 2016, Vol. 36
2016, Vol. 36文章信息
- 王应刚, 张婷, 段毅豪, 梁炜, 曹霄霄
- WANG Yinggang, ZHANG Ting, DUAN Yihao, LIANG Wei, CAO Xiaoxiao.
- 晋中盆地人类聚居地植物多样性分异规律
- Patterns of plant diversity and population distribution in the Jinzhong Basin
- 生态学报[J]. 2016, 36(20): 6556-6564
- Acta Ecologica Sinica[J]. 2016, 36(20): 6556-6564
- http://dx.doi.org/10.5846/stxb201507091454
-
文章历史
- 收稿日期: 2015-07-09
- 修订日期: 2015-12-08





2. 山西大学外国语学院, 太原 030006
2. School of Foreign Languages, Shanxi University, Taiyuan 030006, China
在全球范围内,城市化的发展对生态环境已经造成了一系列较大影响[1-3]。国外一些学者较早开始探索城市化对植被、土壤、水和大气的影响[4-5],我国学者也很早注意到城市化对生物的影响[6]。国内外学者沿城市化梯度对生物及环境等开展了较多研究[7-11]。目前,城市化梯度的研究主要有两种方法[12],一种是沿“城区-郊区-农区”梯度,如国外学者以“城区-郊区-农区”的生态样带为基础进行研究[13-17],马克明和傅伯杰对遵化市的植物多样性进行了研究[18]。彭羽则对廊坊市的本土植物多样性进行了分析[19]。一种方法是根据城市中土地利用类型的不同进行城市化梯度的研究[20-22]。如Porter研究了沿保护区-休闲区-高尔夫球场-住宅区-公寓-商业区植被和树冠碎片的变化[12]。区域城市化对不同人类聚居区的植物多样性产生了显著影响,德国学者Rudiger Wittig对欧洲中西部的6个城市建成区和美国的一个城市建成区的植物同时进行调查研究,揭示了一定地域尺度上植物多样性的同质化状况[23]。然而目前国内尚未有学者在区域尺度上沿“村庄-乡镇-县市”梯度,对其建成区内的野生植物与栽培植物物种多样性进行研究与分析。
晋中盆地属于山西省发展较快的区域,快速的城市化进程对区域生态环境造成了一定的影响。本文以晋中盆地作为研究区,沿村庄-乡镇-县市梯度,调查了各驻地建成区内的野生植物、栽培植物的种类,通过植物多样性研究,阐明晋中盆地区域城市化发展过程植物多样性规律,同时探索城市化过程中造成各村庄、乡镇和县市建成区内植物多样性分异的原因。为盆地内植物多样性保护提供根据。
1 研究区概况晋中盆地位于黄土高原东部山西省境内的汾河谷地,介于我国东南部湿润区与西北部干旱半干旱区之间的过度地带,属于半湿润地区,为典型的大陆性季风气候。地理坐标为111°45′—112°50′ E,36°48′—38°18′ N。盆地四面被丘陵、群山环抱,中央平原区海拔高程在760—840 m之间,年均降雨量为450—550 mm,年均气温在8—12℃之间,年平均湿度为55%—60%,晋中盆地是人类活动比较悠久的地区之一,因其发达的政治、经济和文化,使当地的城市化进程不但起步早而且发展速度比较快。到1950年时,晋中盆地的城市化水平已经达到21.26%,远高于全国10.60%的平均水平。经过半个世纪的历程,在改革开放及其他政策的的推动下,盆地内城镇人口数量急剧增加,到2003年时,盆地内城镇人口比重达到了60.33%。然而快速的城市化对区域气候、地下水、地表水及盆地内的植物多样性产生了较大的影响,引起了一系列的生态问题。
2 研究方法 2.1 样方设置依据植物多样性研究中的样地布设原理和统计学分析的需要,在盆地内3785个自然村中,对每个村庄进行编号,运用随机抽样法,选出86个村庄驻地,组成样本Ⅰ,在样本内的每个村庄驻地建成区内,随机抽取16个100 m×20 m的样方进行调查,每个村庄的调查面积为32000 m2。用同样方法在133个乡镇中,随机抽出48个乡镇驻地,组成样本Ⅱ,每个乡镇驻地建成区内随机调查21个100 m×20 m样方,每个乡镇的调查面积为42000 m2。同理,在太原市、介休市、孝义市、汾阳市、榆次市、平遥县、太谷县、祁县、文水县、交城县、清徐县、阳曲县共12个县市中,随机抽出9个县市驻地,组成样本Ⅲ,每个县市驻地建成区内随机调查26个100 m×20 m的样方,每个县市的调查面积为52000 m2。各驻地分布位置如图 1所示。

|
| 图 1 采样点分布示意图 Fig. 1 Location of the sampling sites |
2012年6月至10月初对各驻地建成区样方内的植物物种进行了现场调查,由于建成区内受到人为干扰较强,已无法进行群落单元研究,故本次仅调查每个样方内物种的有无。在每个设置的100 m×20 m样方内,对每一种植物(包括乔木、灌木、草本)的名称及是否为野生植物或人工栽培植物等进行了记录。本文中记录的植物均指能够在户外越冬,野生植物指能自然生长繁衍不依赖人为栽植;栽培植物是指人为有意识在户外栽种,由栽培植物逸生的植物亦归类为栽培植物。
同时,对每个村庄建成区、乡镇建成区及县市建成区内的人口密度、建筑物压占土地率、地面硬化率进行调查。
2.3 数据处理对每个村庄驻地建成区内调查的16个样方内的植物进行综合统计,获得该村庄驻地建成区内记录到的所有野生植物、栽培植物的名称及物种数,同理,得到每个乡镇驻地建成区、县市驻地建成区内记录到的所有野生植物、栽培植物的名称及物种数。然后使用Excel、Origin8、SPSS13.0软件进行制图分析。
(1) Gleason丰富度指数
由于在村庄建成区、乡镇建成区及县市建成区内的人类活动强烈,自然植被已经被摧毁,只能见到不连续的零星分布的植物,在此情况下,已经无法考虑各个物种之间的数量比例关系。因此,本研究选用不涉及各个物种之间的数量比例关系的Gleason index丰富度指数,计算公式为:
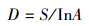
|
式中, S为物种数,A为调查面积,D为Gleason物种丰富度指数。用此公式分别计算被调查每个村庄驻地建成区、乡镇驻地建成区、县市驻地建成区的物种丰富度。
(2) Beta多样性指数
Whittaker Beta多样性是生物多样性的重要组成部分,人类活动对能够对其产生重要的影响[24]。本文将不同等级驻地建成区分别看作不同类型的取样生境,来研究因人类干扰所导致的生境中物种组成变化[25]。
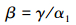
|
式中,γ为某取样生境中的总物种数,α为某生境类型(村庄生境、乡镇生境及县市生境)所有取样点物种的平均数(本研究采用平均丰富度指数D代替α开展计算)
(3) 相似性指数
Jaccard[26]相似性指数能够分析在不同城市化梯度即不同样本生境间出现相同物种的状况。
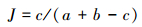
|
式中,c为两样本间共有物种,a和b分别为两个样本的物种数。
3 结果分析 3.1 物种丰富度运用Gleason index对86个村庄驻地建成区、48个乡镇驻地建成区、9个县市驻地建成区的野生植物、栽培植物及总物种丰富度进行分析(图 2—图 4)。

|
| 图 2 村庄驻地物种丰富度 Fig. 2 Species richness in villages seats |

|
| 图 3 乡镇驻地物种丰富度 Fig. 3 Species richness in towns seats |

|
| 图 4 县市驻地物种丰富度 Fig. 4 Species richness in cities seats |
在调查取样的村庄驻地建成区中,野生植物丰富度的大小对于总植物物种丰富度起主导性作用,人工栽培植物的丰富度大多处于较低水平,且两者丰富度相差较大。在乡镇驻地建成区中,野生植物丰富度高于栽培植物丰富度,野生植物丰富度绝大多数低于30而栽培植物丰富度大多介于10—20之间,两者共同促进了总植物物种丰富度的增加。在取样的9个县市驻地建成区中,野生植物丰富度稍低于栽培植物丰富度,在个别县市野生植物丰富度大于栽培植物丰富度,同时城市9的栽培植物丰富度明显较高是由于其采样点为山西省会城市太原市。
对3个样本内的野生植物、栽培植物及总植物物种的平均丰富度分析。如图 5所示,沿“村庄-乡镇-县市”梯度,野生植物丰富度减少,栽培植物丰富度增加,总植物物种丰富度先降低后增加。

|
| 图 5 各样本平均丰富度指数 Fig. 5 Mean species richness of samples |
使用SPSS13.0对3个样本间的野生植物丰富度、栽培植物丰富度和总物种丰富度进行独立样本T检验,结果如表 1所示:野生植物和栽培植物,在各个样本间均有显著性差异。而从总植物物种丰富度来看,村庄与乡镇、乡镇与县市有显著差异,而村庄与县市则无明显差异。
| 植物类别 Plants categories | 野生植物Wild plants | 栽培植物Cultivated plants | 总植物Total plants | ||||||||
| P | t | df | P | t | df | P | t | df | |||
| Ⅰ—Ⅱ | 0.000 | 8.475* | 132 | 0.002 | 3.137* | 132 | 0.000* | 4.860 | 132 | ||
| Ⅰ—Ⅲ | 0.000 | 9.505* | 93 | 0.000 | 8.343* | 93 | 0.843 | 0.222 | 93 | ||
| Ⅱ—Ⅲ | 0.005 | 2.938* | 55 | 0.000 | 3.773* | 55 | 0.013* | 1.890 | 55 | ||
| *P < 0.05 | |||||||||||
β多样性指数反映了人类活动对植物物种多样性的影响,图 6表明:沿村庄-乡镇-县市梯度,植物的β多样性均降低,野生植物多样性指数分别为5.18、5.05、3.51,栽培植物的多样性指数为4.56、3.99、2.96。总植物物种多样性指数为5.01、4.65、3.22。说明,随着城市化强度的增强,各植物物种的多样性降低,物种的同质性增加[27]。

|
| 图 6 各样本β多样性指数 Fig. 6 β-diversity of samples |
Jaccard指数反映了不同样本生境间物种组成之间的相似程度,指数越小,说明物种组成差异越大[28]。
从图 8可知,Jaccard相似性指数,就栽培植物而言,县市-村庄(0.53)、县市-乡镇(0.61)均低于村庄-乡镇(0.78)。同样,野生植物,县市-村庄(0.41)、县市-乡镇(0.52)均低于村庄-乡镇(0.64),说明无论是野生植物还是栽培植物,县市与乡镇、村庄差异较为明显,而村庄与乡镇则有较高的相似性。

|
| 图 8 各样本间相似性指数 Fig. 8 Similarity index between samples |
对村庄、乡镇和县市的野生共有物种进行分析,发现3个样本间共有物种较多集中于菊科、禾本科和藜科3类,而栽培植物则集中于蔷薇科、杨柳科等,具体植物见表 2。从表中可以看出,野生植物大多为一年生草本,耐践踏,且广布于世界温带地区。栽培植物大多在各地广为栽培,兼绿化观赏及较高经济适用性。
| 野生植物Wild plants | 栽培植物Cultivated plants |
| 马唐Digitaria sanguinalis | 月季花Rosa chinensis |
| 狗尾草Setaria viridis | 桃Amygdalus persica |
| 黄鹌菜Youngia japonica | 刺槐Robinia pseudoacacia |
| 蒲公英Taraxacum mongolicum | 菊花Dendranthema morifolium |
| 藜Chenopodium album | 青杨Populus cathayana |
| 葎草Humulus scandens | 垂柳Salix babylonica |
| 牛皮消Cynanchum auriculatum | 旱柳Salix matsudana |
| 马齿苋Portulaca oleracea | 枣 Ziziphus jujuba |
对比分析3个样本特有物种发现,村庄独有的野生植物较多,栽培植物仅有向日葵(Helianthus annuus)和番茄(Lycopersicon esculentum)两种。县市独有的栽培植物较多,独有的小香蒲(Typha minima)、宽叶香蒲(Typha latifolia)、灯心草(Juncus effusus)等几种野生植物也仅均出现1次。乡镇则没有独有的野生植物和栽培植物。村庄和县市的主要独有物种见表 3。
| 村庄独有植物Unique species in villages | 县市独有植物Unique species in cities |
| 酸枣Ziziphus jujuba spinosa | 黄杨Buxus sinica |
| 飞廉Carduus nutans | 黑麦Secale cereale |
| 鼠掌老鹳草Geranium sibiricum | 野牛草Buchloe dactyloides |
| 荆条Vitex negundo heterophylla | 女贞Ligustrum lucidum |
| 牵牛Pharbitis nil | 榆叶梅Amygdalus triloba |
| 牛蒡Arctium lappa | 奥氏蓼Fallopia aubertii |
| 木地肤Kochia prostrata | 二球悬铃木Platanus acerifolia |
| 盐地碱蓬Suaeda salsa | 雪松Cedrus deodara |
样地调查发现,城市化水平的差异与人为干扰强度的不同造成了植物多样性分异。沿村庄-乡镇-县市梯度,城市化程度增加,主要表现在人口密度加大、建筑物压占土地率及地面硬化率升高。如图 7,调查显示,盆地内村庄人口密度较小,为4089人/km2,乡镇与县市的人口密度分别为5525人/km2与9875人/km2。地面硬化率由村庄、乡镇与县市依次为62%、73%及92%左右。建筑物压占土地率亦同城市化水平高低一致,村庄最低为34%,乡镇稍高为42%,县市最高为47%。

|
| 图 7 各样本城市化水平 Fig. 7 Urbanzation of samples |
较低的城市化水平与低强度的人为破坏使得村庄内野生植物的丰富度最高,而县市内人口密度大,污染物排放及除草践踏等人为干扰活动较强,使得县市建成区内的野生植物丰富度最低,乡镇的城市化发展程度介于村庄与县市之间,野生植物丰富度居中。而各样地中栽培植物的丰富度则主要受到人为力量的主导。由于县市发展迅速,经济实力较强,对城市内绿化及公园建设投入大量资金,引入了较多的可供观赏绿化的外来物种,因而其栽培植物物种丰富度最高。乡镇发展次之,栽培植物丰富度较高。村庄的经济发展相对较弱,村庄建成区内用于绿化美化的栽培植物种类较少,大多是农户自己栽植的较常见的的果木,所以栽培植物丰富度最低。而总植物物种丰富度则呈现出乡村、县市较高,乡镇较低的情况,这是由于乡镇建成区要承受比村庄更大的城市化建设,野生植物的生长受到一定程度干扰,同时因其经济发展水平低于县市,人工栽培植物种类较低,故而总植物物种丰富度低于村庄与县市。显然,本文研究因城市化梯度设置的特殊性与Connell提出的“中度干扰理论”[29]是不相关且不矛盾的。
植物的β多样性指数沿村庄-乡镇-县市梯度降低,因为沿此梯度,人口密度增加,地面硬化率升高,建筑物压占土地率增大,一些野生植物因生境受到土地利用方式的改变而消失,生存下来的物种大都是生命力强且耐践踏的禾本科、藜科类等我国广泛分布的习见植物[19],出现了较大的同质化现象。而县市的栽培植物β多样性最低则是由于在各县市发展相当的情况下,因村庄乡镇及县市建设均是为满足人民生活需要,其建筑结构与城市热岛效应等气候条件相似[30],绿化引种植物亦较类同造成的,而县市在这方面表现的更为强烈。说明城市化造成了各植物物种的均一化,这已被众多学者研究证明[31-32]。此外,因村庄与乡镇的发展差距较小,村庄与县市的发展程度差距大,使得村庄与乡镇植物相似性最高,乡镇与县市次之,村庄与县市的相似性最低。
5 结论德国学者Rudiger Wittig指出,在分析城市化导致物种同质化的过程中,“纯粹”的城市化栖息地会比城市与非城市混合的栖息地显示更高的同质化程度[23]。本文在晋中盆地区域尺度上,选择了分别受城市化影响较强、居中及较弱的县市建成区、乡镇建成区和村庄建成区,故而能够更加准确地分析城市化对区域内植物多样性的影响。同时,通过选择调查野生植物与栽培植物两类受人为干扰较强的植物类别,亦能较好指示城市化过程中区域内植物多样性现状。在晋中盆地,村庄建成区内野生植物受人为干扰较轻,栽培植物引入很少,导致其总物种丰富度没有明显降低;乡镇驻地建成区野生植物物种丰富度在城市化建设过程中受到较大影响,而又没有较多地引种栽培植物,导致其总物种丰富度明显降低,这是在未来乡镇建设过程中需要注意的一个问题;县市建成区发展较为迅速,对野生植物生长和分布影响显著,但在县市建成区大量引入栽培植物进行绿化,加之在人为干预下使建成区内土地环境的异质性明显降低和绿化植物物种类同性较高,一方面导致其总物种丰富度明显增加,另一方面也导致各县市建成区内物种的同质化。总之,在村庄、乡镇和县市建成区今后的发展过程中,尽量保留一些野生植物群落及其生境,合理选用和引入适合当地气候环境特点的多种类型的栽培植物进行绿化,既提高植物物种多样性又避免物种同质化趋势。
| [1] | Harveson P M, Lopez R R, Collier B A, Slilvy N J. Impacts of urbanization on Florida Key deer behavior and population dynamics. Biological Conservation , 2007, 134 (3) : 321–331. DOI:10.1016/j.biocon.2006.07.022 |
| [2] | Meyer W B, Turner B L Ⅱ. Human population growth and global land-use/cover change. Annual Review of Ecology and Systematics , 1992, 23 (1) : 39–61. DOI:10.1146/annurev.es.23.110192.000351 |
| [3] | Moore M, Gould P, Keary B S. Global urbanization and impact on health. International Journal of Hygiene and Environmental Health , 2003, 206 (4/5) : 269–278. |
| [4] | Seinfeld J H. Urban air pollution:state of the science. Science , 1989, 243 (4892) : 745–752. DOI:10.1126/science.243.4892.745 |
| [5] | McDonnell M J, Pickett S T A. Ecosystem structure and function along urban-rural gradients:an unexploited opportunity for ecology. Ecology , 1990, 71 (4) : 1232–1237. DOI:10.2307/1938259 |
| [6] | 王献溥. 城市化对生物多样性的影响. 农村生态环境 , 1996, 12 (4) : 32–36. |
| [7] | McDonnell M J, Pickett S T A, Pouyat R V. The application of the ecological gradient paradigm to the study of urban effects//McDonnell M J, Pickett S T A, eds. Humans as Components of Ecosystems. New York:Springer, 1993:175-189. |
| [8] | Blair R B. Land use and avian species diversity along an urban gradient. Ecological Applications , 1996, 6 (2) : 506–519. DOI:10.2307/2269387 |
| [9] | Blair R B, Launer A E. Butterfly diversity and human land use:Species assemblages along an urban grandient. Biological Conservation , 1997, 80 (1) : 113–125. DOI:10.1016/S0006-3207(96)00056-0 |
| [10] | McDonnell M J, Hahs A K. The use of gradient analysis studies in advancing our understanding of the ecology of urbanizing landscapes:current status and future directions. Landscape Ecology , 2008, 23 (10) : 1143–1155. DOI:10.1007/s10980-008-9253-4 |
| [11] | Vakhlamova T, Rusterholz H P, Kanibolotskaya Y, Baur B. Changes in plant diversity along an urban-rural gradient in an expanding city in Kazakhstan, Western Siberia. Landscape and Urban Planning , 2014, 132 : 111–120. DOI:10.1016/j.landurbplan.2014.08.014 |
| [12] | Porter E E, Forschner B R, Blair R B. Woody vegetation and canopy fragmentation along a forest-to-urban gradient. Urban Ecosystems , 2001, 5 (2) : 131–151. DOI:10.1023/A:1022391721622 |
| [13] | McDonnell M J, Pickett S T A, Groffman P, Bohlen B, Pouyat R V, Zipperer W C, Parmelee R W, Carreiro M M, Medley K. Ecosystem processes along an urban-to-rural gradient. Urban Ecosystems , 1997, 1 (1) : 21–36. DOI:10.1023/A:1014359024275 |
| [14] | Pouyat R V, McDonnell M J. Heavy metal accumulations in forest soils along an urban-rural gradient in southeastern New York, USA. Water, Air, and Soil Pollution , 1991, 57 (1) : 797–807. |
| [15] | Pouyat R V, Carreiro M M, McDonnell M J, Pickett S T A, Groffman P M, Parmelee R W, Medley K E, Zipperer W C. Carbon and nitrogen dynamics in oak stands along an urban-rural gradient//McFee W W, Kelly J M, eds. Carbon Forms and Functions in Forest Soils. Madison:Soil Science Society of America Inc., 1995:569-587. |
| [16] | Goldman M B, Groffman P M, Pouyat R V, McDonnell M J, Pickett S T A. CH4 uptake and N availability in forest soils along an urban to rural gradient. Soil Biology and Biochemistry , 1995, 27 (3) : 281–286. DOI:10.1016/0038-0717(94)00185-4 |
| [17] | Williams N S G, Morgan J W, Mcdonnell M J, Mccarthy M A. Plant traits and local extinctions in natural grasslands along an urban-rural gradient. Journal of Ecology , 2005, 93 (6) : 1203–1213. DOI:10.1111/j.1365-2745.2005.01039.x |
| [18] | 马克明, 傅伯杰, 郭旭东. 农业区城市化对植物多样性的影响:遵化的研究. 应用生态学报 , 2001, 12 (6) : 837–840. |
| [19] | 彭羽, 刘雪华, 薛达元, 邵小明, 姜炎彬. 城市化对本土植物多样性的影响——以廊坊市为例. 生态学报 , 2012, 32 (3) : 723–729. |
| [20] | Hostetler N E, McIntyre M E. Effects of urban land use on pollinator (Hymenoptera:Apoidea) communities in a desert metropolis. Basic and Applied Ecology , 2001, 2 (3) : 209–218. DOI:10.1078/1439-1791-00051 |
| [21] | McIntyre N E, Rango J, Fagan W F, Faeth S H. Ground arthropod community structure in a heterogeneous urban environment. Landscape and Urban Planning , 2001, 52 (4) : 257–274. DOI:10.1016/S0169-2046(00)00122-5 |
| [22] | Dorney J R, Guntenspergen G R, Keough J R, Stearns F. Composition and structure of an urban woody plant community. Urban Ecology , 1984, 8 (1/2) : 69–90. |
| [23] | Wittig R, Becker U. The spontaneous flora around street trees in cities-A striking example for the worldwide homogenization of the flora of urban habitats. Flora-Morphology, Distribution, Functional Ecology of Plants , 2010, 205 (10) : 704–709. DOI:10.1016/j.flora.2009.09.001 |
| [24] | 陈圣宾, 欧阳志云, 徐卫华, 肖燚. Beta多样性研究进展. 生物多样性 , 2010, 18 (4) : 323–335. |
| [25] | 卢训令, 汤茜, 梁国付, 丁圣彦. 黄河下游平原不同非农生境中植物多样性. 生态学报 , 2015, 35 (5) : 1527–1536. |
| [26] | Burton M L, Samuelson L J, Pan S. Riparian woody plant diversity and forest structure along an urban-rural gradient. Urban Ecosystems , 2005, 8 (1) : 93–106. DOI:10.1007/s11252-005-1421-6 |
| [27] | McKinney M L. Urbanization as a major cause of biotic homogenization. Biological Conservation , 2006, 127 (3) : 247–260. DOI:10.1016/j.biocon.2005.09.005 |
| [28] | 马克平, 叶万辉, 于顺利, 马克明, 王巍, 关文彬. 北京东灵山地区植物群落多样性研究Ⅷ.群落组成随海拔梯度的变化. 生态学报 , 1997, 17 (6) : 593–600. |
| [29] | Connell J H. Diversity in tropical rain forests and coral reefs. Science , 1978, 199 (4335) : 1302–1310. DOI:10.1126/science.199.4335.1302 |
| [30] | Knapp S, Wittig R. An analysis of temporal homogenisation and differentiation in Central European village floras. Basic and Applied Ecology , 2012, 13 (4) : 319–327. DOI:10.1016/j.baae.2012.06.004 |
| [31] | McKinney M L. Effects of urbanization on species richness:a review of plants and animals. Urban Ecosystems , 2008, 11 (2) : 161–176. DOI:10.1007/s11252-007-0045-4 |
| [32] | Byers J E. Impact of non-indigenous species on natives enhanced by anthropogenic alteration of selection regimes. Oikos , 2002, 97 (3) : 449–458. DOI:10.1034/j.1600-0706.2002.970316.x |