 2016, Vol. 36
2016, Vol. 36文章信息
- 邱赛, 邢艳秋, 徐卫华, 丁建华, 田静
- QIU Sai, XING Yanqiu, XU Weihua, DING Jianhua, TIAN Jing.
- 星载大光斑LiDAR与HJ-1A高光谱数据联合估测区域森林地上生物量
- Estimation of regional forest aboveground biomass combining spaceborne large footprint LiDAR and HJ-1A Hyperspectral Images
- 生态学报[J]. 2016, 36(22): 7401-7411
- Acta Ecologica Sinica[J]. 2016, 36(22): 7401-7411
- http://dx.doi.org/10.5846/stxb201601050027
-
文章历史
- 收稿日期: 2016-01-05
- 修订日期: 2016-01-05





2. 中国科学院生态环境研究中心, 北京 100085
2. Research center for eco-environmental sciences, Chinese academy of sciences, Beijing 100085, China
森林作为陆地生态系统的主体, 在维持碳平衡及保持物种多样性方面发挥着巨大作用, 其在改善环境方面的巨大优势已引起广泛关注[1-3]。森林生物量已被列为监测森林生态系统碳储量的重要指标之一[4]。森林地上生物量包括干、枝和叶的生物量, 是评估森林生态系统碳储量的基础, 如何快速准确的获取森林地上生物量及其分布对研究森林碳循环和估测碳储量具有重要意义[5-7]。
传统上森林地上生物量的估测需要依赖于大量的野外实际调查数据, 不仅费时费力, 还难以实现大区域森林地上生物量的估测[8-10]。随着遥感技术的出现和发展, 光学遥感和微波遥感最早应用到了森林结构参数的估测[5], 但是由于天气条件及信号饱和等问题的影响, 导致光学遥感和微波遥感在森林地上生物量估测方面受到限制[11-13]。作为新型的主动遥感技术, 激光雷达在估测森林结构参数和生物量方面具有明显的优势[14-16]。
ICESat-GLAS (the ice, cloud, and land elevation satellite, ICESat, geoscience laser altimeter system, GLAS)是目前唯一的星载大光斑LiDAR设备, 其激光脉冲不仅能够到达冠顶, 还能够到达地面, 并且能够提供完整的波形数据, 在森林结构参数和森林地上生物量估测中得到广泛应用[16-22]。但是, 由于ICESat-GLAS波形数据成离散条带状分布, 在区域尺度森林地上生物量估测方面受到限制。为了实现区域尺度森林地上生物量的估测, 需联合其他光学遥感数据。相比于多光谱数据, 高光谱遥感数据能同时获取数十至上百个波段的连续光谱图像信息, 具有较高的光谱分辨率, 同时包含了地物更加丰富的空间、辐射和光谱三重信息[23], 在森林制图、森林资源调查等方面具有优势。因此本文尝试采用HJ-1A高光谱数据与GLAS数据联合反演森林地上生物量。
为了提高ICESat-GLAS波形数据估测森林地上生物量的精度, 本研究同时引入森林最大树高和森林郁闭度估测森林地上生物量, 借助HJ-1A高光谱数据的连续性弥补ICESat-GLAS在区域森林地上生物量估测方便的不足。本研究从GLAS波形中提取地形坡度参数TS, 消除地形坡度对估测森林最大树高造成的影响, 提高GLAS估测森林最大树高的精度;通过设定不同的林下植被高度阈值, 降低林下植被对森林郁闭度估测的影响, 提高GLAS森林郁闭度估测精度;最后利用GLAS估测的森林最大树高和森林郁闭度共同建立森林地上生物量模型, 并联合HJ-1A高光谱数据基于支持向量回归机算法实现区域推广, 得到森林地上生物量分布图。
1 研究区和研究数据 1.1 研究区概况本研究选择吉林省汪清林业局经营区(43°05′N-43°40′N, 129°56′E-131°04′E)为研究区(图 1)。该研究区平均海拔高度为806m, 地面高程在360m-1477m范围内, 坡度在0°-45°范围内。该研究区内植物种类繁多, 植被结构复杂, 深山区林相以针阔混交林为主, 呈带状分布在海拔500-1100m内。针叶树以红松(Pinus koraiensisSiebold & Zucc.)、云杉(Picea asperata Mast.)和臭松(Abies nephrolepis (Trautv. ex Maxim.) Maxim.)为主;阔叶树以紫椴(Tilia amurensisRupr. var. amurensis)、蒙古栎(Quercus mongolicaFisch. ex Ledeb.)、枫桦(Betula costata Trautv.)、色木槭(Acer monoMaxim.)和白桦(Betula platyphylla Suk.)为主。

|
| 图 1 研究区及野外调查样地位置 Fig. 1 The study area and the field sampling point position 圆形代表野外采样点 |
GLAS数据共包含15种数据产品(GLA01, GLA02, GLA03, …, GLA15), 本研究区获取了L3E (2006年3月), L3F (2006年6月), L3G (2006年11月), L3H (2007年3月)和L3I (2007年10月)共5期GLA01和GLA14数据产品, 数据获取网址为http://nsidc.org/data/icesat/。GLA01产品中包含了完整的波形数据, 对于陆地而言, 每个波形数据包含544帧数据, 每一帧对应真实距离0.15m。GLA14产品中包含地面高程信息及经纬度等信息, 两者之间使用头文件中的索引号和光斑号进行匹配[24]。由于GLAS波形数据易受到云层和系统噪声的影响, 因此首先通过GLA14产品中记录的无饱和指数(i_satNdx=0)和无云指数(i_FRir_qaFlg=15)选择出受云层和系统噪声影响小的高质量波形数据, 最终得到1363组有效的GLAS波形数据。
1.2.2 HJ-1A/HSI高光谱数据环境减灾小卫星星座A/B星上搭载的超光谱成像仪(Hyperspectral Imaging Radiometer, HSI), 是世界上第一个基于空间调制干涉成像原理设计的民用高光谱成像仪[25]。HSI共包含115个窄波段信息, 光谱范围为0.45-0.95μm, 空间分辨率为100m, 平均光谱分辨率为4.32nm。幅宽为50km, 重访周期为96h, 且具备±30°的侧视能力。本研究共获取了4景HSI高光谱遥感影像, 数据获取网址为http://www.cresda.com/n16/index.html, 由于受云层及多景拼接的影响, HSI数据获取时间有差异, 其中2景获取时间为2010年6月5日, 另外2景获取时间为2010年6月9日。
1.2.3 地面数据本研究分别于2006年9月和2007年9月, 沿着ICESat-GLAS轨道方向随机选取183个GLAS光斑建立野外数据调查样地, 每个样地的水平投影面积为500m2。通过激光测高仪测量样地内每株树的树高, 用坡度仪测量样地内的坡度值, 使用配有三星NV3型号相机的180°鱼眼镜头测量样地内的郁闭度, 为了消除灌木杂草的影响, 鱼眼镜头的离地高度设为1.5m, 另外为了保证实测郁闭度的准确度及代表性, 对样地周围50m范围内郁闭度差异较大的地区, 亦用鱼眼镜头进行了测量, 最后与样地内的郁闭度进行平均作为此样地的森林郁闭度。针对吉林省汪清地区实际情况, Deo[26]采用单木二次采样的方式在样地内进行数据收集, 建立了单木地上生物量模型, 模型精度较好, 较稳定, 并且对森林类型及树种不敏感, 能够满足林业上的要求, 模型公式如公式(1)所示:

|
(1) |
式中, B为单木地上单木生物量(t/hm2), D为胸径(cm)。
本研究采用此模型计算样方内的单木生物量, 随后将单木地上生物量求和得到样方内的总地上生物量, 最后将总地上生物量除以样方面积得到单位面积上的森林地上生物量。在统计建模中建模样方数多数是验证样方数的两倍, 因此本研究中将数据按近似2:1的比例随机分成两组, 一组包含122组数据用于模型建模, 另一组包含61组数据用于模型验证。
2 研究方法 2.1 ICESat-GLAS波形数据预处理利用IDL_Reader程序包将二进制格式的GLA01和GLA14数据产品转换成由电压(单位伏特(v))表示的数据。由于获取的数据时期不同, 为了统一进行分析计算, 本研究将数据进行标准化, 即每一时刻的接收能量除以回波总能量。对标准化后的数据进行滤波、高斯分解拟合。通过设定背景噪声阈值确定波形起波点与止波点位置, 背景噪声阈值的设定参考以往研究, 采用波形前100帧数据的平均值与其4倍的标准差之和[27], 如公式(2)所示:
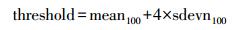
|
(2) |
式中, threshold为背景噪声阈值, mean100为前100帧数据的平均值, sdevn100为前100帧数据的标准差。
2.2 HJ-1A/HSI高光谱数据处理HSI数据是2级产品, 已经进行了几何校正和辐射定标, 本研究在此基础上利用地形图对两种数据进行了几何精校正, 然后利用FLAASH大气校正模块对HSI高光谱数据进行大气校正。由于GLAS光斑直径为70m, HJ-1A/HSI数据的分辨率为100m, 为了数据匹配, 首先利用双线性内插法将HJ-1A/HSI的分辨率调整为70m。HSI数据包含115个波段, 数据冗余大, 并且相邻波段间相关性高, 因此本研究采用最小噪声分离法对大气校正后的高光谱数据降维, 将尽可能多的数据集中在少量的几个MNF分量中, 并减少相关性。本研究中前3个MNF分量的信息占总信息的98%, 所以本研究采用前3个MNF分量进行分析。将GLAS光斑中心点坐标加入到MNF分量图中, 由于GLAS光斑中心点不一定落在像元的中心, 所以有些GLAS光斑可能覆盖至少两个像元, 因此本研究中将GLAS光斑覆盖所有像元的平均值作为光斑对应的MNF值[28]。
2.3 ICESat-GLAS估测森林最大树高一般情况下, 在平坦地区, 波形长度即为森林最大树高, 但在林地中, 受坡度、光斑直径和地形起伏变化的影响, 波形展宽现象明显[29], 如图 2所示, 当地形坡度为TS时, 波形的最大展宽即最大的估测误差为Dtan (TS), 其中D为GLAS光斑直径。本研究尝试从GLAS波形数据中提取地形坡度参数TS来减弱地形坡度对森林最大树高估测造成的误差。地形坡度参数TS参照Mahoney等[30]提出的方法进行计算, 具体计算公式如公式(3)、(4)、(5)所示, 波形长度W为波形起波点(SigStart)与止波点(SigEnd)之间的垂直距离(如图 3所示), 其计算公式如公式(6)所示, 图 3中展示的是GLAS波形及波形参数。拟利用波形长度和TS建立的森林最大树高模型如公式(7)所示。

|
| 图 2 地形坡度对GLAS波形估测森林冠层高度的影响 Fig. 2 Influence of terrain slope on GLAS waveform estimating forest canopy height |

|
| 图 3 ICESat-GLAS波形数据及波形参数 Fig. 3 ICESat-GLAS waveform and parameters |
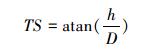
|
(3) |
式中, TS为地形坡度参数(m), h为光斑内地面最高点与地面最低点之间的垂直距离(m), D为光斑直径(m)。
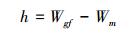
|
(4) |
式中, h为光斑内地面最高点与地面最低点之间的垂直距离(m), Wgf为地面回波对应的高斯分量的波形长度(m), Wm为由于大气衰减和信号饱和等导致的地面回波波形展宽(m)。

|
(5) |
式中, A为地面回波峰值, Wm为由于大气衰减和信号饱和等导致的地面回波波形展宽(m)。
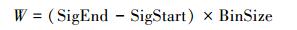
|
(6) |
式中, SigStart为波形起波点, SigEnd为波形止波点, BinSize为每帧数据代表的真实距离, 对陆地而言, 每一帧代表真实距离0.15m, 因此BinSize取值为0.15m。
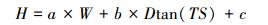
|
(7) |
式中, H为森林最大树高(m), W为波形长度(m), D为光斑直径(m), TS为地形坡度参数(°), a, b, c为相应系数。
2.4 ICESat-GLAS估测森林郁闭度森林郁闭度是指森林中乔木树冠遮蔽地面的程度, 是森林蓄积量及森林生物量估测的重要影响因素之一[31-33]。ICESat-GLAS发射的激光脉冲穿过大气层后首先会到达森林冠顶, 其中部分被反射回传感器, 部分会继续向下传播, 当再遇到冠层时会继续被反射, 而部分激光脉冲会最终透过叶片缝隙到达地面, 并被地面反射。由此可以假设, 森林郁闭度越大, 植被冠层反射回的能量越多, 相应地面反射回的能量将减少;同理, 森林郁闭度越小, 植被冠层反射回的能量将减少, 被地面反射回的能量将增多。本研究从ICESat-GLAS波形数据中计算乔木层植被回波能量和回波总能量, 并将乔木层植被回波能量与回波总能量的比值用于估测森林郁闭度。如公式(8)所示:
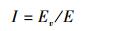
|
(8) |
式中, Ev为乔木层植被回波能量, E为回波总能量, I为森林郁闭度。
本研究的关键是确定乔木层植被回波与林下植被回波的分界线, 由以上分析可知, 波形回波能量会受到林下植被的影响, 结合野外样地调查分析, 林下植被高度在1-3m左右, 因此本研究将林下植被高度分为1m、2m和3m分别进行研究。本研究将乔木层植被与林下植被分界线Lvg定义为地面回波峰值位置减去林下植被高度的位置, 如图 3所示。乔木层植被回波能量Ev为起波点(SigStart)与Lvg之间波形能量和。回波总能量E为起波点(SigStart)与止波点(SigEnd)之间波形能量和。
2.5 ICESat-GLAS估测森林地上生物量一般在假设其他因素相同的情况下, 树高越高, 林木生物量越高, 相应的森林地上生物量越高;森林郁闭度越大, 森林越茂盛, 进而森林地上生物量越高。如图 4(a)(b)所示, 森林最大树高和森林郁闭度与森林地上生物量具有线性关系, adj.R2分别为0.53和0.44, 所以本研究联合森林最大树高和森林郁闭度共同建立森林地上生物量模型。由于森林最大树高与森林郁闭度的单位及数据范围不一致, 因此本研究先对数据进行归一化, 归一化公式如公式(9)所示, 联合森林最大树高和森林郁闭度建立的森林地上生物量公式如公式(10)所示:

|
| 图 4 森林最大树高和森林郁闭度与森林地上生物量的关系 Fig. 4 The relationship between aboveground forest biomass and maximum forest height and forest canopy density |
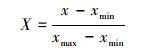
|
(9) |
式中, X为归一化后的值, x为归一化前的值, xmin为最小值, xmax为最大值。
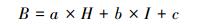
|
(10) |
式中, B为森林地上生物量(t/hm2), H为归一化森林最大树高, I为归一化森林郁闭度, a, b, c为相应的系数。
2.6 基于支持向量回归机算法区域森林地上生物量制图GLAS数据呈离散条带状分布, 无法实现区域估测, 而高光谱数据能同时获取数十至上百个波段的连续光谱图像信息, 因此本研究将此二种数据联合, 实现GLAS对森林地上生物量的区域估测。支持向量机是在统计学理论上发展起来的, 一种针对有限样本情况, 实现结构风险最小化思想的机器学习方法, 相比于神经网络等其他机器学习算法, 支持向量回归机算法具有更强的逼近能力及泛化能力, 避免了神经网络出现的局部最小问题。本研究采用的是LIBSVM工具包实现的, 具体算法如下:
(1)通过公式(10)计算研究区内所有GLAS光斑内的森林地上生物量值, 并将其与相应的3个MNF分量构成的样本数据加入LIBSVM中, 按2:1的比例随机分为训练集和测试集。
(2)选择核函数, 由于径向基核函数(如公式(11)所示)在支持向量回归机中应用广泛, 因此本研究亦采用径向基核函数。
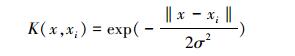
|
(11) |
式中, σ为函数的宽度参数, 用于控制函数的径向作用范围, xi为核函数中心, x为因变量。
(3)设定参数C和σ的寻优范围, 种群最大数量, 种群交叉概率和交叉验证次数, 采用遗传算法对LIBSVM程序包中的惩罚参数C和核函数参数σ两个参数进行寻优。C值越大, 训练数据建模越复杂, 会出现过学习问题;C值越小, 建模越简单, 会出现欠学习问题。σ值影响映射到高维特征空间中样本数据的复杂程度, 其值越大越复杂, 寻优效率也会降低, 一般将参数C和σ寻优范围设为[0.01, 1000]之间。种群最大数量会影响遗传算法的收敛性和寻优效率, 其值越大, 寻优效果越好, 但是寻优效率会降低, 通常将范围设定在[20, 200]之间, 本文设定种群最大数量为200。种群交叉频率越大, 寻优效率也会降低, 一般将取值范围设在[0.2, 0.9]之间, 本文中将种群交叉概率0.4。交叉验证次数是确定将样本集分为多少个较小的子集对寻优结果进行验证, 其值的设定需要考虑样本数量和寻优效率, 一般设定范围为[2, 6], 本文中将交叉验证次数设定为4。
(4)利用寻优得到的参数及训练集数据建模, 并用测试集数据对所建模型的泛化能力进行测试。
(5)运用此模型计算无GLAS光斑覆盖区域的森林地上生物量, 最终得到研究区所有区域的森林地上生物量, 即生成森林地上生物量图。
3 结果与分析 3.1 ICESat-GLAS估测森林最大树高模型图 5所示为地形坡度参数TS与地形坡度间的散点图, 可以看出地形坡度参数TS与地形坡度间存在明显的线性关系(adj.R2=0.78)。因此本研究利用TS和波形长度W建立森林最大树高模型, 所建森林最大树高模型的adj.R2=0.78, RMSE=2.51m (建模数据为122组), 模型验证的adj.R2=0.85, RMSE=1.67m (模型验证数据为61组), 图 6为模型验证散点图, 模型公式如公式(12)所示。

|
| 图 5 地形坡度参数TS与地形坡度的散点图 Fig. 5 The scatter plot between TS and terrain slope |

|
| 图 6 森林最大树高模型验证结果 Fig. 6 The validation result of maximum forest height model |
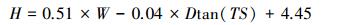
|
(12) |
式中, H为森林最大树高(m), W为波形长度(m), D为光斑直径(m), TS为地形坡度参数/°
分析图 6发现, 实测值与估测值之间存在偏差, 本文对偏差较大样地所对应的GLAS数据及野外数据进行分析, 发现这些样地植被覆盖度较高, 林下植被丰富, 且地形较复杂, 由于枝叶的遮挡, 会导致野外实测树高出现测量偏差, 此外林下植被对GLAS波形也会造成一定的影响, 进而影响森林最大树高的估测。
3.2 ICESat-GLAS估测森林郁闭度模型表 1为设定不同林下植被高度得到的森林郁闭度建模结果。分析表 1发现, 将林下植被高度设定为2m时模型的效果是最好的, 所建模型的adj.R2=0.64, RMSE=0.13(建模数据为122组)。因此本研究采用林下植被高度2m进行后续研究, 模型验证的adj.R2=0.65, RMSE=0.12(模型验证数据为61组), 模型公式如公式(13)所示。图 7为参数I与野外实测森林郁闭度间的散点图及拟合直线, 图 8为模型的验证结果。

|
| 图 7 参数I与野外实测森林郁闭度间的散点图 Fig. 7 The scatter plot between parameterI and observed forest canopy density |

|
| 图 8 森林郁闭度模型的验证结果 Fig. 8 The verification result of the linear forest canopy density model |
| 林下植被高度Under vegetation height/m | adj.R2 | RMSE |
| 1 | 0.52 | 0.21 |
| 2 | 0.64 | 0.13 |
| 3 | 0.60 | 0.20 |
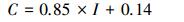
|
(13) |
式中, C为实测森林郁闭度, I为GLAS中乔木层植被回波能量与回波总能量之比。
分析图 7和图 8发现, 尽管设定了林下植被高度值, 尽量消除林下植被对乔木层植被回波的影响, 但是森林郁闭度仍存在被低估或高估的现象。结合野外实地调查数据发现, 这些样地的地形条件较复杂, 地形坡度或地形起伏较大, 可见地形条件也是影响森林郁闭度估测准确度的因素之一。研究区内以中龄林为主, 存在部分幼龄林, 其郁闭度会有所差异, 导致估测结果出现误差。研究区内树木种类繁多, 不同树种其生长状况有差异, 郁闭度也会不同。
3.3 ICESat-GLAS估测森林地上生物量模型本研究同时考虑森林垂直结构参数和森林水平结构参数, 基于支持向量回归机算法, 利用ICESat-GLAS估算的森林最大树高和森林郁闭度用于森林地上生物量建模。表 2所示为单独利用森林最大树高和同时利用森林最大树高与森林郁闭度的建模结果。从表 2中可以看出, 只利用GLAS估测的森林最大树高建立的森林地上生物量模型的adj.R2为0.50, RMSE为12.36 t/hm2, 引入森林郁闭度后, 模型的精度有所提高, adj.R2由0.50增加到0.62, RMSE减小到10.88 t/hm2, 说明森林郁闭度对提高森林地上生物量估测精度有一定的帮助。利用森林最大树高和森林郁闭度建立的森林地上生物量模型公式如公式(14)所示。
| 参数Parameter | adj.R2 | RMSE/(t/hm2) |
| H | 0.50 | 12.36 |
| H, C | 0.62 | 10.88 |
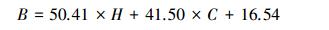
|
(14) |
式中, B为实测森林地上生物量(t/hm2), H为GLAS归一化森林最大树高, C为GLAS归一化森林郁闭度。
图 9为利用森林最大树高和森林郁闭度建立的森林地上生物量模型验证结果, 模型验证的adj.R2=0.60, RMSE=11.52 t/hm2。利用公式(14)计算研究区内所有GLAS光斑的森林地上生物量, 用于整个研究区森林地上生物量的反演。

|
| 图 9 森林地上生物量模型验证结果 Fig. 9 The verification result of the forest aboveground biomass model |
本研究将GLAS光斑数据与HJ-1A/HSI高光谱数据联合, 基于支持向量回归机算法实现了GLAS数据对区域森林地上生物量的估测。GLAS估测的森林地上生物量与前3个MNF分量(MNF1, MNF2, MNF3)间的SVR模型建模的adj.R2=0.73, RMSE=16.28 t/hm2, 模型验证的adj.R2=0.70, RMSE=21.42 t/hm2, 模型最优参数C=4.9902, σ=1.9298, 图 10所示为最终得到的森林地上生物量分布图。

|
| 图 10 森林地上生物量分布图 Fig. 10 The map of aboveground forest biomass |
分析图 10发现, 整个研究区的森林地上生物量在0-164 t/hm2范围内, 从图 10中可以发现研究区的北部和西南部森林地上生物量值较高, 而南部和东北部地区生物量值较低, 森林地上生物量为零的这些区域分布的主要是居民区、道路和河流。本研究利用所有野外数据对得到的森林地上生物量分布图进行验证, 验证结果如图 11所示, 估测森林地上生物量与野外实测森林地上生物量间存在明显线性关系(adj.R2=0.62, RMSE=11.11 t/hm2)。尽管引入森林郁闭度, 森林地上生物量模型的精度有所提高, 但是森林地上生物量的估测值与实测值间仍然存在差异。导致结果存在差异的原因很多, 对数据进行统计分析发现, 森林地上生物量随海拔先增加后减少(图 12), 森林地上生物量主要集中在400-800m之间, 主要是因为随着海拔的增高, 气温在降低, 在海拔1000m以上气温平均在3℃左右, 不利于树木的生长, 会出现很多的“小老树”[13]。因此, 海拔高度亦是影响森林地上生物量估测准确度的一个因素, 如何降低海拔的影响仍然需要进一步的研究。

|
| 图 11 实测森林地上生物量与估测森林地上生物量对比 Fig. 11 Comparison of AGB estimated by GLAS and HJ-1A/HSI data and observed AGB |

|
| 图 12 森林地上生物量随海拔高度的变化 Fig. 12 The change of forest above ground biomass with different altitude |
ICESat-GLAS可以实时快速准确的获取区域乃至全国的森林结构信息, 尤其是垂直结构信息, 为了提高ICESat-GLAS数据估测森林地上生物量的精度和弥补ICESat-GLAS数据估测区域森林地上生物量的不足, 本研究将ICESat-GLAS波形数据与HJ-1A/HSI高光谱影像联合, 基于支持向量回归机算法, 建立GLAS估测森林地上生物量与HJ-1A高光谱影像的3个MNF分量间的SVR模型, 提高了区域森林地上生物量的估测精度, 生成了整个研究区的森林地上生物量分布图, 方便对区域森林地上生物量的直观观测。
提取的地形坡度参数TS跟实测地形坡度的相关性较高, 利用从GLAS数据中提取的地形坡度参数TS, 降低地形对GLAS波形数据展宽造成的影响, 避免了以往借助DEM数据, 由于DEM数据分辨率造成的误差。相比于Xing等[15]利用波形长度和地形指数(从DEM提取)建立的森林最大树高模型, 模型精度得到提高。探究了林下植被对GLAS波形估测森林郁闭度的影响, 通过设定不同的林下植被高度值(1, 2, 3m)进行分析以降低林下植被的影响, 结果表明将林下植被高度设为2m时, 模型的精度最高。
森林郁闭度对森林地上生物量的影响不容忽视, 相比于以往只利用单一森林参数(比如树高、胸径等)建立的森林地上生物量模型, 同时引入森林最大树高和森林郁闭度估测森林地上生物量, 即同时考虑了森林垂直结构参数和森林水平结构参数对森林地上生物量的影响。相比于只利用森林最大树高建立的森林地上生物量模型, 模型精度有所提高, 建立的多元线性回归模型精度较高, 联合GLAS数据和HJ-1A/HSI数据, 基于支持向量回归机算法建立了森林地上生物量SVR模型, 通过SVR模型生成了森林地上生物量分布图, 且森林地上生物量的估测值与野外实测值具有很好的线性关系(adj.R2=0.62, RMSE=11.11 t/hm2), 对未来进一步研究区域森林地上生物量, 提高森林地上生物量和碳储量的估测精度有一定的参考价值。相比于多元统计方法, 在小样本情况下, 支持向量回归机算法具有更强的泛化能力。
研究采用的数据存在时间差异, 野外数据是在2006年和2007年获取的, GLAS波形数据获取时间是2006年和2007年, HJ-1A/HSI数据是2010年获取的, 数据之间存在3a左右的间隔, 且GLAS数据是3、6、10月份和11月份, 高光谱数据是6月份的, 地面数据是9月份的, 在6-9月份之间是森林的生长旺季, 时间差异会对结果造成影响, 但是研究区位于以中龄林为主的长白山原始森林区, 树木的生长量有限, 数据获取时间不同步对森林地上生物量估测结果的影响应该较小。树种及林龄等都会对森林最大树高和森林郁闭度造成影响, 但其影响机理及影响权重尚不确定, 在今后的研究中可以尝试进行相应的研究, 以期提高模型准确性。目前主要是对汪清地区进行了实验研究, 对其他地区尚缺少相应年份的有效验证数据, 尚不能确定其是否具有普适应, 研究结果可以对其他地区的研究提供一定的参考, 并为今后进行区域乃至全国森林碳储量的研究提供了新的思路和方法。
| [1] | Piao S L, Fang J Y, Ciais P, Peylin P, Huang Y, Sitch S, Wang T. The carbon balance of terrestrial ecosystems in China. Nature , 2009, 458 (7241) : 1009–1013. DOI:10.1038/nature07944 |
| [2] | 刘双娜, 周涛, 舒阳, 戴铭, 魏林艳, 张鑫. 基于遥感降尺度估算中国森林生物量的空间分布. 生态学报 , 2012, 32 (8) : 2320–2330. |
| [3] | 杜虎, 曾馥平, 王克林, 宋同清, 温远光, 李春干, 彭晚霞, 梁宏温, 朱宏光, 曾昭霞. 中国南方3种主要人工林生物量和生产力的动态变化. 生态学报 , 2014, 34 (10) : 2712–2724. |
| [4] | Houghton R A, Hall F, Goetz S J. Importance of biomass in the global Carbon cycle. Journal of Geophysical Research:Biogeosciences , 2009, 114 (G2) : G00E03. DOI:10.1029/2009JG000935 |
| [5] | 黄金龙, 居为民, 郑光, 康婷婷. 基于高分辨率遥感影像的森林地上生物量估算. 生态学报 , 2013, 33 (20) : 6497–6508. |
| [6] | 娄雪婷, 曾源, 吴炳方. 森林地上生物量遥感估测研究进展. 国土资源遥感 , 2011, 23 (1) : 1–8. |
| [7] | 汤旭光, 刘殿伟, 王宗明, 贾明明, 董张玉, 刘婧怡, 徐文明. 森林地上生物量遥感估算研究进展. 生态学杂志 , 2012, 31 (5) : 1311–1318. |
| [8] | Boudreau J, Nelson R F, Margolis H A, Beaudoin A, Guindon L, Kimes D S. Regional aboveground forest biomass using airborne and spaceborne LiDAR in Québec. Remote Sensing of Environment , 2008, 112 (10) : 3876–3890. DOI:10.1016/j.rse.2008.06.003 |
| [9] | 张慧芳, 张晓丽, 黄瑜. 遥感技术支持下的森林生物量研究进展. 世界林业研究 , 2007, 20 (4) : 30–34. |
| [10] | 黄玫, 季劲钧, 曹明奎, 李克让. 中国区域植被地上与地下生物量模拟. 生态学报 , 2006, 26 (12) : 4156–4163. |
| [11] | Zhu X L, Liu D S. Improving forest aboveground biomass estimation using seasonal Landsat NDVI time-series. ISPRS Journal of Photogrammetry and Remote Sensing , 2015, 102 : 222–231. DOI:10.1016/j.isprsjprs.2014.08.014 |
| [12] | 郭云, 李增元, 陈尔学, 田昕, 凌飞龙. 甘肃黑河流域上游森林地上生物量的多光谱遥感估测. 林业科学 , 2015, 51 (1) : 140–149. |
| [13] | 王立海, 邢艳秋. 基于人工神经网络的天然林生物量遥感估测. 应用生态学报 , 2008, 19 (2) : 261–266. |
| [14] | Laurin G V, Chen Q, Lindsell J A, Coomes D A, Del frate F, Guerriero L, Pirotti F, Valentini R. Above ground biomass estimation in an African tropical forest with lidar and hyperspectral data. ISPRS Journal of Photogrammetry and Remote Sensing , 2014, 89 : 49–58. DOI:10.1016/j.isprsjprs.2014.01.001 |
| [15] | Xing Y Q, de Gier A, Zhang J J, Wang L H. An improved method for estimating forest canopy height using ICESat-GLAS full waveform data over sloping terrain:a case study in Changbai mountains, China. International Journal of Applied Earth Observation and Geoinformation , 2010, 12 (5) : 385–392. DOI:10.1016/j.jag.2010.04.010 |
| [16] | Lefsky M A, Harding D J, Keller M, Cohen W B, Carabajal C C, Del Bom Espirito-Santo F, Hunter M O, de Oliveira R. Estimates of forest canopy height and aboveground biomass using ICESat. Geophysical Research Letters , 2005, 32 (22) . DOI:10.1029/2005GL023971 |
| [17] | Enffle F, Heinzel J, Koch B. Accuracy of vegetation height and terrain elevation derived from ICESat/GLAS in forested areas. International Journal of Applied Earth Observation and Geoinformation , 2014, 31 : 37–44. DOI:10.1016/j.jag.2014.02.009 |
| [18] | Zhang G, Ganguly S, Nemani R R, White M A, Milesi C, Hashimoto H, Wang W L, Saatchi S, Yu Y F, Myneni R B. Estimation of forest aboveground biomass in California using canopy height and leaf area index estimated from satellite data. Remote Sensing of Environment , 2014, 151 : 44–56. DOI:10.1016/j.rse.2014.01.025 |
| [19] | Sun G, Ranson K J, Kimes D S, Blair J B, Kovacs K. Forest vertical structure from GLAS:an evaluation using LVIS and SRTM data. Remote Sensing of Environment , 2008, 112 (1) : 107–117. DOI:10.1016/j.rse.2006.09.036 |
| [20] | Pflugmacher D, Cohen W, Kennedy R, Lefsky M. Regional applicability of forest height and aboveground biomass models for the Geoscience Laser Altimeter System. Forest Science , 2008, 54 (6) : 647–657. |
| [21] | 黄克标, 庞勇, 舒清态, 付甜. 基于ICESat GLAS的云南省森林地上生物量反演. 遥感学报 , 2013, 17 (1) : 172–179. |
| [22] | 董立新, 吴炳方, 唐世浩. 激光雷达GLAS与ETM联合反演森林地上生物量研究. 北京大学学报:自然科学版 , 2011, 47 (4) : 703–710. |
| [23] | 谭炳香, 李增元, 陈尔学, 庞勇. 高光谱与多光谱遥感数据的森林类型识别. 东北林业大学学报 , 2005, 33 (增刊) : 61-63–70-70. |
| [24] | Schutz B E, Zwally H J, Shuman C A, Hancock D, Dimarzio J P. Overview of the ICESat mission. Geophysical Research Letters , 2005, 32 (21) . DOI:10.1029/2005GL024009 |
| [25] | Zhao X, Lu J, Gong A D, Xiao Z Q, Kang Q, Li Q. Operational spectrum Reconstruction of data from the Fourier transform hyperspectral imager onboard HJ-1A satellite. Chinese Science Bulletin , 2010, 55 (17) : 1808–1812. DOI:10.1007/s11434-010-3090-y |
| [26] | Deo R K.Modelling and Mapping of above-Ground Biomass and Carbon Sequestration in the Cool Temperate Forest of North-East China.Enschede:International Institute for Geo-Information Science and Earth Observation, 2008. |
| [27] | Duncanson L I, Niemann K O, Wulder M A. Estimating forest canopy height and terrain relief from GLAS waveform metrics. Remote Sensing of Environment , 2010, 114 (1) : 138–154. DOI:10.1016/j.rse.2009.08.018 |
| [28] | 李海涛, 顾海燕, 张兵, 高连如. 基于MNF和SVM的高光谱遥感影像分类研究. 遥感信息 , 2007 (5) : 12–15. |
| [29] | Yang W Z, Ni-Meister W, Lee S. Assessment of the impacts of surface topography, off-nadir pointing and vegetation structure on vegetation lidar waveforms using an extended geometric optical and radiative transfer model. Remote Sensing of Environment , 2011, 115 (11) : 2810–2822. DOI:10.1016/j.rse.2010.02.021 |
| [30] | Mahoney C, Kljun N, Los S O, Chasmer L, Hacker J M, Hopkinson C, North P R J, Rosette J A B, van Gorsel E. Slope estimation from ICESat/GLAS. Remote Sensing , 2014, 6 (10) : 10051–10069. DOI:10.3390/rs61010051 |
| [31] | 杜晓明, 蔡体久, 琚存勇. 采用偏最小二乘回归方法估测森林郁闭度. 应用生态学报 , 2008, 19 (2) : 273–277. |
| [32] | 李崇贵, 蔡体久. 森林郁闭度对蓄积量估测的影响规律. 东北林业大学学报 , 2006, 34 (1) : 15–17. |
| [33] | 朱教君, 康宏樟, 胡理乐. 应用全天空照片估计林分透光孔隙度(郁闭度). 生态学杂志 , 2005, 24 (10) : 1234–1240. |