 2016, Vol. 36
2016, Vol. 36文章信息
- 金宇, 周可新, 高吉喜, 穆少杰, 张小华
- JIN Yu, ZHOU Kexin, GAO Jixi, MU Shaojie, ZHANG Xiaohua.
- 基于随机森林模型的国家重点保护陆生脊椎动物物种优先保护区的识别
- Identifying the priority conservation areas for key national protected terrestrial vertebrate species based on a random forest model in China
- 生态学报[J]. 2016, 36(23): 7702-7712
- Acta Ecologica Sinica[J]. 2016, 36(23): 7702-7712
- http://dx.doi.org/10.5846/stxb201512082454
-
文章历史
- 收稿日期: 2015-12-08
- 修订日期: 2016-04-25





2. 环境保护部南京环境科学研究所, 南京 210042
2. Nanjing Institute of Environmental Sciences, Ministry of Environmental Protection, Nanjing 210042, China
近年来, 随着全球气候变化的加剧和社会经济的发展, 生物多样性也受到强烈的影响:一方面, 气候变化(全球温度升高、降水模式改变)使物种的地理分布(适生区域)发生了迁移(改变)[1-2], 另一方面, 人类活动导致栖息地破碎化程度加剧, 而采集和捕猎等活动也使野生动植物种群数量锐减。近年来的研究表明, 全球生物多样性正以前所未有的速度丧失[3]。世界自然保护联盟(International Union for Conservation of Nature, IUCN)发布的《受威胁物种红色名录》表明, 全球约有1/4哺乳动物, 1200多种鸟类以及3万多种植物面临灭绝危险[4]。如何有效保护生物多样性已成为全世界共同面临的几大环境问题之一。
中国生物多样性丰富, 是全球12个“巨大多样性国家”之一[5], 在全球的生物多样性保护事业中发挥至关重要的作用。目前, 中国政府已经采取一系列有效措施应对这一挑战, 如成立了中国生物多样性保护国家委员会, 制定了《中国生物多样性保护战略与行动计划》等[6]。2013年, 中国提出了生态红线战略, 使生物多样性保护工作迈上了一个新台阶。生态红线是指对维护国家和区域生态安全及社会经济可持续发展具有重要战略意义的关键生态保护区域[7]。生态红线的划定能够维持关键物种、生态系统与种质资源生存的最小面积, 有效地保护生物多样性。中国国家重点保护陆生脊椎动物物种的优先保护区是生态红线的重要组成部分, 包含在生态红线中。如何准确地识别和确定优先保护区是生物多样性保护研究中的热点问题之一[8], 也是进行保护活动的前提条件。准确的识别和确定优先保护区能够指导资源的合理分配, 使保护效益达到最大化[9-10]。目前, 世界上许多国家和地区已经开展了生物多样性优先保护区的识别和确定研究, 并在保护中取得了很好的效果[11-14]。
准确有效地获取物种的适宜分布区对于识别和确定优先保护区至关重要[15-16], 物种分布预测技术和地理信息系统(GIS)为生物多样性优先保护区的识别提供了一个可靠的方法[17]。该方法基于物种分布相关数据和环境因素, 利用物种分布预测模型来获取物种的适宜分布区域, 这在全球范围内得到了广泛的应用[18-19]。Austin和Meyers[20]通过广义线性模型和广义加法模型对森林树种的适宜分布区域进行预测, 提出了森林生物多样性的保护与管理措施。Godoy-Bürki等[21]利用MAXENT模型对安第斯山脉的505种原生植被物种的适宜分布区进行预测, 并划分不同的保护优先级, 得到了安第斯山脉原生植被的优先保护区, 并指出优先保护区与生物多样性热点地区重合度不高。Stockman等[22]利用GARP模型对42种无脊椎动物的适宜分布区进行了预测。国内学者在这方面的研究也很多。毕迎凤[23]运用组合模型对铁杉在中国的分布进行了预测, 得到了铁杉的适宜分布区, 并对不同模型的预测结果进行比较。LI等[24]利用组合模型预测了未来气候变化对我国208种特有或濒危动物物种适宜分布区的影响, 发现其中一半动物物种的潜在生境面积将逐渐下降, 另外一半物种的潜在生境面积将有所增加, 135个物种当前适宜分布区的留存率将低于50%。目前已使用的有很多物种分布模型, 包括回归模型、生态位模型和机器学习模型等。不同研究者对现有的模型进行了比较, 均认为随机森林(random forests)模型整体表现最佳[25-27]。
本研究选择了国家一、二级保护动物物种中的263种国家重点保护陆生脊椎动物物种作为研究对象, 采用随机森林模型对每个物种进行建模, 预测各个物种在背景点的适生概率。在此基础上, 得到国家重点保护陆生脊椎动物物种的生境适宜性指数, 并对其进行空间自相关分析, 实现国家重点保护陆生脊椎动物物种优先保护区的识别和确定, 最后对优先保护区进行保护空缺分析。准确可靠地识别和确定国家重点保护陆生脊椎动物物种的优先保护区, 有助于明确最急需保护的区域, 为政府部门的决策提供科学依据, 使有限的资源得到合理分配和充分利用。
1 材料和方法 1.1 物种的分布数据动物物种的分布数据来自中国科学院动物研究所开发的中国物种信息系统(CSIS)(http://monkey.ioz.ac.cn/bwg-cciced/chinese/cesis/csispage.htm)[28], 该系统中记录了一万多种动物物种, 包括物种的历史分布点、受威胁状况、在我国的分布状况及保护等级等信息。本研究选择物种的标准是重要性、特有性以及数据的完整性, 选择了其中的国家一、二级保护动物362种, 剔除鱼类等水生、半水生物种以及分布点数据不足的物种共99种(表 1), 保留了263种动物物种作为研究对象, 并根据中国物种红色名录[29]对物种的分布点数据进行了检查, 以保证数据的准确性。其中, 86种物种为国家一级保护动物, 177种为国家二级保护动物, 26种为中国特有物种。本研究选取的263种物种中, 被IUCN《受威胁物种红色名录》评定为极危(CR)物种的有18种, 濒危(EN)物种58种, 易危(VU)物种50种, 受到不同程度威胁的物种占所选物种的47.9%。
| 序号 Serial number | 物种名 Species | 保护级别 Conservation grade | 序号 Serial number | 物种名 Species | 保护级别 Conservation grade |
| 1 | 白腹军舰鸟Fregata andrewsi | 一级 | 2 | 东方白鹳Ciconia ciconia | 一级 |
| 3 | 黑头角雉Tragopan melanocephalus | 一级 | 4 | 灰腹角雉Tragopan blythii | 一级 |
| 5 | 野马Equus caballus | 一级 | 6 | 豚鹿Axis porcinus | 一级 |
| 7 | 白掌长臂猿Hylobates lar | 一级 | 8 | 戴帽叶猴Trachypithecus shortridgei | 一级 |
| 9 | 麋鹿Elaphurus davidianus | 一级 | 10 | 坡鹿Ruservus eldii | 一级 |
| 11 | 黔金丝猴Rhinopithecus brelichi | 一级 | 12 | 四爪陆龟Testudo horsfieldii | 一级 |
| 13 | 扬子鳄Alligator sinensis | 一级 | 14 | 白背兀鹫Gyps bengalensis | 二级 |
| 15 | 白眼鵟鹰Butastur teesa | 二级 | 16 | 斑尾林鸽Columba palumbus | 二级 |
| 17 | 彩鹮Plegadis falcinellus | 二级 | 18 | 仓鸮Tyto alba | 二级 |
| 19 | 橙胸咬鹃Harpactes oreskios | 二级 | 20 | 短尾鹦鹉Loriculus vernalis | 二级 |
| 21 | 鹳嘴翡翠Halcyon capensis | 二级 | 22 | 海鸬鹚Phalacrocorax pelagicus | 二级 |
| 23 | 黑鹮Pseudibis davisoni | 二级 | 24 | 黑颏果鸠Ptilinopus leclancheri | 二级 |
| 25 | 黑嘴端凤头燕鸥Sterna bernsteini | 二级 | 26 | 红顶绿鸠Treron formosae | 二级 |
| 27 | 红脚鲣鸟Sula sula | 二级 | 28 | 红领绿鹦鹉Psittacula krameri | 二级 |
| 29 | 红腿小隼Microhierax caerulescens | 二级 | 30 | 红胸黑雁Branta ruficollis | 二级 |
| 31 | 黄脚绿鸠Treron phoenicoptera | 二级 | 32 | 灰头绿鸠Treron pompadora | 二级 |
| 33 | 兰屿角鸮Otus elegans | 二级 | 34 | 蓝八色鸫Pitta cyanea | 二级 |
| 35 | 蓝耳翠鸟Alcedo meninting | 二级 | 36 | 栗鸮Phodilus badius | 二级 |
| 37 | 镰翅鸡Dendragapus falcipennis | 二级 | 38 | 绿胸八色鸫Pitta sordida | 二级 |
| 39 | 毛腿渔鸮Ketupa blakistoni | 二级 | 40 | 双辫八色鸫Pitta phayrei | 二级 |
| 41 | 铜翅水雉Metopidius indicus | 二级 | 42 | 乌雕鸮Bubo coromandus | 二级 |
| 43 | 乌灰鹞Circus pygargus | 二级 | 44 | 细痣疣螈Tylototriton asperrimus | 二级 |
| 45 | 靴隼雕Hieraaetus pennata | 二级 | 46 | 渔鵰Ichthyophaga humilis | 二级 |
| 47 | 针尾绿鸠Treron apicauda | 二级 | 48 | 镇海棘螈Echinotriton chinhaiensis | 二级 |
| 49 | 棕翅鵟鹰Butastur liventer | 二级 | 50 | 棕腹隼雕Hieraaetus kienerii | 二级 |
| 51 | 棕头鹃鸠Macropygia ruficeps | 二级 | 52 | 驯鹿Rangifer tarandus | 二级 |
| 53 | 印度穿山甲Manis crassicaudata | 二级 | 54 | 凹甲陆龟Manouria impressa | 二级 |
环境变量是随机森林模型进行建模和预测的依据。本研究选择了12个能够反映气候特征、栖息地和人类影响的环境变量(表 2)来预测物种的适宜分布区[18, 24, 30-31]。其中年平均温度、最冷月份最低温度、最暖月份最高温度、温度的季节性变异、年降水量和降水的季节性变异为生物气候变量; 土地利用类型、生物群落区和湿地类型为地表类型变量; 海拔为地形变量; 人类足迹指数和净初级生产力为人文、生态指数变量。所有的变量中除了3个地表类型变量为分类型变量, 其它9个变量均为连续型变量。
| 环境变量Environment variables | 缩写Abbreviations | 来源Data sources |
| 年平均温度Annual mean temperature | T.am | WORLDCLIM数据库(http://www.worldclim.org) |
| 最冷月份最低温度Min temperature of coldest month | T.min | WORLDCLIM数据库(http://www.worldclim.org) |
| 最暖月份最高温度Max temperature of warmest month | T.max | WORLDCLIM数据库(http://www.worldclim.org) |
| 温度的季节性变异Temperature seasonality | T.sd | WORLDCLIM数据库(http://www.worldclim.org) |
| 年降水量Annual precipitation | P.annual | WORLDCLIM数据库(http://www.worldclim.org) |
| 降水的季节性变异Precipitation seasonality | P.cv | WORLDCLIM数据库(http://www.worldclim.org) |
| 生物群落区Biomes | Biome | 世界野生动物基金会(http://worldwildlife.org) |
| 土地利用类型Land use types | Landuse | 美国地质调查局(http://edc2.usgs.gov) |
| 湿地类型Wetland types | Wetland | 中国科学院遥感应用研究所 |
| 人类足迹指数Human footprint index | Footprint | 美国宇航局社会经济数据和应用中心(https://www.ciesin.columbia.edu/wild_areas/) |
| 净初级生产力Net primary productivity | NPP | 中国科学院遥感应用研究所 |
| 海拔Elevation | Elev | 美国地质调查局(http://eros.usgs.gov) |
随机森林模型是由Breiman在2001年开发出来的一种汇总式自学习数据处理方法, 是一种现代分类与回归技术, 同时, 它也是一种组合式的自学习技术[32]。随机森林模型是基于分类树算法进行模拟和迭代, 在变量和数据的使用上进行随机化, 生成若干分类树, 再汇总分类树的结果[33], 从而进行判别。随机森林模型对多元共线性不敏感; 对于大量数据的处理效率较高, 可以预测多达几千个解释变量的作用; 在数据缺失时(如变量个数远大于样本量)仍然有很高的精度。该模型可以通过多种工具实现, 如FORTRUN、SAS (Statistics Analysis System)、R语言等, 本文采用R语言作为模型运行的平台。
将中国大陆地区作为研究区域, 采用公里网格选取预测背景点, 每隔0.2经度和纬度选择一个点, 共计11968个背景点。通过地理信息系统(ArcGIS)软件提取每个物种的分布点和背景点的所有环境变量值。将每个物种的分布数据和背景数据分为训练集和测试集, 其中训练集分别由随机抽取的各75%的物种分布数据和背景数据组成, 测试集是由剩余的各25%的物种分布数据和背景数据组成。训练集用于建模, 测试集用作检验模型精度。建模后, 对每个物种进行预测, 得到背景点的物种的适生概率, 将每个背景点上所有物种的适生概率进行迭加并利用公式(1)进行标准化, 得到国家重点保护陆生脊椎动物物种生境适宜性指数。
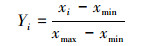
|
(1) |
式中, Yij为第i个点标准化后的生境适宜性指数值, Xi为第i个点的生境适宜性指数值, Xmin为第生境适宜性指最小值, Xmax为生境适宜性指最大值。
模型的预测精度检验是基于模型测试集数据的预测结果来评价模型的预测能力, 采用接收工作机特征曲线下的面积(AUC)和真实技巧统计值(TSS)来评价模型的预测精度。AUC的值为0.90-1.00, 说明模型表现很好; 0.80-0.90, 模型表现好; 0.70-0.80, 模型表现一般; 0.60-0.70, 模型表现差; 0.50-0.60, 模型预测失败[34]。TSS是灵敏度和特异度的函数[35], 公式简单, 易于理解。TSS值的范围是-1到1, 值越大, 说明预测的精度越高。TSS的值为0.85-1, 说明模型表现很好; 0.7-0.85, 模型表现好; 0.55-0.7, 模型表现一般; 0.4-0.55, 模型表现差; < 0.4, 说明预测失败[36]。
1.4 优先保护区的识别及空缺分析在ArcGIS 9.3中将生境适宜性指数进行反距离加权插值运算, 得到1km分辨率的生境适宜性指数图。将生境适宜性指数图中的每个像元转换成1km×1km的矢量网格, 一个正方形就是一个评价单元。我们采用GeoDa软件中空间自相关分析的方法探测生境适宜性指数的空间聚集状况, 识别生境适宜性指数的高值聚集区, 每个评价单元的空间邻接关系创建空间权重矩阵, 邻接方式为共边或共点邻接。进而得到国家重点保护陆生动物物种的优先保护区, 以保证大部分物种的长期存活。同时, 在ArcGIS软件中分别将优先保护区与自然保护区及生物多样性保护优先区进行叠加, 对优先保护区的被保护情况进行分析。自然保护区数据截止到2013年底, 包括国家级、省级和市县级自然保护区。生物多样性保护优先区数据来自《中国生物多样性保护战略与行动计划》中的内陆陆地和水域生物多样性保护优先区域[37]。为了降低空间尺度对空间自相关分析的影响[38], 我们按照陆生动物的地理分区[39], 将生境适宜性指数图划分为东北区、华北区、蒙新区、西南区、华中区、青藏区以及华南区, 共7个陆生动物地理分布区。
空间自相关性使用全局和局部两种指标来度量, 首先使用全局指标探测7个地理分布区生境适宜性指数的空间模式, 确定整个区域是否具有空间自相关性以及相关性的显著水平。再对7个地理分布区进行局部空间自相关性分析, 进一步考虑是否存在观测值的高值或低值的局部空间聚集, 并识别高值区与低值区。表示空间自相关的指标和方法很多, 其中较为常用的是Moran′s I指数[40-41]。Moran′s I指数是用于衡量空间要素间的相互关系, 值的范围在-1到1之间。1表示有强烈的空间正相关, -1则表示有强烈的空间负相关, 0表示不存在空间相关性[42-43]。
全局Moran′s I的计算公式为:
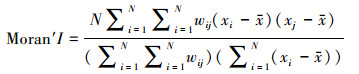
|
(2) |
局部Moran′s I的计算公式为:

|
(3) |
式中, xi和xj分别是i和j所在位置的观测值, wij为权重, δ为xi标准差。
通过SPSS系统聚类方法对未被保护的优先保护区进行分析, 选择面积、国家重点保护陆生脊椎动物物种数、生境适宜性指数、植被类型数以及人类干扰指数这5个指标, 对未被保护的优先保护区的保护顺序进行划分, 将未被保护的优先保护区划分为3类, 即最优先保护、优先保护和中等优先保护[44]。
2 结果 2.1 模型精度检验计算AUC和TSS这2个模型精度检验指标的值, AUC的最小值为0.813, 最大值为0.981, 平均值为0.896;TSS的最小值0.735, 最大值为0.929, 平均值为0.807。说明模型的预测能力较好, 预测精度较高。
2.2 优先保护区的识别通过全局Moran′s I的计算公式(式2)计算得到7个地理分布区生境适宜性指数的全局Moran′s I指数(表 3)。结果表明, 各地理分布区生境适宜性指数的全局空间自相关系数均大于0.90, (P < 0.05)。说明7个地理分布区中评价单元的生境适宜性指数存在显著的空间自相关, 可以对其进行空间自相关聚类分析。
| 陆生动物地理分区 Terrestrial zoogeographical regions | 全局Moran′s I指数 Global Moran′s I |
| 东北区Northeast region | 0.914490 |
| 华北区North China region | 0.913328 |
| 华中区Central China region | 0.908987 |
| 青藏区Qinghai Tibet region | 0.936081 |
| 蒙新区Inner Mongolia-Xinjiang region | 0.907958 |
| 华南区South China region | 0.972359 |
| 西南区Southwest region | 0.947173 |
通过局部Moran′s I的计算公式(式3)探测7个地理分布区生境适宜性指数的空间聚集情况并通过ARCGIS软件绘制高值聚集区, 得到了国家重点保护陆生脊椎动物物种的优先保护区(图 1)。优先保护区是生境适宜性指数高值聚集的区域, 生境适宜性指数高说明该区域的国家重点保护动物物种数量多、分布相对集中、人类干扰弱, 是国家重点保护陆生脊椎动物物种分布较多的地区。国家重点保护陆生脊椎动物物种的优先保护区主要分布在我国的西部地区, 包括西南地区的秦岭-大巴山山区、云南省与印度及缅甸的交界地区、武陵山山区、喜马拉雅山-横断山脉山区、阿尔泰山脉山区、天山山脉山区、昆仑山山脉山区; 东北的大、小兴安岭、东北-华南的沿海地区及长江中下游地区有少量分布。通过计算, 优先保护区的面积为103.16万km2, 约占我国中国国土面积的10.90%。优先保护区的斑块个数为355个, 其中面积最大的为9.67万km2, 面积最小的为1.37km2, 平均斑块面积为2153.75km2。

|
| 图 1 国家重点保护陆生脊椎动物物种的优先保护区 Fig. 1 Priority conservation areas of the key national protected terrestrial vertebrate species |
将优先保护区与我国中国现有的自然保护区及中国陆地生物多样性保护优先区的面状数据进行叠加, 得到了优先保护区目前的被保护情况。如图 2所示, 优先保护区被自然保护区覆盖的面积为18.51万km2, 占保护区总面积的14.54%;优先保护区在我国陆地生物多样性保护优先区中的面积为44.54万km2, 占生物多样性保护优先区总面积的19.17%。综合来看, 优先保护区中被保护的面积为50.40万km2, 占优先保护区总面积的48.86%, 说明国家重点保护陆生脊椎动物物种的优先保护区受自然保护区和生物多样性保护优先区的被保护率不高, 未被充分保护。

|
| 图 2 优先保护区的被保护状况 Fig. 2 The protected status of priority conservation areas |
对未被保护的优先保护区进行保护优先顺序划分, 结果如图 3所示。最应优先保护的是国家重点保护陆生脊椎动物物种较丰富、生境适宜性较高、植被类型复杂、面积较小、人类活动干扰强度大的斑块, 主要分布在我国中东部以及西南地区。最优先保护区域大多分布在自然保护区或生物多样性保护优先区周围, 与自然保护区或生物多样性保护优先区的自然条件相似。同时, 我国中东部地区的自然保护区本身面积较小、分布分散、覆盖率较低, 具有大量的保护空缺[45]。建议将未被保护的优先保护区, 特别是其中最优先保护的区域, 作为生物多样性保护的重点地区, 通过优化升级自然保护区加以保护。

|
| 图 3 未被保护的优先保护区保护的优先顺序 Fig. 3 The protected order of the unprotected priority conservation areas |
国家重点保护陆生脊椎动物物种的优先保护区主要分布在中国西部山脉的山区, 分布较为集中, 斑块面积较大。这可能是由于山区地形复杂, 海拔的差异形成了不同的气候梯度, 造成生态位的重叠, 有利于不同物种生存。此外, 山区人类干扰的强度相对较低, 导致这些地区的生物多样性极其丰富, 拥有大量的特有动植物物种。目前, 全球34个生物多样性热点地区中有4个在中国有分布, 分别是喜马拉雅山脉、印缅地区、中亚山区和中国西南山区[46], 本文的研究结果表明国家重点保护陆生动物物种的优先保护区与4片生物多样性保护热点地区在中国境内的部分均有重叠, 说明优先保护区与生物多样性热点地区的一致性较高, 具有很高的保护价值和保护潜力。东北的大、小兴安岭、东北-华南的沿海及长江中下游地区分布较少, 分布较为分散且斑块面积小, 破碎化程度高, 这可能是由于这些地区多海拔变化小, 无法形成多样的地形及气候条件; 同时, 又处在经济发达的地区, 人类干扰强度大。但这些地区的优先保护区大多处在鸟类的中部和东部迁徙路线上[47], 鸟类迁徙过程中的停歇地是迁徙鸟类在繁殖地和非繁殖地之间的联系枢纽, 对于迁徙鸟类完成其完整的生活史过程具有重要作用[48]。针对这些地区的优先保护区开展保护工作, 对我国的生物多样性保护意义重大。
通过将国家重点保护陆生脊椎动物物种优先保护区与主体功能区划中的重点生态功能区及全国生态功能区划进行比较分析(图 4), 发现陆生脊椎动物物种优先保护区在重点生态功能区和全国生态功能区划的很多地区有一定重叠, 特别是在生物多样性较高的秦岭-大巴山山区、武陵山山区、喜马拉雅山-横断山脉山区、阿尔泰山脉山区、天山山脉山区及东北的大、小兴安岭地区。说明这些地区在生态功能发挥和生物多样性维护方面均具有较高的保护价值。然而, 由于划定目的和划定方法的不同, 优先保护区与重点生态功能区及全国生态功能区划虽有重叠, 但重叠的范围不大。以南岭山脉为例, 南岭山脉不仅属于水源涵养功能区, 还是生物多样性保护生态功能区, 在重点生态功能区和全国生态功能区划中划定的面积分别为1.23万km2和7.59万km2, 优先保护区在重点生态功能区及全国生态功能区划中的分布面积分别为0.13万km2和0.54万km2。在对与主体功能区划中的重点生态功能区及全国生态功能区划重叠, 但未加保护的斑块进行保护时, 需要综合考虑多个不同方面及层次的保护需求, 兼顾不同生态功能、生物多样性保护生态功能中的不同类别及国家重点保护陆生脊椎动物物种的保护。

|
| 图 4 优先保护区在重点生态功能区及全国生态功能区划中的分布情况 Fig. 4 Distribution of priority conservation areas in key ecological function zoning and the national ecological function zoning |
当然, 中国关键陆生脊椎动物物种的优先保护区也不是一成不变的, 本文选择了263种动物物种, 其中被优先保护区保护的物种数为256种, 保护率达97.34%。对于未被优先保护区保护的物种以及由于缺少数据未被纳入到分析中的物种, 可以结合这些物种的分布情况, 对优先保护区作适当的补充和调整。同时, 影响动物物种分布的环境因素(如气候、人类干扰、土地利用、植被演替以及潜在的灾难性事件等)、社会经济发展水平以及国家政策的改变, 可能引起动物的适生区域发生迁移和变化, 需要对物种的分布情况进行调查更新, 优先保护区的地理位置和范围也应该进行相应的调整和变更, 从而实现动态、长效的保护与管理。
本文采用随机森林模型预测了263种国家重点保护陆生脊椎动物物种的适生概率, 通过迭加及标准化方法得到生境适宜性指数, 采用空间自相关方法确定了国家重点保护陆生脊椎动物物种的优先保护区。我们认为国家重点保护陆生脊椎动物物种的优先保护区对于我国的生物多样性保护具有重要意义, 优先保护区的确定明确了保护行动中急需关注和保护的区域, 为相关政府部门更有效地分配和利用有限的保护资源提供科学依据。
| [1] | Wuethrich B. How climate change alters rhythms of the wild. Science , 2000, 287 (5454) : 793–795. DOI:10.1126/science.287.5454.793 |
| [2] | Hughes L. Biological consequences of global warming:is the signal already apparent?. Trends in Ecology & Evolution , 2000, 15 (2) : 56–61. |
| [3] | McCarthy D P, Donald P F, Scharlemann J P W, Buchanan G M, Balmford A, Green J M H, Bennun L A, Burgess N D, Fishpoo L D C, Garnett S T, Leonard D L, Maloney R F, Morling P, Schaefer H M, Symes A, Wiedenfeld D A, Butchart S H M. Financial costs of meeting global biodiversity conservation targets:current spending and unmet needs. Science , 2012, 338 (6109) : 946–949. DOI:10.1126/science.1229803 |
| [4] | 薛达元. 《名古屋议定书》的主要内容及其潜在影响. 生物多样性 , 2011, 19 (1) : 113–119. |
| [5] | Hilton-Taylor C. 2000 IUCN Red List of Threatened Species. IUCN, 2000. |
| [6] | 薛达元. 《中国生物多样性保护战略与行动计划》的核心内容与实施战略. 生物多样性 , 2011, 19 (4) : 387–388. |
| [7] | Lü Y H, Ma Z M, Zhang L W, Fu B J, Gao G Y. Redlines for the greening of China. Environmental Science & Policy , 2013, 33 : 346–353. |
| [8] | Kati V, Devillers P, Dufrêne M, Legakis A, Vokou D, Lebrun P. Hotspots, complementarity or representativeness? Designing optimal small-scale reserves for biodiversity conservation. Biological Conservation , 2004, 120 (4) : 471–480. DOI:10.1016/j.biocon.2004.03.020 |
| [9] | Brooks T M, Mittermeier R A, da Fonseca G A B, Gerlach J, Hoffmann M, Lamoreux J F, Mittermeier C G, Pilgrim J D, Rodrigues A S L. Global biodiversity conservation priorities. Science , 2006, 313 (5783) : 58–61. DOI:10.1126/science.1127609 |
| [10] | Linke S, Kennard M J, Hermoso V, Olden J D, Stein J, Pusey B J. Merging connectivity rules and large-scale condition assessment improves conservation adequacy in river systems. Journal of Applied Ecology , 2012, 49 (5) : 1036–1045. DOI:10.1111/jpe.2012.49.issue-5 |
| [11] | Naoe S, Katayama N, Amano T, Akasaka M, Yamakita T, Ueta M, Matsuba M, Miyashita T. Identifying priority areas for national-level conservation to achieve Aichi Target 11:a case study of using terrestrial birds breeding in Japan. Journal for Nature Conservation , 2015, 24 : 101–108. DOI:10.1016/j.jnc.2014.10.002 |
| [12] | Huang J H, Lu X H, Huang J H, Ma K P. Conservation priority of endemic Chinese flora at family and genus levels. Biodiversity and Conservation , 2016, 25 (1) : 23–35. DOI:10.1007/s10531-015-1027-0 |
| [13] | Mittermeier R A, van Dijk P P, Rhodin A G J, Nash S D. Turtle hotspots:an analysis of the occurrence of tortoises and freshwater turtles in biodiversity hotspots, high-biodiversity wilderness areas, and turtle priority areas. Chelonian Conservation and Biology , 2015, 14 (1) : 2–10. DOI:10.2744/ccab-14-01-2-10.1 |
| [14] | Brooks T M, Cuttelod A, Faith D P, Garcia-Moreno J, Langhammer P, Pérez-Espona S. Why and how might genetic and phylogenetic diversity be reflected in the identification of key biodiversity areas?. Philosophical Transactions of the Royal Society of London B:Biological Sciences , 2015, 370 (1662) : 20140019. DOI:10.1098/rstb.2014.0019 |
| [15] | Gray D, Scarsbrook M R, Harding J S. Spatial biodiversity patterns in a large New Zealand braided river. New Zealand Journal of Marine and Freshwater Research , 2006, 40 (4) : 631–642. DOI:10.1080/00288330.2006.9517451 |
| [16] | Lira-Noriega A, Soberón J, Navarro-Sigüenza A G, Nakazawa Y, Peterson A T. Scale dependency of diversity components estimated from primary biodiversity data and distribution maps. Diversity and Distributions , 2007, 13 (2) : 185–195. DOI:10.1111/j.1472-4642.2006.00304.x |
| [17] | Rodríguez J P, Brotons L, Bustamante J, Seoane J. The application of predictive modelling of species distribution to biodiversity conservation. Diversity and Distributions , 2007, 13 (3) : 243–251. DOI:10.1111/ddi.2007.13.issue-3 |
| [18] | Ko C Y, Lin R S, Ding T S, Hsieh C H, Lee P F. Identifying biodiversity hotspots by predictive models:a case study using Taiwan's endemic bird species. Zoological Studies , 2009, 48 (3) : 418–431. |
| [19] | Wan J Z, Wang C J, Yu J H, Han S J, Wang L H, Wang Q G. The ability of Nature Reserves to conserve medicinal plant resources:a case study in northeast China. Ecological Informatics , 2014, 24 : 27–34. DOI:10.1016/j.ecoinf.2014.06.005 |
| [20] | Austin M P, Meyers J A. Current approaches to modelling the environmental niche of eucalypts:implication for management of forest biodiversity. Forest Ecology and Management , 1996, 85 (1/3) : 95–106. |
| [21] | Godoy-Bürki A C, Ortega-Baes P, Sajama J M, Aagesen L. Conservation priorities in the Southern Central Andes:mismatch between endemism and diversity hotspots in the regional flora. Biodiversity and Conservation , 2014, 23 (1) : 81–107. DOI:10.1007/s10531-013-0586-1 |
| [22] | Stockman A K, Beamer D A, Bond J E. An evaluation of a GARP model as an approach to predicting the spatial distribution of non-vagile invertebrate species. Diversity and Distributions , 2006, 12 (1) : 81–89. DOI:10.1111/ddi.2006.12.issue-1 |
| [23] | 毕迎凤, 许建初, 李巧宏, GuisanA, ThuillerW, ZimmermannN E, 杨永平, 杨雪飞. 应用BioMod集成多种模型研究物种的空间分布--以铁杉在中国的潜在分布为例. 植物分类与资源学报 , 2013, 35 (5) : 647–655. |
| [24] | Li X H, Tian H D, Wang Y, Li R Q, Song Z M, Zhang F C, Xu M, Li D M. Vulnerability of 208 endemic or endangered species in China to the effects of climate change. Regional Environmental Change , 2013, 13 (4) : 843–852. DOI:10.1007/s10113-012-0344-z |
| [25] | Thuiller W, Lafourcade B, Engler R, Araújo M B. BIOMOD-a platform for ensemble forecasting of species distributions. Ecography , 2009, 32 (3) : 369–373. DOI:10.1111/eco.2009.32.issue-3 |
| [26] | 张雷, 刘世荣, 孙鹏森, 王同立. 气候变化对马尾松潜在分布影响预估的多模型比较. 植物生态学报 , 2011, 35 (11) : 1091–1105. |
| [27] | 翟天庆, 李欣海. 用组合模型综合比较的方法分析气候变化对朱鹮潜在生境的影响. 生态学报 , 2012, 32 (8) : 2361–2370. |
| [28] | 解焱, 李典谟, MacKinnonJ. 中国生物地理区划研究. 生态学报 , 2002, 22 (10) : 1599–1615. |
| [29] | 汪松, 解焱. 中国物种红色名录. 北京: 高等教育出版社, 2004. |
| [30] | Olson D M, Dinerstein E, Wikramanayake E D, Burgess N D, Powell G V N, Underwood E C, D'amico J A, Itoua I, Strand H E, Morrison J C, Loucks C J, Allnutt T F, Ricketts T H, Kura Y, Lamoreux J F, Wettengel W W, Hedao P, Kassem K R. Terrestrial ecoregions of the world:a new map of life on earth:a new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. BioScience , 2001, 51 (11) : 933–938. DOI:10.1641/0006-3568(2001)051[0933:TEOTWA]2.0.CO;2 |
| [31] | Sanderson E W, Jaiteh M, Levy M A, Redford K H, Wannebo A V, Woolme G. The human footprint and the last of the wild. Bioscience , 2002, 52 (10) : 891–904. DOI:10.1641/0006-3568(2002)052[0891:THFATL]2.0.CO;2 |
| [32] | Breiman L. Random forests. Machine Learning , 2001, 45 (1) : 5–32. DOI:10.1023/A:1010933404324 |
| [33] | 李欣海. 随机森林模型在分类与回归分析中的应用. 应用昆虫学报 , 2013, 50 (4) : 1190–1197. |
| [34] | Fielding A H, Bell J F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environmental Conservation , 1997, 24 (1) : 38–49. DOI:10.1017/S0376892997000088 |
| [35] | Allouche O, Tsoar A, Kadmon R. Assessing the accuracy of species distribution models:prevalence, kappa and the true skill statistic (TSS). Journal of Applied Ecology , 2006, 43 (6) : 1223–1232. DOI:10.1111/jpe.2006.43.issue-6 |
| [36] | Monserud R A, Leemans R. Comparing global vegetation maps with the Kappa statistic. Ecological Modelling , 1992, 62 (4) : 275–293. DOI:10.1016/0304-3800(92)90003-W |
| [37] | 环境保护部. 中国生物多样性保护战略与行动计划. 北京: 中国环境科学出版社, 2011. |
| [38] | 齐丽丽, 柏延臣. 社会经济统计数据热点探测的MAUP效应. 地理学报 , 2012, 67 (10) : 1317–1326. |
| [39] | 张荣祖, 赵肯堂. 关于《中国动物地理区划》的修改. 动物学报 , 1978, 24 (2) : 196–202. |
| [40] | Zhang C S, Luo L, Xu W L, Ledwith V. Use of local Moran's I and GIS to identify pollution hotspots of Pb in urban soils of Galway, Ireland. Science of the Total Environment , 2008, 398 (1/3) : 212–221. |
| [41] | Hughey K F D, Cullen R, Moran E. Integrating economics into priority setting and evaluation in conservation management. Conservation Biology , 2003, 17 (1) : 93–103. DOI:10.1046/j.1523-1739.2003.01317.x |
| [42] | Moran P A P. Notes on continuous stochastic phenomena. Biometrika , 1950, 37 (1/2) : 17–23. DOI:10.2307/2332142 |
| [43] | Li H F, Calder C A, Cressie N. Beyond Moran's I:testing for spatial dependence based on the spatial autoregressive model. Geographical Analysis , 2007, 39 (4) : 357–375. DOI:10.1111/gean.2007.39.issue-4 |
| [44] | 刘吉平, 吕宪国. 三江平原湿地鸟类丰富度的空间格局及热点地区保护. 生态学报 , 2011, 31 (20) : 5894–5902. |
| [45] | 赵广华, 田瑜, 唐志尧, 李俊生, 曾辉. 中国国家级陆地自然保护区分布及其与人类活动和自然环境的关系. 生物多样性 , 2013, 21 (6) : 658–665. |
| [46] | Myers N, Mittermeier R A, Mittermeier C G, da Fonseca G A B, Kent J. Biodiversity hotspots for conservation priorities. Nature , 2000, 403 (6772) : 853–858. DOI:10.1038/35002501 |
| [47] | Ma Z J, Melville D S, Liu J G, Chen Y, Yang H Y, Ren W W, Zhang Z W, Piersma T, Li B. Rethinking China's new great wall:massive seawall construction in coastal wetlands threatens biodiversity. Science , 2014, 346 (6212) : 912–914. DOI:10.1126/science.1257258 |
| [48] | 马志军, 李博, 陈家宽. 迁徙鸟类对中途停歇地的利用及迁徙对策. 生态学报 , 2005, 25 (6) : 1404–1412. |