, 王洪法. 胶州湾大型底栖动物生物多样性现状. 生态学报 , 2006, 26 (2) : 416–422. |
| [32] | Naeem S, Li S B. Biodiversity enhances ecosystem reliability. Nature , 1997, 390 (6659) : 507–509. DOI:10.1038/37348 |
| [33] | 袁兴中. 河口潮滩湿地底栖动物群落的生态学研究. 上海: 华东师范大学, 2001. |
| [34] | Spieles D J, Mitsch W J. A model of macroinvertebrate trophic structure and oxygen demand in freshwater wetlands. Ecological Modelling , 2003, 161 (3) : 183–194. DOI:10.1016/S0304-3800(02)00342-3 |
| [35] | Lugo A E, Snedaker S C. The ecology of mangroves. Annual Review of Ecology and Systematics , 1974, 5 : 39–64. DOI:10.1146/annurev.es.05.110174.000351 |
| [36] | 唐富江, 姜作发, 刘曼红, 霍堂斌, 李喆, 张丽荣. 新疆吉力湖底栖动物调查. 水产学杂志 , 2009, 22 (4) : 42–45. |
| [37] | 许贻斌, 李秀珠, 吴立峰, 钱小明, 郑惠东, 蔡建堤. 厦门筼筜湖大型底栖生物调查及其评价. 福建水产 , 2009 (2) : 18–23. |
| [38] | 李加林, 许继琴, 张殿发, 杨晓平, 童亿勤, 沈永明. 杭州湾南岸互花米草盐沼生态系统服务价值评估. 地域研究与开发 , 2005, 24 (5) : 58–62. |
| [39] | Neira C, Levin L A, Grosholz E D. Benthic macrofaunal communities of three sites in San Francisco Bay invaded by hybrid Spartina with comparison to uninvaded habitats. Marine Ecology Progress Series , 2005, 292 (1) : 111–126. |
| [40] | 赵永强, 曾江宁, 陈全震, 高爱根, 寿鹿, 廖一波, 徐晓群, 刘晶晶. 不同互花米草(Spartina alterniflora Loise)密度生境中大型底栖动物群落格局. 自然资源学报 , 2009, 24 (4) : 630–639. |
| [41] | 杨明生, 熊邦喜, 杨学芬. 武汉市南湖大型底栖动物的时空分布和氮磷评价. 湖泊科学 , 2007, 19 (6) : 658–663. |
| [42] | 仇乐, 刘金娥, 陈建琴, 王国祥, 常青. 互花米草扩张对江苏海滨湿地大型底栖动物的影响. 海洋科学 , 2010, 34 (8) : 50–55. |
| [43] | 徐晓军, 王华, 由文辉, 刘宝兴. 崇明东滩互花米草群落中底栖动物群落动态的初步研究. 海洋湖沼通报 , 2006 (2) : 89–95. |
| [44] | 赵彩云, 白加德, 柳晓燕, 李俊生. 互花米草入侵对广西北海光滩大型底栖动物群落的影响. 环境科学研究 , 2015, 28 (3) : 377–383. |
| [45] | 张新峰, 李金明, 王立群, 曾现英. 焦河蓝蛤生长、繁殖与环境的关系. 渔业现代化 , 2004 (5) : 16–17. |
| [46] | Windham L, Lathrop R G. Effects of Phragmites australis(common reed) invasion on aboveground biomass and soil properties in brackish tidal marsh of the Mullica River, New Jersey. Estuaries , 1999, 22 (4) : 927–935. DOI:10.2307/1353072 |
| [47] | 徐小雨, 周立志, 朱文中, 许仁鑫, 曹玲亮, 陈锦云, 王勋. 安徽菜子湖大型底栖动物的群落结构特征. 生态学报 , 2011, 31 (4) : 943–953. |
| [48] | 李欢欢, 鲍毅新, 胡知渊, 葛宝明. 杭州湾南岸大桥建设区域潮间带大型底栖动物功能群及营养等级的季节动态. 动物学报 , 2007, 53 (6) : 1011–1023. |