 2016, Vol. 36
2016, Vol. 36文章信息
- 吴亚华, 肖荣波, 王刚, 黄柳菁, 邓一荣, 陈敏
- WU Yahua, XIAO Rongbo, WANG Gang, HUANG Liujing, DENG Yirong, CHEN Min.
- 城市绿地土壤呼吸速率的变化特征及其影响因子
- Controlling factors of variation in soil respiration rate in urban green-space ecosystems
- 生态学报[J]. 2016, 36(22): 7462-7471
- Acta Ecologica Sinica[J]. 2016, 36(22): 7462-7471
- http://dx.doi.org/10.5846/stxb201601080058
-
文章历史
- 收稿日期: 2016-01-08
- 修订日期: 2016-06-13





2. 广东省环境科学研究院, 广州 510045;
3. 福建农林大学, 福州 350002
2. Guangdong Provincial Academy of Environmental Science, Guangzhou 510045, China;
3. Fujian Agriculture and Forestry University, Fuzhou 350002, China
大气中以CO2为主的温室气体浓度持续升高, 导致全球气候变暖, 诱发一系列的生态环境问题, 如海平面上升、极地冰川融化和极端气象灾害等[1]。土壤呼吸过程可向大气中释放大量的CO2, 该过程包括土壤微生物、植物根系及土壤动物等的呼吸作用以及含碳物质的化学氧化作用[2], 是大气中CO2的重要来源之一。据估计, 土壤呼吸作用的CO2排放量是全球CO2排放总量的25%[3], 占陆地生态系统总排放量的60%-90%[4]。土壤呼吸作用的微小变化, 将导致大气中CO2浓度的巨大波动[5]。因此, 强化土壤呼吸作用的研究将有利于揭示不同生态系统在全球碳循环过程中的作用。
城市化进程正不断加快, 全球城市空间面积每年新增10万km2[6], 城市人口大量集聚, 导致物质和能源大量集中消耗和CO2排放量剧增。在城市化过程中, 公园绿地作为城市低碳与生态发展的重要载体, 是城市生态系统中的重要“碳汇”[7]。同时, 城市公园绿地也可通过土壤呼吸作用向大气中释放CO2, 对城市区域大气CO2浓度和碳循环过程有重要影响。由于受到人工管护和人类活动的综合影响, 城市绿地植被特征和土壤性质与自然生态系统差异较大, 可能导致土壤呼吸过程也存在较大差异。罗上华等[8]研究发现, 城市绿地土壤的碳通量比自然土壤和农业土壤的高。Schimel等[4]则发现在干旱和半干旱地区城市化引起了土壤呼吸强度的降低。
长期以来, 对非城市区域的森林、农田和草地等生态系统的土壤呼吸作用已有大量研究[9-10], 而关于城市公园绿地土壤呼吸过程也正日益受到国内外学者的关注。比如, 张香鸽等[11]对南京中山植物园内近自然林、疏林和草坪的土壤呼吸速率变化特征及其影响因子进行了研究;李丹丹等[12]对沈阳冬季城市草地的土壤呼吸速率变化及其与土壤温度和湿度的定量关系进行了研究。然而, 这些研究集中于北亚热带和温带区域, 对南亚热带地区城市绿地土壤呼吸作用的研究较少。
大量研究表明, 土壤呼吸过程的主要环境影响因子是土壤温度和土壤湿度[13-14]。多数研究表明土壤呼吸与土壤温度之间存在显著的相关性, 以指数模型最为典型。另外, 野外监测研究表明, 当土壤含水量过低或过高时, 其对土壤呼吸的影响更明显[15-16]。而事实上, 土壤呼吸过程不仅受到土壤温度和土壤湿度的影响, 同时还受到城市公园绿地植物类型及配置方式的影响[17], 城市不同植物的呼吸作用也对大气中碳循环过程具有一定的影响[18]。因此, 本研究选择广州市海珠湖公园的3种典型植被的土壤为研究对象, 综合分析不同植被类型的土壤呼吸速率的变化特征, 探讨土壤呼吸过程对土壤温度和土壤湿度变化的响应机制, 揭示引起城市绿地土壤呼吸速率季节变化的主要影响因子, 为优化布局城市绿地和实现低碳排放目标提供科学依据。
1 研究区域与研究方法 1.1 试验地概况试验地位于海珠湖公园(23°4′36"E, 113°19′5"N), 该公园是广州中心城区规模最大、保存最完整的湿地生态系统, 被称为广州的“南肾”, 公园面积149.89hm2。其中, 湖心区94.84hm2(水面面积53hm2, 陆地面积41.84hm2), 绿化配套用地55.05hm2。陆地部分是典型的乔灌草镶嵌结构, 有细叶榕(Ficus microcarpa)、樟树(Sect. Camphora)、人面子(Draconto melonduperreanum)、夹竹桃(Nerium indicum)、三角梅(Bougainvillea spectabilis)和马尼拉草(Zoysia matrella)等上百个品种的植物。
该区域气候属南亚热带海洋性季风气候, 雨量充沛, 光照充足, 温暖湿润, 全年温差较小, 干湿季节明显。全年气温较高, 年平均气温超过20℃。年降水量1500-2000mm, 干季(1-3月, 10-12月)降水量少;湿季(4-9月)降水量大, 占全年降水总量的80%-90%以上, 干湿两季分明。
1.2 试验设计为减小不同植被类型分布区域的小气候差异, 试验样地选定在公园唯一一处集中了疏林、灌丛和草地3种植被类型的东北角, 该区域人工管护水平基本一致, 土壤类型一致(图 1), 是开展本研究的理想之地。试验选取细叶榕(Ficus microcarpa)疏林、夹竹桃(Nerium indicum)灌丛和马尼拉草(Zoysia matrella)草地等3种植被类型的土壤为监测研究对象(图 1), 3种植被类型的土壤理化性质见表 1。采用典型样地法, 在3种植被类型中分别选取一个20m×20m标准样地, 在每个样地内随机设置3个2m×2m重复的样方, 并在每个样方中随机选取3个重复测定点。

|
| 图 1 试验样地分布位置 Fig. 1 Location of sample set |
| 样地编号 Sample plot code |
植被类型 Vegetation type |
优势植物 Dominant plant |
土壤体积 含水量 Soil water content/% |
土壤温度 Soil temperature/ ℃ |
土壤容重 Soil bulk density/ (g/m3) |
土壤pH | 土壤总氮 Total nitrogen/ (g/kg) |
土壤有机碳 Soil organic carbon/ (g/kg) |
| SL | 疏林 | 细叶榕(Ficus microcarpa) | 31.9±4.1 | 25.6±0.2 | 1.46±0.01 | 5.7±0.06 | 1.32±0.05 | 13.6±0.2 |
| GC | 灌丛 | 夹竹桃(Nerium indicum) | 28.9±2.0 | 25.6±0.4 | 1.41±0.01 | 5.9±0.12 | 1.21±0.06 | 12.1±0.4 |
| CD | 草地 | 马尼拉草(Zoysia matrella) | 32.8±3.3 | 27.8±0.5 | 1.63±0.01 | 5.5±0.11 | 1.33±0.05 | 12.9±0.5 |
试验在2013年11月至2014年10月间进行, 于每月下旬无雨的9:00至12:00[19], 采用静态箱-气相色谱法对土壤呼吸速率进行观测。2013年10月18日, 将高5cm的PVC材质的底座埋设到土壤监测点, 底座埋入土壤3cm。将底座保留在土壤中, 避免扰动土壤。监测采样时, 先清除底座中的凋落物, 然后将箱体(内径20cm, 高30cm)扣入底座的水槽中, 向水槽注水密封。箱内均装有空气搅拌器, 使箱内气体混合均匀。采样箱的顶部设有采气孔, 与采气三通阀相连接。分别在扣箱后0min、10 min、20min、30 min, 用注射器抽取采样箱内气体样品100mL转存于专用气袋内, 并迅速运回实验室及时分析。
在每次采集气体样品时, 同步监测研究样地内的风速、大气湿度、气温、大气压强、地表 10cm处土壤温度(T10)以及土壤体积含水量(VWC10)等环境因子。其中, 风速、大气湿度、气温和大气压强(1.5m)采用手持气象追踪仪测定(Kestrel4500, 美国);T10和VWC10采用路格温湿度记录仪(L93-7, 杭州路格科技有限公司)测定。
气体样品的测定采用安捷伦GC7890气相色谱仪。CO2检测采用离子火焰化检测器(FID), 工作温度为200℃;CO2的分析采用长度为2m的60-80目的PoraPak Q色谱柱(直径2mm), 工作柱温度55℃, 载气为高纯N2, 流速为30mL/min。
1.4 数据处理通过静态箱计算的CO2通量可用以下公式表示:
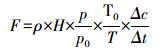
|
(1) |
式中, F为CO2气体交换通量(mg m-2 s-1), ρ为标准状态下CO2密度(1.963mg/mL);H为采样箱高度(m);P为采样点的气压(kPa);P0和T0分别为标准状态下的大气压和空气绝对温度, 即为101.3kPa和273.2K;T为采样时该点的温度(K);Δc/Δt为气体质量浓度随时间的变化率(m3 m-3 s-1);F为正值时表示土壤系统向大气排放CO2, 为负值时表示土壤系统从大气吸收CO2[20]。
整理观测的数据, 剔除异常值和由仪器故障造成的误差值。土壤温度敏感系数(Q10)为反映土壤呼吸(Rs)对温度敏感性的指数, 是温度每增加10℃所导致的Rs改变的熵, 当Rs与T之间存在指数关系时, Q10通过式(2)确定:
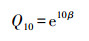
|
(2) |
式中, β为拟合参数。
采用SPSS19.0进行各个环境因子和土壤呼吸速率的相关性分析, 并对土壤呼吸速率与关键影响因子拟合, 采用Origin8.0软件绘制图表, 显著性差异水平设定为P=0.05。
2 结果与分析 2.1 3种植被类型土壤温度、土壤体积含水量及土壤呼吸速率的变化疏林、灌丛和草地3种植被类型土壤温度变化一致, 呈单峰曲线趋势。干季(10月至3月)土壤温度(T10)较低(低于24℃), 12月T10最低;湿季(4月至9月)T10较高, 7月或8月最高。同样, 3种植被类型土壤体积含水量(VWC10)的变化也几乎一致, 干季较低(疏林、灌丛和草地10-3月平均VWC10分别为26.6%、24.7%和24.0%);湿季较高(疏林、灌丛和草地4-9月平均VWC10分别为32.1%、31.8%和32.6%)。3种植被类型T10和VWC10均有较明显的干湿季节变化特点(图 2, P < 0.05), 与所处区域明显的干湿气候特点一致。

|
| 图 2 3种植被类型的Rs、T10和VWC10的季节变化 Fig. 2 Seasonal variations of Rs, T10, and VWC10 of three typical vegetation types |
在整个监测期内, 疏林、灌丛和草地3种植被类型的Rs月变化规律基本一致, 均呈较明显的单峰曲线(图 2a)。11月至2月间, 疏林和草地Rs波动下降, 灌丛Rs稳中有升。疏林和草地Rs在2月最小, 而灌丛的Rs在12月最小, 7月份均最大。然而, 在干湿季节变化过程中, Rs的变化规律有所差异。在干季, 3种植被类型Rs均较小, 且月变化波动幅度较小, 范围分别为(1.66±0.18)-(8.34±0.53) μmol m-2 s-1, (1.27±0.15)-(3.67±0.16) μmol m-2 s-1和(1.94±0.08)-(6.12±0.83) μmol m-2 s-1。在湿季, 3种植被类型Rs较大, 且月变化波动幅度较大, 范围分别为(3.54±0.46)-(13.81±1.31)μmol m-2 s-1, (2.82±0.22)-(12.72±1.16)μmol m-2 s-1和(3.53±0.68)-(9.82±0.96)μmol m-2 s-1, 湿季Rs变化幅度比干季的分别大4.59μmol m-2 s-1, 7.50μmol m-2 s-1和2.11μmol m-2 s-1。
2.2 土壤温度、体积含水量对土壤呼吸速率的影响如图 3所示, Rs随T10的升高而增大, 3种植被类型的Rs与T10具有显著的正相关关系(P < 0.01)。利用不同模型对Rs与T10之间的关系进行模拟分析[21-24], 指数方程(Rs=α·exp (βT10))最适用于描述该研究区域Rs与T10的关系。模拟分析结果表明, T10对疏林、灌丛和草地的Rs变化的解释程度分别可达到46.5%, 42.2%和37.4%。3种植被类型的Rs与VWC10关系比较复杂, 利用不同模型模拟分析发现, 三次曲线模拟效果相对较好, 但也只能解释疏林、灌丛和草地Rs变化的10.3%、23.1%和23.7%。

|
| 图 3 Rs与T10、VWC10的关系 Fig. 3 Relations between Rs and T10, VWC10 |
从图 2可以看出, 6月份灌丛和草地的Rs并未随着T10的升高而继续增大, 而是随着VWC10的上升而减小;同样, 高温的8月并未同7月一样保持较大Rs值, 而是随着VWC10的升高而急剧减小。从一定程度上说, T10和VWC10相互作用、相互影响, 共同作用于城市绿地的土壤呼吸过程。如图 4所示, 与单因子模型相比, 包含了T10和VWC10双因子的指数模型能更好的解释Rs变化, 其解释能力分别为54.6%、56.7%、50.8%。

|
| 图 4 T10、VWC10与Rs之间的关系 Fig. 4 The relationship between Rs and T10, VWC10 |
联合式(1)和(2)估算Q10, 对其全年和干湿两季的变化进行分析, 干湿两季土壤呼吸的温度敏感性不同, 湿季高于干季。如表 2所示, 3种植被类型湿季的Q10比干季的分别高0.44、0.70和0.46。从不同植被类型看, 在干季, 疏林的Q10明显高于灌丛和草地的, 灌丛的Q10最小;在湿季, 灌丛的Q10却最大, 草地的最小。
| 监测期间 Experiment period |
植被类型 vegetation types |
α | β | R2 | Q10 |
| 全年Full year | SL | 1.318 | 0.069 | 0.465 | 1.99 |
| GC | 1.439 | 0.064 | 0.422 | 1.90 | |
| CD | 1.456 | 0.054 | 0.374 | 1.72 | |
| SL | 0.7901 | 0.093 | 0.5381 | 2.55 | |
| 干季Dry season | GC | 1.6151 | 0.0211 | 0.0695 | 1.23 |
| CD | 1.6980 | 0.0388 | 0.1767 | 1.47 | |
| SL | 0.3986 | 0.1097 | 0.2768 | 2.99 | |
| 湿季Wet season | GC | 0.2301 | 0.1219 | 0.4025 | 3.38 |
| CD | 1.0019 | 0.0660 | 0.1823 | 1.93 |
已有研究表明陆地不同生态系统的Rs呈现明显的季节变化。本研究结果发现城市绿地3种植被类型的Rs变化呈明显的单峰曲线变化趋势, 且干湿季Rs差异显著, 湿季Rs较大, 干季Rs较小, 这与同纬度其它相关研究结果基本一致[25-26]。一般来说, Rs季节变化主要原因是研究区域外部环境因子的季节性改变。在干季, 低温少雨, 微生物活性低, 抑制了城市绿地的土壤呼吸作用, Rs下降;在湿季, 环境温度高, 降雨增多, 土壤温度和体积含水量均有所增加, 绿地植被及其土壤中的微生物新陈代谢作用旺盛, 土壤呼吸作用增强, Rs普遍增加。另外, Rs最小值一般出现在冬季, 最大值则出现在夏季[27-28]。本研究结果同样发现, 3种植被类型7月的Rs最大, 而12月或2月的最小。但值得注意的是与7月同样高温的8月, 其Rs却较低, Farque等[29]也有类似的发现。这主要是由于城市绿地Rs不仅受土壤温度的影响, 同时也受土壤体积含水量的影响。由于8月份的VWC10很高, 疏林、灌丛和草地的VWC10分别达到42.3%、36.8%和39.9%(分别为田间持水量的119.2%、111.6%和107.0%), 从而抑制了城市绿地土壤呼吸过程。Richard等[15]指出, 当土壤水分超过了田间持水量, 土壤CO2通量会大幅减少。
3.2 土壤温度、体积含水量对土壤呼吸的影响土壤温度可调控酶的活性, 而酶的活性变化可直接或间接地影响土壤中微生物代谢过程、植物根系分泌物及有机质的分解过程, 进而影响土壤呼吸过程。已有研究表明土壤温度是影响土壤呼吸过程的关键环境因子, 几乎影响土壤呼吸的全过程[13, 30]。与其它研究结果类似[21], 本研究发现Rs与T10呈明显的指数关系, 3种植被类型的T10对Rs变异的解释程度均在40%左右。对土壤呼吸过程而言, 土壤温度对土壤微生物活动的影响有着至关重要的作用。当土壤温度较低时, 土壤微生物活动受限, 优势微生物的数量少、丰度小;随着温度的升高, 参与呼吸的微生物数量和种类增加, 并且呼吸酶的活性也增强[31]。但也有研究[32-33]表明, 土壤温度对土壤呼吸过程的影响存在一定温度范围, 高于或低于这一范围, 指数关系不再适合描述两者的关系。谭炯锐等[32]的研究发现, 当土壤温度小于零度时, 土壤呼吸速率明显高于拟合方程求解的值, 土壤呼吸速率随土壤温度的升高而减小。另外, 疏林、灌丛和草地在整个观测期间的Q10分别为1.99、1.90和1.72, 均小于全球Q10平均值(2.4), 但处于我国陆地生态系统Q10范围内(1.28-4.75), 而且非常接近我国亚热带Q10的平均值(1.94)[21]。Luo等[31]认为, 土壤呼吸Q10因地理位置不同而具有较大差异的主要原因是不同地区气候温度不一样, 其土壤呼吸Q10对土壤温度的敏感性也就不同。此外, 本研究发现, 干湿两季Q10有明显的差异, 湿季高于干季。这是因为, 土壤水分条件也会影响土壤呼吸的温度敏感性, Reichstein等[34]发现, 湿度大的地方Q10比排水良好的地方的高。而Dorr和Munnich[35]在研究德国草地和云杉林土壤呼吸的温度敏感性时发现, 湿润年份Q10较低, 干旱年份的较高。
VWC10是影响城市绿地土壤呼吸作用的另一个重要环境因子, 其可直接影响土壤微生物及植被根系的生理过程, 也可间接影响底物和O2扩散, 影响机制较为复杂[36-37]。在本研究中, VWC10分别解释疏林、灌丛和草地Rs变异的10.3%、23.1%和23.7%, 明显低于土壤温度对Rs变异的解释程度。但值得注意的是, 在7、8月温度最高时, 8月较7月Rs有明显的下降, 并且6月灌丛和草地的Rs并未随T10升高而增大, 从图 2中可以看出6、8月的土壤体积含水量均超过30%, 此时土壤湿度可能已成为影响土壤呼吸作用的主导因子。由于影响土壤呼吸的因子除土壤温度和土壤湿度外, 还有土壤碳氮含量、植物和土壤孔隙度等, 这些因素又往往相互作用, 共同影响土壤呼吸[38], 因此在不同情况下, 土壤体积含水量影响土壤呼吸的机制不同, 其影响大小不同。当土壤体积含水量变化范围很小, 其对土壤呼吸的影响有可能被其他因子所掩盖, 一般很难显现出来, 只有土壤体积含水量超出极端范围, 影响到了微生物与植物根系活动的时候, 对土壤呼吸的影响才凸显出来。
然而, 城市绿地土壤的水热条件往往相互作用、相互影响, 难以区分其各自对城市绿地土壤呼吸过程的作用。本研究结果显示, 综合了T10和VWC10的双因子复合模型对土壤呼吸速率的变化解释能力明显强于单因子模型, 解释能力均超过50%, 这与其它研究结果类似[33, 39]。杨金艳等[33]发现, 复合模型的解释能力甚至达到67.5%-90.6%。但也有研究表明, 复合模型并没有提高对土壤呼吸速率变化的解释能力[40], 这可能是因为不同研究区域的土壤体积含水量的范围不同。Kucera和kirkham[41]指出, 土壤体积含水量只有达到土壤微生物永久性萎蔫点或者超过田间持水量的情况下, 土壤体积含水量才对土壤呼吸的影响起主导作用, 而在一般情况下, 土壤体积含水量对土壤呼吸的影响不大。总的来说, 城市绿地的土壤呼吸速率在大的时间尺度上(一年以上)主要受土壤温度的影响;但在小的时间尺度上, 城市绿地土壤体积含水量的每次峰值变化都会引起土壤呼吸作用的变化, 本研究中6、8月Rs的降低可能正是由于监测取样前一天的降雨造成的。
4 结论(1)海珠湖公园城市绿地的Rs变化受季节变化的影响较大。在湿季, Rs较大且变化幅度较大, 疏林、灌丛和草地的Rs变化范围分别为(3.53±0.46)-(13.81±1.31)μmol m-2 s-1、(2.82±0.22)-(12.72±1.16)μmol m-2 s-1和(2.80±0.30)-(9.83±0.96)μmol m-2 s-1;在干季, Rs较小且变化幅度较小, 疏林、灌丛和草地的Rs变化范围分别为(1.66±0.18)-(3.26±0.20)μmol m-2 s-1、(1.27±0.15)-(3.67±0.16)μmol m-2 s-1和(1.94±0.08)-(6.82±1.13)μmol m-2 s-1。
(2)T10和VWC10均对土壤呼吸过程有重要的影响。单因子指数模型可以很好的拟合T10与Rs的关系, T10对Rs变化的解释能力在40%左右;VWC10对Rs变化的解释能力在10%-24%左右;T10和VWC10往往相互作用、相互影响, 共同作用于土壤呼吸过程, 双因素(T10和VWC10)复合模型的解释能力较单因子模型有明显提高, 均在50%以上, 复合模型为Rs=α·exp (βT10+γVWC10)。
(3)干湿季Q10差异显著, 疏林、灌丛和草地湿季的Q10比干季的分别高0.44、0.70和0.46。
(4)本研究重点考虑了T10和VWC10对城市绿地土壤呼吸速率季节动态变化的影响, 但城市绿地土壤呼吸过程的影响因素相当复杂。比如土壤有机质含量、土壤微气候、植被根系分泌物和植被生物量分配等均可能对土壤呼吸过程有重要影响, 未来有必要综合考虑相关因素的影响。
| [1] | Harper C W, Blair J M, Fay P A, Knapp A K, Carlisle J D. Increased rainfall variability and reduced rainfall amount decreases soil CO2 flux in a grassland ecosystem. Global Change Biology , 2005, 11 (2) : 322–334. DOI:10.1111/gcb.2005.11.issue-2 |
| [2] | Singh J S, Gupta S R. Plant decomposition and soil respiration in terrestrial ecosystems. The Botanical Review , 1977, 43 (4) : 449–528. DOI:10.1007/BF02860844 |
| [3] | Buchmann N. Biotic and abiotic factors controlling soil respiration rates in Picea abies stands. Soil Biology and Biochemistry , 2000, 32 (11-12) : 1625–1635. DOI:10.1016/S0038-0717(00)00077-8 |
| [4] | Schimel D S, House J I, Hibbard K A, Bousquet P, Ciais P, Peylin P, Braswell B H, Apps M J, Baker D, Bondeau A, Canadell J, Churkina G, Cramer W, Denning A S, Field C B, Friedlingstein P, Goodale C, Heimann M, Houghton R A, Melillo J M, Moore Ⅲ B, Murdiyarso D, Noble I, Pacala S W, Prentice I C, Raupach M R, Rayner P J, Scholes R J, Steffen W L, Wirth C. Recent patterns and mechanisms of carbon exchange by terrestrial ecosystems. Nature , 2001, 414 (6860) : 169–172. DOI:10.1038/35102500 |
| [5] | Schlesinger W H, Andrews J A. Soil respiration and the global carbon cycle. Biogeochemistry , 2000, 48 (1) : 7–20. DOI:10.1023/A:1006247623877 |
| [6] | Seto K C, Güneralp B, Hutyra L R. Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. Proceedings of the National Academy of Sciences of the United Stated of America , 2012, 109 (40) : 16083–16088. DOI:10.1073/pnas.1211658109 |
| [7] | 朱旺生.城市绿地系统树种规划研究[D].南京:南京林业大学, 2011. http://cdmd.cnki.com.cn/Article/CDMD-10298-1011401333.htm |
| [8] | 孙倩, 方海兰, 梁晶, 钱秀苇, 刘鸣达, 张庆费, 郝瑞军, 郝冠军. 上海典型城市草坪土壤呼吸特征. 生态学杂志 , 2009, 28 (8) : 1572–1578. |
| [9] | 张东秋, 石培礼, 张宪洲. 土壤呼吸主要影响因素的研究进展. 地球科学进展 , 2005, 20 (7) : 778–785. |
| [10] | 杨玉盛, 董彬, 谢锦升, 陈光水, 高人, 李灵, 王小国, 郭剑芬. 森林土壤呼吸及其对全球变化的响应. 生态学报 , 2004, 24 (3) : 583–591. |
| [11] | 张鸽香, 徐娇, 王国兵, 武珊珊, 阮宏华. 南京城市公园绿地不同植被类型土壤呼吸的变化. 生态学杂志 , 2010, 29 (2) : 274–280. |
| [12] | 李丹丹.沈阳城市绿地土壤呼吸特征及其碳汇分析[D].沈阳:辽宁大学, 2012. http://cdmd.cnki.com.cn/Article/CDMD-10140-1013104100.htm |
| [13] | 陈骥, 曹军骥, 魏永林, 刘吉宏, 马扶林, 陈迪超, 冯嘉裕, 夏瑶, 岑燕. 青海湖北岸高寒草甸草原非生长季土壤呼吸对温度和湿度的响应. 草业学报 , 2014, 23 (6) : 78–86. |
| [14] | 王风玉, 周广胜, 贾丙瑞, 王玉辉. 水热因子对退化草原羊草恢复演替群落土壤呼吸的影响. 植物生态学报 , 2003, 27 (5) : 644–649. |
| [15] | Bowden R D, Newkirk K M, Rullo G M. Carbon dioxide and methane fluxes by a forest soil under laboratory-controlled moisture and temperature conditions. Soil Biology and Biochemistry , 1998, 30 (12) : 1591–1597. DOI:10.1016/S0038-0717(97)00228-9 |
| [16] | Carlyle J C, Than U B. Abiotic controls of soil respiration beneath an eighteen-year-old Pinus radiata stand in south-eastern Australia. Journal of Ecology , 1988, 76 (3) : 654–662. DOI:10.2307/2260565 |
| [17] | 周志翔, 邵天一, 唐万鹏, 王鹏程, 刘学全, 徐永荣. 城市绿地空间格局及其环境效应--以宜昌市中心城区为例. 生态学报 , 2004, 24 (2) : 186–192. |
| [18] | Freibauer A, Rounsevell M D A, Smith P, Verhagen J. Carbon sequestration in the agricultural soils of Europe. Geoderma , 2004, 122 (1) : 1–23. DOI:10.1016/j.geoderma.2004.01.021 |
| [19] | Mosier A, Schimel D, Valentine D, Bronson K, Parton W. Methane and nitrous oxide fluxes in native, fertilized and cultivated grasslands. Nature , 1991, 350 (6316) : 330–332. DOI:10.1038/350330a0 |
| [20] | 裴淑玮.华北农田施肥对温室气体及含氮气体排放影响[D].北京:北京林业大学, 2012. http://cdmd.cnki.com.cn/Article/CDMD-10022-1012350062.htm |
| [21] | 展小云, 于贵瑞, 郑泽梅, 王秋凤. 中国区域陆地生态系统土壤呼吸碳排放及其空间格局--基于通量观测的地学统计评估. 地理科学进展 , 2012, 31 (1) : 97–108. |
| [22] | 李霞, 孙睿, 李远, 王修信, 谢东辉, 严晓丹, 朱启疆. 北京海淀公园绿地二氧化碳通量. 生态学报 , 2010, 30 (24) : 6715–6725. |
| [23] | 王旭, 闫瑞瑞, 邓钰, 闫玉春, 辛晓平. 放牧对呼伦贝尔草甸草原土壤呼吸温度敏感性的影响. 环境科学 , 2014, 35 (5) : 1909–1914. |
| [24] | 朱楠, 王彬, 王云琦, 孙素琪, 吴超, 刘李紫渊. 重庆缙云山4种典型林分生长季土壤呼吸特征及其与环境因子的关系. 中国水土保持科学 , 2014, 12 (2) : 16–23. |
| [25] | Zhang Q, Lei H M, Yang D W. Seasonal variations in soil respiration, heterotrophic respiration and autotrophic respiration of a wheat and maize rotation cropland in the North China Plain. Agricultural and Forest Meteorology , 2013, 180 : 34–43. DOI:10.1016/j.agrformet.2013.04.028 |
| [26] | 李涛, 李芹, 王树明, 李春, 赵东兴, 张勇, 高梅. 云南河口不同林龄人工橡胶林土壤CO2浓度的变化规律及其影响因素. 热带作物学报 , 2015, 36 (1) : 9–15. |
| [27] | 颜廷武, 尤文忠, 张慧东, 魏文俊, 王睿照, 毛沂新. 辽东山区落叶松人工林非生长季土壤呼吸及其影响因子研究. 辽宁林业科技 , 2015 (1) : 20–22. |
| [28] | 付芝红, 呼延佼奇, 李锋, 宋英石, 赵丹, 李慧. 城市不同地表覆盖类型对土壤呼吸的影响. 生态学报 , 2013, 33 (18) : 5500–5508. |
| [29] | Epron D, Farque L, Lucot E, Badot P M. Soil CO2 efflux in a beech forest:the contribution of root respiration. Annals of Forest Science , 1999, 56 (4) : 289–295. DOI:10.1051/forest:19990403 |
| [30] | 孟春, 王立海, 沈微. 针阔混交林择伐作业后土壤呼吸与土壤温度和湿度的关系. 东北林业大学学报 , 2008, 36 (5) : 33–35. |
| [31] | Luo Y Q, Zhou X H. Soil Respiration and the Environment. New York:Academic Press , 2006 : 257–305. |
| [32] | 谭炯锐, 查同刚, 张志强, 孙阁, 戴伟, 方显瑞, 徐枫. 土壤温湿度对北京大兴杨树人工林土壤呼吸的影响. 生态环境学报 , 2009, 18 (6) : 2308–2315. |
| [33] | 杨金艳, 王传宽. 土壤水热条件对东北森林土壤表面CO2通量的影响. 植物生态学报 , 2006, 30 (2) : 286–294. |
| [34] | Reichstein M, Rey A, Freibauer A, Tenhunen J, Valentini R, Banza J, Casals P, Cheng Y F, Grünzweig J M, Irvine J, Joffre R, Law B E, Loustau D, Miglietta F, Oechel W, Ourcival J M, Pereira J S, Peressotti A, Ponti F, Qi Y, Rambal S, Rayment M, Romanya J, Rossi F, Tedeschi V, Tirone G, Xu M, Yakir D. Modeling temporal and large-scale spatial variability of soil respiration from soil water availability, temperature and vegetation productivity indices. Global Biogeochemical Cycles , 2003, 17 (4) . DOI:10.1029/2003GB002035 |
| [35] | Davidson E A, Verchot L V, Cattânio J H, Ackerman I L, Carvalho J E M. Effects of soil water content on soil respiration in forests and cattle pastures of eastern Amazonia. Biogeochemistry , 2000, 48 (1) : 53–69. DOI:10.1023/A:1006204113917 |
| [36] | Ilstedt U, Nordgren A, Malmer A. Optimum soil water for soil respiration before and after amendment with glucose in humid tropical acrisols and a boreal mor layer. Soil Biology and Biochemistry , 2000, 32 (11-12) : 1591–1599. DOI:10.1016/S0038-0717(00)00073-0 |
| [37] | Stark J M, Firestone M K. Mechanisms for soil moisture effects on activity of nitrifying bacteria. Applied and Environmental Microbiology , 1995, 61 (1) : 218–221. |
| [38] | 陈全胜, 李凌浩, 韩兴国, 阎志丹. 水分对土壤呼吸的影响及机理. 生态学报 , 2003, 23 (5) : 972–978. |
| [39] | 于雷, 王玉杰, 王云琦, 张会兰, 孙素琪, 周彬. 重庆缙云山针阔混交林地土壤呼吸速率及温度敏感性特征分析. 环境科学研究 , 2014, 27 (1) : 57–63. |
| [40] | Chen Q S, Wang Q B, Han X G, Wan S Q, Li L H. Temporal and spatial variability and controls of soil respiration in a temperate steppe in northern China. Global Biogeochemical Cycles , 2010, 24 (2) . DOI:10.1029/2009GB003538 |
| [41] | Kucera C L, Kirkham D R. Soil respiration studies in tallgrass prairie in Missouri. Ecology , 1971, 52 (5) : 912–915. DOI:10.2307/1936043 |