 2016, Vol. 36
2016, Vol. 36文章信息
- 闫琰, 姚杰, 张新娜, 张春雨, 赵秀海
- YAN Yan, YAO Jie, Zhang Xinna, ZHANG Chunyu, ZHAO Xiuhai.
- 吉林蛟河不同演替阶段针阔混交林木本植物幼苗空间分布与年际动态
- Spatial distribution and interannual dynamics of tree seedlings at different successional stages in a conifer and broad-leaved mixed forest in Jiaohe, Jilin Province, China
- 生态学报[J]. 2016, 36(23): 7644-7654
- Acta Ecologica Sinica[J]. 2016, 36(23): 7644-7654
- http://dx.doi.org/10.5846/stxb201512062442
-
文章历史
- 收稿日期: 2015-12-06
- 修订日期: 2016-05-30

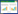




天然林更新主要是指植物从开花, 结实到种子的生产、扩散和萌发, 以及幼苗的定居、存活、生长和建成的生态学过程[1-2]。其中, 幼苗被认为是森林更新过程对外界环境最敏感的时期[3-4], 其建植成功与否直接关系到未来植物群落的更新[5]。与此同时幼苗定居对植被的更新具有筛选作用[6], 通过影响植物种群的数量组成、分布格局来影响森群落动态变化, 最终决定了森林的长期演替格局, 维系了森林生态系统植被的多样性[7]。
空间格局与生态学过程之间的相互作用是群落发展的动力[8]。幼苗作为林下生态系统的重要组分, 是木本植物生活史中生死过程和动态特性最明显的时期[9], 其空间分布格局会对幼苗的自然更新过程会产生重要的影响。在此过程中, 一些机制能够促进生物多样性的维持[5]。Swamy等认为, 根据Janzen-Connell假说, 树种的更新格局总是偏斜并远离于母树[10]。这是由于寄主特异性天敌, 如病原体、专性植食者等, 容易从成年个体扩散到离它们较近的后代; 同时, 由于大多数树种的种子散布在母树周围, 分布于母树附近的同种幼苗个体过于密集, 病原体和专性植食者的繁殖很可能会加剧, 使得幼苗因负密度制约(Negative density dependence)而导致大量死亡[11-12]。这两个过程使幼苗的存活率随离母树或同种成树距离的增加而升高, 在制约同种幼苗增补的同时, 为其他物种的幼苗更新提供了空间, 从而维持了物种共存。此外, Queenborough等也认为幼苗的空间格局对物种的长期分布格局有着重要影响[13]。
另一方面, 森林的更新格局与林分的空间异质性有重要的关系[8]。在不同类型的群落中, 地上成体植株的空间异质性为幼苗的增补、定植等过程提供了重要基础, 影响着整个群落的更新特征和演替空间模式[14]。然而, 以往对幼苗空间分布格局的研究多集中于特定群落类型[15-17], 对不同演替阶段群落中幼苗空间分布格局的研究鲜见报道。
以阔叶红松(Pinus koraiensis)林为演替顶级的次生针阔混交林是我国北温带地区重要的地带性植被类型, 吉林省蛟河地区是阔叶红松林的主要分布区之一。早期的人为干扰, 如过渡采伐、放牧等活动使大量原始森林遭到破坏, 退化成次生林群落。近年来在国家“天然林保护”与“退耕还林”等生态政策的推动下, 这些林分形成了针阔混交林次生演替过程中的不同群落类型。尽管如此, 森林的天然更新仍然是当地植被恢复过程中面临的首要难题。因此, 本文以吉林省蛟河地区不同演替阶段的针阔混交林固定监测样地为基础, 对2012-2014年连续3 a的木本植物幼苗调查数进行了初分析, 旨在:(1) 探讨木本植物幼苗的空间分布规律及其年际间差异; (2) 探讨木本植物幼苗和成树在空间分布上的相关性规律。以揭示温带地区不同演替阶段森林群落的更新规律, 为该地区生态恢复提供理论基础。
1 研究地概况与研究方法 1.1 研究区概况蛟河林业试验区管理局林场位于吉林省蛟河市前进乡境内(43°51′-44°05′N, 127°35′-127°51′E, 海拔330-1176 m), 属长白山系张广才岭山脉。该地区植被属于长白山植物区系, 天然次生针阔混交林和原始阔叶红松林是当地的主要植被类型[18]。乔木层树种主要包括红松、杉松、东北枫、五角枫、胡桃楸(Juglans mandshurica)、水曲柳和紫椴(Tilia amurensis)等。林下灌木层树种主要有暴马丁香(Syringa reticulatavar.mandshurica)、毛榛(Corylus mandshurica)、东北山梅花(Philadelphus schrenkii)和刺五加(Acanthopanax senticosus)等。主要草本有美汉草(Meehania fargesii)、狭叶荨麻(Urtica angustifolia)、荷青花(Hylomecon japonica)、华东蹄盖蕨(Athyrium niponicum)和猴腿蹄盖蕨(Athyrium brevifrons)等。该地区处于中纬度北温带, 属受季风影响的温带大陆性气候, 春季干旱而多大风, 夏季炎热而多雨水, 秋季短暂而多晴天, 冬季寒冷漫长。年平均气温3.8℃, 最热月7月日均温为21.7℃, 最冷月1月份平均温度-18.6℃。降水多集中在夏季, 约为700-800 mm。土壤类型主要为暗棕色森林土[19]。
1.2 样地建立与调查根据CTFS (Center for Tropical Forest Science)的技术规范(http://www.ctfs.si.edu/), 2010年在研究区内根据林分的发育阶段和树种组成, 选取3种不同发育阶段的针阔混交林建立固定监测样地, 包括中龄林(21.84 hm2, 420 m×520 m)、成熟林(42 hm2, 500 m×840 m)和老龄林(30 hm2, 500 m×600 m), 样地概况如表 1所示。3块样地形成了一个由次生林逐渐向顶级植被类型过渡的典型演替系列。调查记录各样地内所有胸径(DBH)≥1 cm的植株个体物种名称、胸径(cm)、树高(m)、枝下高(m)和冠幅(m), 并给每个植株挂牌、定位。
| 林型 Forest type |
样地面积 Plot area |
经纬度 Longitude and latitude |
海拔/m Elevation |
样站数 No. of stations |
演替阶段 Succession stages |
原始植被 Primary vegetation |
| HF | 21.84 hm2 (420 m×520 m) |
43°58.383′N 127°44.317′E |
468-519 | 99 | 初期 | 阔叶红松林 |
| MF | 42.00 hm2 (500 m×840 m) |
43°57.783′N 127°44.389′E |
459-517 | 209 | 中期 | |
| OGF | 30.00 hm2 (500 m×600 m) |
43°58.071′N 127°45.539′E |
576-784 | 143 | 后期 | |
| HF:中龄林half-mature forest; MF:成熟林mature forest; OGF:老龄林old-growth forest | ||||||
2011年7月采用机械布点的方法, 在3个样地内共设置了451个“种子雨-幼苗”观测样站, 其中中龄林99个、成熟林209个、老龄林143个(图 1)。每个样站由1个0.8 m×0.8 m的种子雨收集器及其四周距其1 m处的4个1 m×1 m幼苗小样方组成(图 2)。

|
| 图 1 样站分布示意图 Fig. 1 The layout of monitoring stations |

|
| 图 2 样站幼苗小样方设置示意图 Fig. 2 Quadrat layout at each of the sampling station |
对苗区内胸径小于1 cm的木本植物幼苗进行调查, 记录每棵幼苗的物种名称、年龄、高度和基径, 同时对每颗幼苗进行挂牌标记。幼苗的年龄是根据芽鳞痕的个数来确定。幼苗高度为自然状态下基部到最高顶芽的距离。2012年7-8月进行了第1次幼苗调查工作, 以后每年7-8月复查调查1次, 并记录幼苗的出生和死亡状态[20]。本文选取2012-2014年间的3次幼苗调查数据进行分析。
1.5 数据分析方法根据各样站内的幼苗数量(N)划分为不同的等级: N0=0; 0 < N1≤5; 6≤N2≤10; 11≤N3≤15; 16≤N4≤20; 21≤N5≤25; 26≤N6≤30; 31≤N7≤35; 36≤N8≤40; 41≤N9≤45; 46≤N10≤50; 51≤N11≤55; 56≤N12≤60; 61≤N13≤65; 66≤N14≤70; 71≤N15≤75; 76≤N16≤80; 81≤N17≤85; 86≤N18≤90; 91≤N19≤95; 96≤N20≤100; N21 > 100。统计各样站内每个等级的幼苗个体数。
选取Syrjala检验[21], 来分析样站尺度幼苗密度和丰富度的空间分布在年际间的差异。Syrjala检验是通过计算两个样本的Cramer-von Mises检验来分析两样本之间的空间分布差异。Syrjala检验的零假设是在整个研究区域内, 两样本的正态分布是一致的。设在取样点k的坐标为(xk, yk), di(xk, yk)为第i个种群k点的密度值。构建一个独立于种群大小的检验, 所观测到的密度数据首先被标准化:
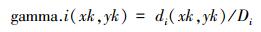
|
式中, Di是di(xk, yk)观测值的总和。第i个种群在点(xk, yk)处的累积分布的值, 也就是GAMMA.i(xk, yk), 是所有正态密度观察值的总和, gamma.i(xk, yk)。由Syrjala检验零假设的统计量为两个种群之间累积分布的差值的平方, 然后所有样本点求和, 即,
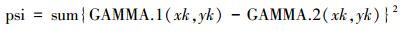
|
psi是根据研究区域矩形的角点而改变的, psi被计算4次, 每1个角点1次, 所得的平均psi值用来做检验提供假。观察到的psi显著性尺度是由它在所有2K个成对排列所得的检验统计值中的序列位置而决定的。
本文中所有数据分析与绘图均采用国际通用软件R-3.2.0 (http://www.r-project.org)和Microsoft Excel 2010完成。
2 结果与分析 2.1 种子雨组成2012-2014年间, 中龄林共收集到14个物种的种子, 隶属于8科10属; 成熟林共收集到21个物种的种子, 隶属于11科15属; 老龄林共收集到20个物种的种子, 隶属于11科14属20种植物(表 2)。3个样地中, 多数物种的种子雨密度都是在2013年达到峰值, 但也有少数物种如中龄林中的杉松、胡桃楸, 成熟林中的辽椴和老龄林中的千金榆等的种子雨密度却在2013年最低(表 2)。
| 物种Species | HF | MF | OGF | ||||||
| 2012 | 2013 | 2014 | 2012 | 2013 | 2014 | 2012 | 2013 | 2014 | |
| 紫椴Tilia amurensis | 245.72 | 262.26 | 122.54 | 436.32 | 503.91 | 525.58 | 392.01 | 401.34 | 195.61 |
| 白桦Betula platyphylla | - | 33.36 | 22.60 | 47.55 | 1385.65 | 465.89 | 1.85 | 10.04 | - |
| 水曲柳Fraxinus mandshurica | 46.04 | 357.28 | 43.62 | 261.76 | 666.81 | 21.28 | 9.99 | 121.05 | 8.84 |
| 五角枫Acer pictumsubsp. mono | 9.26 | 26.58 | 4.04 | 136.11 | 407.64 | 86.22 | 133.89 | 249.72 | 10.18 |
| 枫桦Betula costata. | - | 0.46 | 2.67 | - | 119.33 | 253.88 | - | 229.43 | 344.78 |
| 千金榆Carpinus cordata | 0.28 | - | 0.32 | 41.40 | 0.35 | 40.91 | 90.47 | 0.80 | 104.96 |
| 东北枫Acer mandshuricum | 3.65 | 6.53 | 3.93 | 17.55 | 34.58 | 29.39 | 33.43 | 69.13 | 23.22 |
| 黄檗Phellodendron amurense | - | - | - | 0.05 | 0.22 | - | 89.16 | 51.82 | 72.92 |
| 春榆Ulmus davidianavar. japonica | - | 0.22 | 67.03 | - | 0.02 | 19.19 | - | 2.05 | 26.93 |
| 辽椴Tilia mandshurica | 0.03 | - | - | 45.08 | 1.88 | 33.43 | 7.75 | 1.29 | 5.60 |
| 蒙古栎Quercus mongolica | - | - | - | 4.42 | 19.26 | 12.36 | 9.20 | 11.00 | 1.91 |
| 青楷枫Acer tegmentosum | 0.03 | 0.38 | 0.11 | 0.77 | 3.38 | 2.59 | 0.27 | 26.75 | 6.20 |
| 沙松Abies holophylla | 1.85 | 0.63 | 2.18 | 0.14 | 3.39 | 5.79 | 1.03 | 10.48 | 2.59 |
| 花楸Sorbus pohuashanensis | - | - | - | 0.14 | 0.41 | 2.41 | 2.70 | 1.70 | 5.82 |
| 稠李Padus racemosa | 0.16 | 0.06 | - | 0.03 | - | - | 4.31 | 2.38 | 0.71 |
| 三花枫Acer triflorum | - | - | - | - | 3.00 | 0.65 | - | 0.30 | - |
| 胡桃楸Juglans mandshurica | 0.69 | 0.43 | 0.63 | 0.38 | 0.35 | 0.65 | 0.25 | 0.25 | 0.16 |
| 软枣猕猴桃Actinidia arguta | - | - | - | - | 0.08 | 0.51 | - | 2.34 | 0.24 |
| 簇毛枫Acer barbinerve | - | - | - | - | 0.08 | 0.09 | - | 1.25 | 0.36 |
| 红松Pinus koraiensis | 0.02 | - | - | 0.02 | 0.09 | 0.80 | - | 0.02 | 0.16 |
| 长白落叶松Larix olgensis | - | - | - | 0.02 | - | - | - | - | - |
| 合计Total | 307.73 | 688.19 | 269.67 | 991.73 | 3150.43 | 1501.63 | 776.29 | 1193.15 | 811.17 |
研究期间, 3个样地内调查到的木本植物幼苗均为该样地存在的主要树种, 没有发现其他树种的幼苗。样地内幼苗总数都在2013年达到最大, 2014年幼苗数量最少, 约为2012年、2013年幼苗数量的一半(表 3)。2012-2014年间, 中龄林共监测到10科11属18种木本植物幼苗, 其中2012年调查到16种1338株幼苗, 没有发现辽椴(Tilia mandshurica)和千金榆; 2013年调查到18个物种的幼苗, 共1942株; 而2014年共调查到724株17种幼苗, 没有发现辽椴(表 3)。成熟林共监测到9科11属17种木本植物幼苗, 其中2012年调查到17种幼苗, 共4041株; 2013和2014年分别调查到5160和2379株幼苗, 但都没有发现毛榛(Corylus mandshurica, 表 3)。老龄林3年内均调查到16个相同树种的幼苗, 隶属于9科10属, 幼苗株树分别为2199、2229和917(表 3)。
| 物种Species | HF | MF | OGF | ||||||
| 2012 | 2013 | 2014 | 2012 | 2013 | 2014 | 2012 | 2013 | 2014 | |
| 水曲柳Fraxinus mandshurica | 51.1(88.9) | 34.2(87.9) | 36.9(61.6) | 34.7(76.6) | 26.4(79.4) | 30.2(59.8) | 3.2(28.0) | 1.9(16.1) | 2.2(10.5) |
| 杉松Abies holophylla | 23.2(19.2) | 6.3(19.2) | 6.5(13.1) | 34.4(42.1) | 3.4(20.1) | 2.5(15.3) | 45.0(51.7) | 3.2(18.2) | 6.3(23.1) |
| 五角枫Acer pictumsubsp. mono | 8.8(46.5) | 26.2(69.7) | 21.4(51.5) | 13.3(72.7) | 32.4(90.9) | 26.4(73.7) | 13.4(66.4) | 33.2(83.9) | 28.1(62.2) |
| 东北枫Acer mandshuricum | 4.1(33.3) | 21.2(69.7) | 15.6(42.4) | 3.5(37.8) | 16.2(75.1) | 10.0(51.2) | 7.9(55.2) | 18.8(66.4) | 17.9(48.3) |
| 胡桃楸Juglans mandshurica | 3.4(23.2) | 1.3(14.1) | 1.9(10.1) | 0.7(10.0) | 0.2(4.3) | 0.2(1.9) | 0.5(5.6) | 0.1(1.4) | 0.1(0.7) |
| 红松Pinus koraiensis | 2.1(9.1) | 0.4(3.0) | 0.3(2.0) | 2.2(23.4) | 1.0(13.9) | 1.1(10.0) | 1.7(21.0) | 1.0(11.9) | 1.6(9.1) |
| 春榆Ulmus davidiana var. japonica | 2.0(6.1) | 2.5(17.2) | 4.8(8.1) | 0.6(7.7) | 1.1(14.8) | 1.0(8.1) | 12.6(49.7) | 6.0(30.1) | 7.0(18.9) |
| 簇毛枫Acer barbinerve | 1.7(7.1) | 1.1(6.1) | 1.5(3.0) | 3.2(13.9) | 2.7(18.7) | 4.8(14.4) | 5.5(23.1) | 9.2(37.8) | 6.4(15.4) |
| 青杨Populus cathayana | 1.7(1.0) | 2.6(1.0) | 5.1(1.0) | - | - | - | - | - | - |
| 紫椴Tilia amurensis | 0.7(4.0) | 2.1(9.1) | 4.0(14.1) | 2.0(18.2) | 8.6(36.4) | 19.5(57.4) | 1.3(13.3) | 4.4(21.7) | 14.8(41.3) |
| 裂叶榆Ulmus laciniata | 0.3(3.0) | 0.3(5.1) | 0.7(4.0) | 0.7(10.5) | 0.5(9.6) | 1.0(8.6) | 1.6(9.8) | 1.3(9.1) | 2.3(7.7) |
| 黄檗Phellodendron amurense | 0.2(2.0) | 0.2(2.0) | 0.3(1.0) | 0.0(0.5) | 0.0(0.5) | 0.1(0.5) | 0.5(4.9) | 1.0(10.5) | 1.3(4.9) |
| 青楷枫Acer tegmentosum | 0.2(2.0) | 1.3(8.1) | 0.4(3.0) | 0.6(5.7) | 2.8(29.2) | 0.8(7.2) | 1.0(7.7) | 11.8(53.8) | 4.8(20.3) |
| 稠李Prunus avium | 0.1(1.0) | 0.1(1.0) | 0.1(1.0) | - | - | - | - | - | - |
| 山杨Populus davidiana | 0.1(1.0) | 0.1(1.0) | 0.1(1.0) | - | - | - | - | - | - |
| 乌苏里鼠李Rhamnus ussuriensis | 0.1(1.0) | 0.1(1.0) | 0.1(1.0) | - | - | - | - | - | - |
| 辽椴Tilia mandshurica | - | 0.1(1.0) | - | 2.9(23.4) | 2.3(23.0) | 0.7(4.3) | 3.3(26.6) | 1.6(11.2) | 0.4(2.8) |
| 千金榆Carpinus cordata | - | 0.3(4.0) | 0.1(1.0) | 0.6(8.1) | 2.0(25.8) | 1.5(11.5) | 2.0(16.1) | 6.1(28.7) | 6.3(20.3) |
| 毛榛Corylus mandshurica | 0.1(0.5) | ||||||||
| 蒙古栎Quercus mongolicus | - | - | - | 0.2(4.3) | 0.1(2.4) | 0.1(1.4) | 0.5(4.2) | 0.2(2.8) | 0.2(1.4) |
| 三花枫Acer triflorum | - | - | - | 0.1(2.9) | 0.4(6.7) | 0.2(1.4) | 0.0(0.7) | 0.2(2.8) | 0.1(0.7) |
| 总计Total | 1338 | 1942 | 724 | 4041 | 5160 | 2379 | 2199 | 2229 | 917 |
各样地树种幼苗数量与出现的样方数比较结果表明:水曲柳、五角枫和东北枫等幼苗数量较多的物种, 出现的样方数也相对较多(表 3)。3次调查期间, 中龄林水曲柳幼苗数量最多, 出现样站的比例均在61.6%以上, 其中2012年和2013年出现的样站数明显多于2014年; 成熟林水曲柳、五角枫和东北枫出现的样站数分别达到总样站数的59.8%、72.7%和37.8%以上; 老龄林中五角枫和东北枫出现的样站数分别达到总样站数的62.2%和48.3%以上。3个样地中, 尽管2012年的杉松幼苗数量远高于五角枫, 但其出现的样站数明显小于五角枫(表 3)。
根据幼苗出现样站相对频率分布可知:在2012-2014年间, 3个样地中分绝大多数样站出现幼苗数为1-10株, 并且都呈现出偏峰现象; 2014年3个样地均以幼苗数为1-5株的样站最多, 并且幼苗数为0株和1-5株的样站数量均明显高于2012年和2013年; 成熟林在2013年幼苗数为6-10株的样站数最多, 而老龄林在2012年幼苗数为6-10株的样站数最多(图 3)。

|
| 图 3 各样站出现的幼苗个体数和物种数 Fig. 3 The number of individuals and species in each of station |
对样站中出现幼苗的物种数进行统计发现, 中龄林在3次调查期间物种的频率分布均出现偏锋现象, 而且年际间差异明显: 2012年有6个样站中没有发现任何树种幼苗, 85个样站(占样站总数的85.9%)中出现的物种数小于5; 2013年出现1-5个幼苗物种的样站数最多, 为29个, 远高于2012年和2014年调查结果; 2014年有18个样站中没有出现任何幼苗, 61个样站(样站总数的61.6%)发现0-2个物种(图 3)。而成熟林和老龄林物种频率分布的年际差异不明显, 且均为正态分布, 样站数最多的为3-4个树种(图 3)。
2.3 幼苗空间分布差异采用Syrjala检验分析了同一样地内幼苗个体总数、物种数以及4个主要物种幼苗多度的年际间空间分布差异(表 4)。结果表明:中龄林幼苗物种数和个体数的空间分布在2013年和2014年存在显著差异; 水曲柳、杉松和东北枫幼苗数量空间分布在年际间存在显著差异, 而五角枫幼苗数量在年际间差异不显著, 表明五角枫幼苗的产生和分布在中龄林中比较稳定。成熟林幼苗物种数和幼苗个体总数的空间分布存在显著的年际差异, 水曲柳、五角枫和东北枫幼苗数量的空间分布在年际间差异显著, 而杉松幼苗数量在年际间差异不显著, 表明杉松幼苗的产生和分布在成熟林中比较稳定。老龄林中, 2012年幼苗个体总数的空间分布与2013年、2014年差异显著; 2012年杉松幼苗数量空间分布与2013年差异显著; 而幼苗的物种数以及水曲柳、五角枫和东北枫幼苗数量的空间分布在年际间变化不显著, 表明老龄林林下幼苗的空间分布比中龄林和成熟林相对稳定。
| 林型 Forest type |
年组 Year group |
物种数 No. of species |
个体数 No. of individuals |
水曲柳 Fraxinus mandshurica |
五角枫 Acer mono |
杉松 Abies holophylla |
东北枫 Acer mandshuricum |
| HF | 2012-03 | 0.018 | 0.168 | 0.035 | 0.319 | 0.395 | 0.331 |
| 2012-04 | 0.223 | 0.377 | 0.362* | 0.128 | 1.193* | 0.949 | |
| 2013-04 | 0.189** | 0.737** | 0.560** | 0.475 | 0.329 | 1.827** | |
| MF | 2012-03 | 0.038 | 0.847* | 0.144* | 1.252** | 0.365 | 0.498 |
| 2012-04 | 0.124* | 0.999* | 0.339 | 1.009* | 0.5 | 1.921* | |
| 2013-04 | 0.053 | 0.171 | 0.154 | 0.611* | 0.554 | 0.881** | |
| OGF | 2012-03 | 0.053 | 1.360* | 0.288 | 0.522 | 1.790* | 0.363 |
| 2012-04 | 0.034 | 1.429* | 0.405 | 0.339 | 1.27 | 0.333 | |
| 2013-04 | 0.058 | 0.067 | 0.659 | 0.128 | 0.743 | 0.125 |
对3个样地内水曲柳、五角枫、杉松和东北枫的死亡幼苗和成树空间分布进行比较可以发现:尽管3个样地中死亡的幼苗基本分布在母树附近, 并且整体格局与母树保持一致, 但死亡幼苗的空间分布格局在年际间存在一定差异(图 4-图 6); 2013-2014年间五角枫和东北枫的死亡幼苗在3个样地中分布较为广泛(图 4, 图 5); 老龄林中, 杉松在2013-2014年间死亡的幼苗在样地中分布的较少(图 6)。

|
| 图 4 2012-2013年、2013-2014年和2012-2014年间中龄林4个主要树种的死亡幼苗及其胸径大于20 cm母树的空间分布格局 Fig. 4 Spatial distribution patterns of the dead seedlings for the four main species during the year of 2012-2013, 2013-2014 and 2012-2014, and their parent trees with DBH≥20 cm in the HF 灰色叉号为母树空间分布, 黑色方块为出现该树种幼苗死亡的样站分布 |

|
| 图 5 2012-2013年、2013-2014年和2012-2014年间成熟林4个主要树种的死亡幼苗及其胸径大于20 cm母树的空间分布格局 Fig. 5 Spatial distribution patterns of the dead seedlings for the four main species during the year of 2012-2013, 2013-2014 and 2012-2014, and their parent trees with DBH≥20 cm in the MF 灰色叉号为母树空间分布, 黑色方块为出现该树种幼苗死亡的样站分布 |

|
| 图 6 2012-2013年、2013-2014年和2012-2014年间老龄林4个主要树种的死亡幼苗及其胸径大于20 cm母树的空间分布格局 Fig. 6 Spatial distribution patterns of the dead seedlings for the four main species during the year of 2012-2013, 2013-2014 and 2012-2014, and their parent trees with DBH≥20 cm in the OGF 灰色叉号为母树空间分布, 黑色方块为出现该树种幼苗死亡的样站分布 |
群落幼苗与地上成体植株的空间分布格局决定了群落更新特征, 成体植株的空间分布特征对幼苗建成具有重要影响[14]。因此, 幼苗的空间分布对群落更新具有重要意义。本文通过对蛟河地区不同演替阶段针阔混交林样地内木本植物及幼苗的调查分析, 发现3个样地内调查到的木本植物幼苗均为该样地存在的主要树种, 没有发现其他树种的幼苗。木本植物幼苗的数量分布在不同演替阶段之间都表现出明显的差异:水曲柳在演替初期和中期的林分中出现频率较大, 五角枫在演替中期和后期的林分中中出现频率最大, 东北枫在3个样地中出现频率都比较大; 而胡桃楸、杉松和红松的幼苗在3个样地中出现的频率都很低。这一方面可能是由于不同演替阶段的林分中, 树种的数量和组成存在差异, 使得不同物种的幼苗分布在不同群落间存在差异; 另一方面, 啮齿类动物和人类对红松、胡桃楸等种子的取食和采摘也会对幼苗的分布造成不同程度的影响。研究期间在3个样地内水曲柳、五角枫和东北枫的种子数量在所有树种中均排在前5位, 而胡桃楸、杉松和红松的种子数量较少(未发表数据), 这与陈贝贝等部分研究结果一致[22]; 此外, 幼苗的空间分布可能与种子的传播特性有关, 水曲柳、五角枫和东北枫的种子主要通过风力传播, 良好的扩散能力也能使得种子和幼苗在样地内广泛分布[23], 而胡桃楸、红松和杉松的种子主要通过重力传播, 扩散能力有限。
3.2 幼苗密度与丰富度的空间异质性本研究结果显示, 在4 m2样站尺度上3个不同演替阶段的群落中幼苗密度存在着较大的空间变异性, 而幼苗丰富度的空间异质性较低。这与李晓亮等对西双版纳热带森林树种幼苗空间分布研究的部分结果一致[24], 他们认为幼苗密度的空间变异是由微生境的空间异质性导致的, 微生境的空间变异又会反来影响幼苗的存活[25]。而物种数量由于受群落物种区系控制, 空间异质性较低[14]。也有学者认为森林中幼苗空间格局可能比其周围的环境因素更重要[16], 因为幼苗密度的空间格局会影响到幼苗的存活状态[24]。例如, Janzen和Connell[11-12]认为分布于母树附近的幼苗由于受到负密度制约效应会表现出较高的死亡率, 而本研究中3个样地内主要树种的幼苗都集中于母树附近(图 3-图 5)。同时, 以往的研究已经证实了负密度制约效应是该研究地区幼苗存活的主要驱动机制[20], 这在一定程度上可以反映出幼苗分布的空间异质性对幼苗密度的影响。
3.3 幼苗空间分布的年际差异不同年份之间幼苗个体数和物种数的空间分布是存在差异的。这主要与种子的产量与扩散有关。首先, 种子是幼苗更新的来源, 幼苗的数量和分布很大程度上依赖于种子的可获得性[26], 种子数量多的物种跟容易扩散到更多的样站[27]。另一方面, 不同物种的种子扩散方式有很大差异, 有些物种的种子在经历了种子雨扩散之后, 还会经历二次扩散。二次扩散可以降低种子的聚集程度, 使种子距离母树更远。因此, 种子扩散会导致种子萌发位置的改变进而影响幼苗的分布。同时, 种子具有休眠的特性, 只有光照、温度等环境因素适宜的时候种子才会萌发, 这也影响到幼苗的分布格局。除此之外, 不同演替阶段群落中的物种组成和林分结构及环境都有很大的差异, 因此各林分内幼苗空间分布在年际间的差异也不尽相同。
3.4 死亡幼苗空间分布与成树关系除了种子产量和种子扩散方式, 成体植株的空间分布特征也能够影响幼苗的空间分布格局[28]。本研究中, 在不同演替阶段的群落内, 水曲柳、五角枫、杉松和东北枫这4个主要树种的死亡幼苗在不同年际间的空间分布格局与其母树分布基本一致。这可能是由于选取的4个物种的母树在群落中有较多的个体, 产生的种子量较多, 使母树附近有足够的种源, 进而使得幼苗与母树有着相似的空间分布格局[24]; 另一方面, 由于种子的传播限制, 大多数树种并不能把种子散布到离母树较远的地方[29-30], 相反, 随着与母树距离的增加, 种子密度会逐渐降低[31], 虽然这4个树种的种子都依靠风力传播, 但是传播的距离并不远, 促使幼苗聚集在母树周围。因此, 在负密度制约效应的作用下, 邻近母树的幼苗更容易死亡。此外, 五角枫、东北枫和沙松的死亡幼苗的空间分布在年际间存在差异, 这可能与幼苗的密度有关。3个样地中, 五角枫和东北枫在2013-2014年间死亡幼苗的空间分布要明显大于2012-2013年, 而这两个物种在2013年的幼苗密度均大于2012年, 这从侧面进一步证实了负密度制约效应对幼苗空间分布的影响。
| [1] | 郭秋菊.择伐和火干扰对长叶松幼苗更新的影响[D].杨凌:西北农林科技大学, 2013. http://cdmd.cnki.com.cn/Article/CDMD-10712-1014162926.htm |
| [2] | Rockwood L L. Introduction to Population Ecology. Oxford: Blackwell Publishing, 2006. |
| [3] | 尹华军, 程新颖, 赖挺, 林波, 刘庆. 川西亚高山65年人工云杉林种子雨、种子库和幼苗定居研究. 植物生态学报 , 2011, 35 (1) : 35–44. |
| [4] | Du X J, Guo Q F, Gao X M, Ma K P. Seed rain, soil seed bank, seed loss and regeneration of Castanopsis fargesii(Fagaceae) in a subtropical evergreen broad-leaved forest. Forest Ecology and Management , 2007, 238 (1/3) : 212–219. |
| [5] | 苏嫄, 焦菊英, 王志杰. 陕北黄土丘陵沟壑区坡沟立地环境下幼苗的存活特征. 植物生态学报 , 2014, 38 (7) : 694–709. |
| [6] | Clark J S, Macklin E, Wood L. Stages and spatial scales of recruitment limitation in southern Appalachian forests. Ecological Monographs , 1998, 68 (2) : 213–235. DOI:10.1890/0012-9615(1998)068[0213:SASSOR]2.0.CO;2 |
| [7] | 康冰, 刘世荣, 王得祥, 张莹, 刘红茹, 杜焰玲. 秦岭山地典型次生林木本植物幼苗更新特征. 应用生态学报 , 2011, 22 (12) : 3123–3130. |
| [8] | 韩有志, 王政权. 天然次生林中水曲柳种子库的空间格局与过程. 植物生态学报 , 2002, 26 (2) : 170–176. |
| [9] | 路兴慧.海南岛热带低地雨林自然恢复过程中木本植物幼苗功能性状及增补动态[D].北京:中国林业科学研究院, 2012. http://cdmd.cnki.com.cn/Article/CDMD-82201-1012423729.htm |
| [10] | Swamy V, Terborgh J, Dexter K G, Best B D, Alvarez P, Cornejo F. Are all seeds equal? Spatially explicit comparisons of seed fall and sapling recruitment in a tropical forest. Ecology Letters , 2011, 14 (2) : 195–201. DOI:10.1111/j.1461-0248.2010.01571.x |
| [11] | Janzen D H. Herbivores and the number of tree species in tropical forests. The American Naturalists , 1970, 104 (940) : 501–528. DOI:10.1086/282687 |
| [12] | Connell J H. On the role of natural enemies in preventing competitive exclusion in some marin animals and rain forest trees//Den Boer P J, Gradwell G R, eds. Dynamics of Population. Wageningen:Center for Agricultural Publishing and Documentation, 1971:289-312. |
| [13] | Queenborough S A, Burslem D F R P, Garwood N C, Valencia R. Neighborhood and community interactions determine the spatial pattern of tropical tree seedling survival. Ecology , 2007, 88 (9) : 2248–2258. DOI:10.1890/06-0737.1 |
| [14] | 尚占环, 龙瑞军, 马玉寿, 丁路明. 青藏高原"黑土滩"次生毒杂草群落成体植株与幼苗空间异质性及相似性分析. 植物生态学报 , 2008, 32 (5) : 1157–1165. |
| [15] | 韩有志, 程志枫, 常杰, 王永强. 水曲柳人工林下天然更新幼苗的空间格局. 山西农业大学学报 , 2000 (4) : 335–338. |
| [16] | Armesto J J, Díaz I, Papic C, Willson M F. Seed rain of fleshy and dry propagules in different habitats in the temperate rainforests of Chiloé Island, Chile. Austral Ecology , 2001, 26 (4) : 311–320. DOI:10.1046/j.1442-9993.2001.01116.x |
| [17] | Piao T F, Comita L S, Jin G Z, Kin J H. Density dependence across multiple life stages in a temperate old-growth forest of northeast China. Oecologia , 2013, 172 (1) : 207–217. DOI:10.1007/s00442-012-2481-y |
| [18] | Zhao X H, Corral-Rivas J, Zhang C Y, Temesgen H, Gadow K V. Forest observational studies-an essential infrastructure for sustainable use of natural resources. Forest Ecosystems , 2014, 1 : 8. DOI:10.1186/2197-5620-1-8 |
| [19] | Zhang C Y, Jin W B, Gao L S, Zhao X H. Scale dependent structuring of spatial diversity in two temperate forest communities. Forest Ecology and Management , 2014, 316 : 110–116. DOI:10.1016/j.foreco.2013.07.025 |
| [20] | Yan Y, Zhang C Y, Wang Y X, Zhao X H, Von Gadow K. Drivers of seedling survival in a temperate forest and their relative importance at three stages of succession. Ecology and Evolution , 2015, 5 (19) : 4287–4299. DOI:10.1002/ece3.2015.5.issue-19 |
| [21] | Syrjala S E. A statistical test for a difference between the spatial distributions of two populations. Ecology , 1996, 77 (1) : 75–80. DOI:10.2307/2265656 |
| [22] | 陈贝贝, 赵秀海, 倪瑞强, 黄珍. 长白山阔叶红松林乔木树种幼苗组成与空间分布. 东北林业大学学报 , 2012, 40 (10) : 39–42. |
| [23] | 张健, 李步杭, 白雪娇, 原作强, 王绪高, 叶吉, 郝占庆. 长白山阔叶红松林乔木树种幼苗组成及其年际动态. 生物多样性 , 2009, 17 (4) : 385–396. |
| [24] | 李晓亮, 王洪, 郑征, 林露湘, 邓晓保, 曹敏. 西双版纳热带森林树种幼苗的组成、空间分布和旱季存活. 植物生态学报 , 2009, 33 (4) : 658–671. |
| [25] | Baraloto C, Goldberg D E. Microhabitat associations and seedling bank dynamics in a neotropical forest. Oecologia , 2004, 141 (4) : 701–712. DOI:10.1007/s00442-004-1691-3 |
| [26] | Nathan R, Safriel U N, Noy-Meir S, Schiller G. Spatiotemporal variation in seed dispersal and recruitment near and far from pinus halepensis trees. Ecology , 2000, 81 (8) : 2156–2169. DOI:10.1890/0012-9658(2000)081[2156:SVISDA]2.0.CO;2 |
| [27] | Chen L, Wang L, Baiketuerhan Y, Zhang C Y, Zhao X H, von Gadow K. Seed dispersal and seedling recruitment of trees at different successional stages in a temperate forest in northeastern China. Journal of Plant Ecology , 2013, 7 (4) : 337–346. |
| [28] | 王蕾.长白山针阔混交林种子雨动态及扩散限制研究[D].北京:北京林业大学, 2010. http://cdmd.cnki.com.cn/Article/CDMD-10022-2010128865.htm |
| [29] | 张健, 郝占庆, 李步杭, 叶吉, 王绪高, 姚晓琳. 长白山阔叶红松(Pinus koraiensis)林种子雨组成及其季节动态. 生态学报 , 2008, 28 (6) : 2445–2454. |
| [30] | 张玉波, 李景文, 张昊, 邹大林, 武逢平, 程春龙, 李俊清, 李帅英. 胡杨种子散布的时空分布格局. 生态学报 , 2005, 25 (8) : 1994–2000. |
| [31] | Willson M F, Whelan C J. Variation of dispersal phenology in a bird-dispersed shrub, cornus drummondii. Ecological Monographs , 1993, 63 (2) : 151–172. DOI:10.2307/2937178 |