 2016, Vol. 36
2016, Vol. 36文章信息
- 陈国鹏, 鲜骏仁, 曹秀文, 刘锦乾, 杨永红, 雷炜
- CHEN Guopeng, XIAN Junren, CAO Xiuwen, LIU Jinqian, YANG Yonghong, LEI Wei.
- 林窗对岷江冷杉幼苗生存过程的影响
- Effects of canopy gap on the survival dynamics of Abies faxoniana seedlings in a subalpine coniferous forest
- 生态学报[J]. 2016, 36(20): 6475-6486
- Acta Ecologica Sinica[J]. 2016, 36(20): 6475-6486
- http://dx.doi.org/10.5846/stxb201503170515
-
文章历史
- 收稿日期: 2015-03-17
- 网络出版日期: 2016-01-22

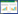




2. 四川农业大学环境学院, 温江 611130
2. College of the Environmental sciences, Sichuan Agricultural University, Wenjiang 611130, China
作为群落的重要组成部分, 种群的结构不仅对群落结构有直接影响, 而且能客观体现群落的发展趋势和动态[1-2]。植物种群数量动态是其个体生存能力与环境相互作用的结果[3], 开展种群年龄结构及其数量动态的研究在反映种群生态学特征、展现种群与环境适应的结果、阐明群落发展趋势等方面都具有重要意义[4-6]。林窗是森林群落中林冠树木死亡而在林地上形成的不连续的林中空地[7], 在森林植被更新和演替过程中发挥重要作用[8]。林窗的形成使森林群落内部的光、热、水等环境因子发生显著的变化[8-9], 引起林窗内外更新幼苗定居、生长的分异[7, 10-12]。已有研究多从静态视角去揭示林窗对幼苗密度及生长的影响[12-13], 缺乏对幼苗种群动态的了解。而开展林窗内外幼苗种群结构与动态的追踪将有助于充分认识森林更新过程中林窗的作用及影响机制。
以岷江冷杉(Abiesfaxoniana)为主要建群种的西南亚高山暗针叶林是青藏高原东缘山地森林植被的主体, 也是长江上游水源涵养林中起重要作用的植被类型[14-16]。岷江冷杉林的更新一直是学者们关注的热点, 程伟等[16]、康东伟等[17]、谭留夷等[18]均指出岷江冷杉种群动态过程中幼苗储量丰富、但高达90%的幼苗死亡率严重影响着种群的顺利更新[16]。岷江冷杉更新过程中最关键的幼苗阶段一直处于不稳定状态[19-20], 箭竹密度、苔藓厚度等均将阻碍岷江冷杉幼苗的发生和生长[14-15, 21]。林窗更新是岷江冷杉群落更新的主要方式[12-14], 其更新幼苗的种群结构、生存过程、数量变化特征以及抵御外界随机干扰的能力是否较林下种群优越?相关报道却鲜见, 这在一定程度上制约着对岷江冷杉林更新过程的认知和对林窗作用的评价。
本研究以岷江冷杉更新过程中最关键的幼苗阶段及林窗的作用为关注点, 采用空间代替时间的方法, 在岷江冷杉林中开展幼苗更新状况调查, 分析林窗内和林下岷江冷杉幼苗的年龄结构、存活曲线、死亡率和消失率曲线、生存状况及数量波动特征, 以阐明林窗对岷江冷杉幼苗种群结构特征、生存过程及数量动态趋势的影响, 以期为深入认识亚高山暗针叶林群落动态与稳定性奠定基础, 并为岷江冷杉林的更新保育提供基础数据和理论依据。
1 研究区概况研究区位于四川省平武县的王朗国家级自然保护区, 地理位置介于32°49′—33°02′N, 103°55′—104°10′E, 海拔2300—4980 m。该区域属于丹东-松潘半湿润气候, 年平均气温2.9 ℃, 年积温(≥10 ℃)1056.5 ℃, 最高和最低气温分别为26.2 ℃和-17.8℃, 多年平均降水量859.9 mm、并集中在5—8月。森林植被主要由原始云杉(Picea spp.)冷杉林、针阔混交林、桦木(Betulaspp.)林和人工针叶林组成。土壤包括有山地棕壤、山地暗棕壤、亚高山草甸土、高山草甸土和高山流石滩荒漠土。岷江冷杉林主要分布在研究区海拔2600—3500 m的区域内[9, 11-22]。
2 研究方法 2.1 调查方法在对研究区初步踏查后, 选择位于大窝凼的岷江冷杉-缺苞箭竹(Fargesia denudata)-苔藓林为对象(样地中心坐标:33°00′11.17″N, 104°01′34.43″E, 海拔2910 m), 平均林龄180 a, 平均胸径和树高分别为45 cm和38 m, 林分郁闭度介于0.4—0.6之间。乔木层的优势种为岷江冷杉, 密度可达1533株/hm2;其次为紫果云杉(Picea purpurea)、大翅色木槭(Acer mono var. macropterum)和方枝圆柏等(Sabina saltuaria)(表 1)。灌木层主要有缺苞箭竹, 密度为9546株/hm2;其次为苦糖果(Lonicera stanishii)、悬钩子(Rubus corchorifolius)等多种灌木和幼苗, 平均盖度40%, 平均高度1.4 m(表 2)。草本层主要有紫花碎米荠(Cardamine tangutorum)、鹿药(Smilacina japonica)和铁线蕨(Adiantum capillus-veneris)等, 平均盖度30%, 平均高度30 cm;苔藓层主要为山羽藓(Abietinella abietina)和塔藓(Hylocomium splendens), 平均盖度90%以上, 平均厚度8 cm[11, 22]。
| 树种Species | 密度 Density/(株/hm2) |
树高 Height/m |
胸径 DBH/cm |
重要值 Important value/% |
| 岷江冷杉Abies faxoniana | 1533 | 36.89 | 40.35 | 55.59 |
| 紫果云杉Picea purpurea | 400 | 46.71 | 64.14 | 31.83 |
| 大翅色木槭Acer mono var.macropterum | 100 | 13.67 | 15.33 | 4.92 |
| 方枝圆柏Sabina saltuaria | 68 | 22 | 28.13 | 3.60 |
| 红桦Betula albo-sinensis | 34 | 18 | 23.24 | 1.73 |
| 红毛花楸Sorbus rufopilosa | 73 | 15 | 16.37 | 2.33 |
| 种名Species | 平均高度 Averagingheight /m |
密度 Density /(株/hm2) |
重要值 Important value/% |
| 缺苞箭竹Fargesiadenudata | 2.68 | 9546 | 21.24 |
| 苦糖果Lonicerastanishii | 1.46 | 3794 | 12.71 |
| 岷江冷杉Abiesfaxoniana | 0.25 | 4267 | 10.48 |
| 悬钩子Rubuscorchorifolius | 0.97 | 2674 | 10.06 |
| 秀丽莓Rubusamabilis | 1.16 | 2547 | 9.43 |
| 大翅色木槭Acer monovar.macropterum | 1.39 | 2385 | 6.92 |
| 紫花卫矛Euonymus porphyreus | 2.43 | 648 | 3.21 |
| 紫果云杉Piceapurpurea | 0.91 | 567 | 3.07 |
| 山梅花Philadelphusmagdalenae | 2.96 | 572 | 3.01 |
| 红桦Betulaalbo-sinensis | 2.22 | 357 | 2.37 |
| 无柄杜鹃Rhododendron watsonii | 0.96 | 287 | 2.05 |
| 冰川茶镳子Ribesglaciale | 1.93 | 504 | 1.61 |
| 陇西忍冬Loniceratangutica | 2.06 | 264 | 1.6 |
| 红毛五加Acanthopanaxgiraldii | 1.58 | 34 | 1.53 |
| 花楸Sorbus sp. | 2.38 | 237 | 1.49 |
| 忍冬Loniceraspp. | 2.29 | 356 | 1.29 |
| 峨眉蔷薇Rosa omeiensis | 1.9 | 235 | 1.01 |
沿垂直于等高线的方向随机设置长300 m、宽2 m的调查样带3条, 样带间隔50 m以上。在整个样带内, 林冠林窗(后文简称林窗)面积为372 m2(占20.67%), 林下面积为1428 m2(占79.33%)。分别测量林窗和林下冷杉幼苗(规定高度小于100 cm[11])基径和苗高。共调查到幼苗768株(林窗480株, 林下288株), 基径介于1.00—13.69 mm(林窗为1.00—13.18 mm, 林下为1.14—13.69 mm), 苗高介于7.4—82.5 cm(林窗为8.3—82.5 cm, 林下为7.4—63.6 cm)。采用“空间推导时间”的方法, 用基径推导苗龄, 以1.0 mm的基径间隔为1龄级划分岷江冷杉幼苗龄级[11, 20, 23], 将林窗、林下和总体的幼苗均划分为13个龄级。
2.2 静态生命表编制统计每个龄级的幼苗数, 编制岷江冷杉幼苗的静态生命表[24-25]。计算公式如下:

|
式中, x为龄级数;Ax是在x龄级内的实际存活数;ax为经过匀滑处理后Ax的修正值[18];lx为在x龄级开始时标准化存活个体数(一般转化值为1000);dx为从x到x+1龄级间隔期内标准化死亡数;qx为从x到x+1龄级间隔期间死亡率;Lx为从x到x+1龄级间隔期间存活的个体数;Tx为从x龄级到超过x龄级的个体总数;ex为进入x龄级个体的生命期望寿命;Kx为消失率。
2.3 存活曲线拟合以存活数lx为纵坐标, 以龄级为横坐标, 绘制岷江冷杉幼苗的存活曲线, 根据各年龄级的个体数, 在Origin 8.0软件(OriginLab Corporation, USA)中采用指数函数和幂函数进行存活数和龄级关系的拟合, 依据决定系数、F检验值和AIC值(赤池信息量准则)判定模型拟合的效果。如果指数方程Nx=N0e-bx拟合效果好, 存活曲线为DeevyⅡ型;若幂函数拟合效果好, 存活曲线符合DeevyⅢ型[4]。
2.4 生存分析采用生存率函数S(i)、积累死亡率函数F(i)、死亡密度函数f(i)和危险率函数λ(i)进行幼苗种群结构动态分析, 以阐明其生存规律[26], 公式如下:

|
式中, Pi为存活频率;hi为区间长度, 即龄级宽度。
2.5 种群动态定量化分析采用陈晓德种群动态数量化方法描述岷江冷杉幼苗种群数量变化动态[27], 计算公式如下:
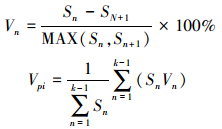
|
式中, Vn为种群从n到n+1级的个体数量变化;Vpi为整个种群结构的数量变化动态指数;Sn、Sn+1分别为第n和n+1级种群个体数;k为种群大小级数量。
Vpi仅适用于不考虑未来外部环境干扰的种群结构动态的比较, 当考虑未来的外部干扰时:
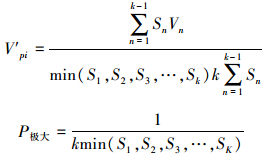
|
Vn值区间为[-1, 1], Vn取正、负、零值时分别反映种群两相邻年龄级个体数量的增长、衰退、稳定的结构动态关系, Vpi、Vpi′取正、负、零值时反映的动态关系与Vn一致。P为种群对外界随机干扰所承担的风险概率, 只有当P的值为最大时才对种群动态Vpi构成最大的影响。
此外, 为刻画种群的动态过程, 采用谱分析方法对幼苗的周期性波动进行分析[28-30]。
3 结果分析 3.1 年龄结构如图 1所示, 林窗、林下和总体的幼苗年龄结构金字塔均为中间大两头小的纺锤型, 以4龄级的个体数最多, 分别为90(18.75%)、44(15.28%)和134(17.45%);其次为3龄级的个体数, 再次为5龄级个体数;13龄级的个体数最少, 分别为1、2、3, 所占比例分别为0.21%、0.69%和0.39%;林下10龄级的幼苗稍有波动, 个体数量多于9龄级个体数。

|
| 图 1 岷江冷杉幼苗年龄结构图 Fig. 1 The age structure of A. faxoniana seedlings |
岷江冷杉幼苗种群的生命期望(ex)波动见表 3—表 5所示, 最高生命期望均出现在1龄级, 以林下最高(4.212)、总体次之(3.217)、林窗最小(2.837)。随着龄级的增大, 种群的生命期望呈下降趋势, 但林窗种群在10龄级和11龄级时出现反弹;到12龄级时, 林下、林窗和总体种群的生命期望分别降为0.783、0.192和0.428, 以林下的降幅最大。同时, 随着龄级增加, 林窗种群对总体种群生命期望的影响逐渐减弱, 在1—6龄级阶段, 总体种群的生命期望更接近于林窗种群的生命期望, 之后则更接近于林下种群。
| 龄级 Age stage |
Ax | ax | lx | dx | qx | Lx | Tx | ex | lnlx | Kx | Sx |
| 1 | 22 | 174 | 1000 | 167 | 0.167 | 917 | 2837 | 2.837 | 6.908 | 0.182 | 0.833 |
| 2 | 69 | 145 | 833 | 161 | 0.193 | 753 | 2084 | 2.500 | 6.725 | 0.215 | 0.807 |
| 3 | 90 | 117 | 672 | 144 | 0.214 | 601 | 1483 | 2.206 | 6.511 | 0.240 | 0.786 |
| 4 | 90 | 92 | 529 | 109 | 0.207 | 474 | 1009 | 1.908 | 6.270 | 0.231 | 0.793 |
| 5 | 73 | 73 | 420 | 98 | 0.233 | 371 | 638 | 1.521 | 6.039 | 0.265 | 0.767 |
| 6 | 56 | 56 | 322 | 121 | 0.375 | 261 | 377 | 1.171 | 5.774 | 0.470 | 0.625 |
| 7 | 35 | 35 | 201 | 92 | 0.457 | 155 | 222 | 1.102 | 5.304 | 0.611 | 0.543 |
| 8 | 19 | 19 | 109 | 0 | 0.000 | 109 | 112 | 1.030 | 4.693 | 0.000 | 1.000 |
| 9 | 18 | 19 | 109 | 86 | 0.789 | 66 | 46 | 0.425 | 4.693 | 1.558 | 0.211 |
| 10 | 3 | 4 | 23 | 11 | 0.500 | 17 | 29 | 1.267 | 3.135 | 0.693 | 0.500 |
| 11 | 2 | 2 | 11 | 0 | 0.000 | 11 | 18 | 1.533 | 2.442 | 0.000 | 1.000 |
| 12 | 2 | 2 | 11 | 6 | 0.500 | 9 | 9 | 0.783 | 2.442 | 0.693 | 0.500 |
| 13 | 1 | 1 | 6 | 1.749 |
| 龄级 Age stage |
Ax | ax | lx | dx | qx | Lx | Tx | ex | lnlx | Kx | Sx |
| 1 | 17 | 64 | 1000 | 125 | 0.125 | 938 | 4212 | 4.212 | 6.908 | 0.134 | 0.875 |
| 2 | 32 | 56 | 875 | 109 | 0.125 | 820 | 3392 | 3.876 | 6.774 | 0.134 | 0.875 |
| 3 | 41 | 49 | 766 | 94 | 0.122 | 719 | 2673 | 3.491 | 6.641 | 0.131 | 0.878 |
| 4 | 44 | 43 | 672 | 109 | 0.163 | 617 | 2056 | 3.060 | 6.510 | 0.178 | 0.837 |
| 5 | 36 | 36 | 563 | 94 | 0.167 | 516 | 1540 | 2.738 | 6.332 | 0.182 | 0.833 |
| 6 | 29 | 30 | 469 | 78 | 0.167 | 430 | 1111 | 2.369 | 6.150 | 0.182 | 0.833 |
| 7 | 25 | 25 | 391 | 78 | 0.200 | 352 | 759 | 1.943 | 5.968 | 0.223 | 0.800 |
| 8 | 20 | 20 | 313 | 78 | 0.250 | 273 | 486 | 1.554 | 5.745 | 0.288 | 0.750 |
| 9 | 13 | 15 | 234 | 63 | 0.267 | 203 | 282 | 1.205 | 5.457 | 0.310 | 0.733 |
| 10 | 18 | 11 | 172 | 47 | 0.273 | 148 | 134 | 0.780 | 5.147 | 0.318 | 0.727 |
| 11 | 8 | 8 | 125 | 78 | 0.625 | 86 | 48 | 0.385 | 4.828 | 0.981 | 0.375 |
| 12 | 3 | 3 | 47 | 16 | 0.333 | 39 | 9 | 0.192 | 3.847 | 0.405 | 0.667 |
| 13 | 2 | 2 | 31 | 3.442 |
| 龄级 Age stage |
Ax | ax | lx | dx | qx | Lx | Tx | ex | lnlx | Kx | Sx |
| 1 | 39 | 238 | 1000 | 160 | 0.160 | 920 | 3217 | 3.217 | 6.908 | 0.174 | 0.840 |
| 2 | 101 | 200 | 840 | 143 | 0.170 | 769 | 2448 | 2.913 | 6.734 | 0.186 | 0.830 |
| 3 | 131 | 166 | 697 | 134 | 0.193 | 630 | 1818 | 2.606 | 6.547 | 0.214 | 0.807 |
| 4 | 134 | 134 | 563 | 105 | 0.187 | 511 | 1307 | 2.322 | 6.333 | 0.206 | 0.813 |
| 5 | 109 | 109 | 458 | 101 | 0.220 | 408 | 900 | 1.965 | 6.127 | 0.249 | 0.780 |
| 6 | 85 | 85 | 357 | 105 | 0.294 | 305 | 595 | 1.666 | 5.878 | 0.348 | 0.706 |
| 7 | 60 | 60 | 252 | 88 | 0.350 | 208 | 387 | 1.536 | 5.530 | 0.431 | 0.650 |
| 8 | 39 | 39 | 164 | 29 | 0.179 | 149 | 238 | 1.452 | 5.099 | 0.198 | 0.821 |
| 9 | 31 | 32 | 134 | 42 | 0.313 | 113 | 125 | 0.926 | 4.901 | 0.375 | 0.688 |
| 10 | 21 | 22 | 92 | 50 | 0.545 | 67 | 57 | 0.620 | 4.527 | 0.788 | 0.455 |
| 11 | 10 | 10 | 42 | 21 | 0.500 | 32 | 26 | 0.614 | 3.738 | 0.693 | 0.500 |
| 12 | 5 | 5 | 21 | 8 | 0.400 | 17 | 9 | 0.428 | 3.045 | 0.511 | 0.600 |
| 13 | 3 | 3 | 13 | 6 | 2.534 |
在不同种群中, 岷江冷杉幼苗存活曲线的趋势基本一致(图 2), 但林窗幼苗因基数较大而更早遭遇到竞争的负面影响。对存活曲线进行拟合的结果表明(表 6), 幂模型和指数模型均能极显著(P < 0.001)地揭示林窗、林下以及总体岷江冷杉幼苗种群的动态;从回归系数R2、F检验值以及赤池信息量准则(AIC)来看, 指数模型的拟合效果要优于幂模型。说明林窗、林下以及总体岷江冷杉幼苗种群的存活曲线更趋于Deevey-Ⅱ型, 呈对角线型;对林窗和林下种群指数模型系数进行F检验, 系数在不同种群间无显著差异(F=0.7633, P=0.6690)。

|
| 图 2 岷江冷杉幼苗种群存活曲线 Fig. 2 Survival curve of A. faxoniana seedlings population |
| 种群 Type of population |
拟合模型 | R2 | F | AIC | 存活曲线类型 Type of survival curve |
| 林窗In canopy gaps | y=1145.51x-0.80 | 0.8187 | 58.36 | 136.29 | Deevey-Ⅱ |
| y=1382.49e-0.26x | 0.9761 | 497.85 | 109.45 | ||
| 林下Under forest canopy | y=1148.37x-0.74 | 0.8134 | 67.33 | 135.56 | Deevey-Ⅱ |
| y=1337.76e-0.24x | 0.9809 | 705.45 | 105.94 | ||
| 总体All stands | y=1161.20x-0.61 | 0.7792 | 75.96 | 136.87 | Deevey-Ⅱ |
| y=1275.57e-0.18x | 0.9696 | 585.93 | 111.10 |
不同生境中岷江冷杉幼苗种群的死亡率曲线(qx)和消失率曲线(Kx)变化趋势基本一致(图 3), 均呈随龄级增大而逐渐增大的趋势, 但数值、增幅及其波动性又各有差别。林窗种群的死亡率和消失率在5龄级前增幅不大, 6、7龄级增幅较大, 8龄级的值降为0, 9龄级时又增至最大值(0.789和1.558), 而后又降为0, 在12龄级时又有反弹。林下种群的死亡率和在消失率在11龄级前都呈缓慢增加的趋势, 11龄级时达到峰值0.625和0.981。在7龄级前, 林下种群的死亡率和消失率均小于林窗种群和总体种群。总体种群的死亡率和消失率变化趋势是林窗种群和林下种群的综合反映, 数值上更接近于林窗种群的值, 特别是9龄级前。

|
| 图 3 岷江冷杉幼苗种群的死亡率和消失率 Fig. 3 Mortality and vanish rate curve of A. faxoniana seedlings population |
生存分析的结果见图 4所示, 随着龄级的增大, 岷江冷杉幼苗种群的生存率(S(i))、死亡密度(f(i))呈下降趋势, 而累积死亡率(F(i))和危险率(λ(i))呈增加趋势。生存率在下降过程中(图 4), 7龄级前的降幅大于之后降幅;林窗种群的生存率较低, 林下的较高, 最大差值可达0.203(7龄级)。林窗种群的累计死亡率最高、总体的次之、林下的最小, 且表现出前期增幅大、后期增幅小的特点(图 4)。死亡密度在下降过程中出现波动, 如林窗种群在7龄级和9龄级、林下种群在4龄级和11龄级时;林窗种群的死亡密度在1—3龄级高于林下种群, 此后多低于林下种群。危险率以林窗种群高、林下种群低, 在随龄级变化过程中, 生长后期其单调上升的增幅在减小(图 4)。总体种群的4个函数值均介于林窗种群和林下种群的值之间, 但更接近于林窗种群。

|
| 图 4 岷江冷杉幼苗种群的生存率、累计死亡率、死亡密度和危险率曲线 Fig. 4 Survival rate, cumulative mortality rate, mortality density and hazard rate functional curve of A. faxoniana seedlings population |
岷江冷杉幼苗种群相邻龄级结构动态量化指数(Vn)在1龄级最小, 随着龄级的增加逐渐增大, 从4龄级开始增为正数, 此后又随龄级增加呈先增大后减小的趋势(图 5), 但这种趋势在林窗、林下以及总体种群中的表现各有差异。在林窗种群, V8、V10、V11依次比V7、V9和V10小, V3和V11为0(图 5), 最大值V9为83.33%。在林下种群, V6、V9和V12比前1龄级的有减小, V9为0, 峰值V11为62.50%, (图 5);在总体中, 最大值为V10(52.38%, 图 5)。

|
| 图 5 岷江冷杉幼苗种群龄级结构动态量化指数 Fig. 5 Dynamic index of population size structure of A.faxoniana seedlings population |
在不考虑外界环境干扰时, 岷江冷杉幼苗种群年龄结构的动态指数以林窗种群的最高(0.1209), 林下种群最小(0.0895), 总体的介于前两者之间(0.1059, 图 6)。对随机干扰的敏感性指数P极大和受随机干扰时的种群年龄结构指数Vpi′均以林窗种群最高、林下种群次之、总体种群的最小。

|
| 图 6 岷江冷杉幼苗种群动态的陈晓德量化指数 Fig. 6 ChenXiaode quantitative index of A. faxoniana seedlings population |
对种群动态的谱分析表明:幼苗种群更新动态是通过岷江冷杉不同龄级的株数分布波动表现的。图 7可看出, 岷江冷杉幼苗天然更新波动性明显, 基波(A1)最大, 林窗种群、林下种群和总体种群的A1值分别为1.624、0.778和1.205。基波表现基本周期的波动, 其周期长度为种群本身所固有, 由种群波动特性所决定。除受基波影响外, 岷江冷杉幼苗种群的小周期波动在不同种群中表现各异, 林窗种群的A5、A6谐波有波动, 分别对应9龄级和11龄级;林下种群仅A4谐波有轻微波动, 未发现其他谐波明显波动。

|
| 图 7 岷江冷杉幼苗种群动态的谱分析 Fig. 7 Spectral analysis of the population dynamic of A. faxoniana seedlings |
幼苗阶段被认为是森林更新过程中最重要和敏感的阶段[6, 31]。在岷江冷杉的天然更新过程中, 幼苗的补充数量及存活情况决定其种群的更新成败[14, 16]。王朗自然保护区内岷江冷杉-箭竹林内岷江冷杉幼苗储量为4.27×103株/hm2, 较卧龙自然保护区同一林型的幼苗储量丰富[15, 32];年龄结构金字塔呈中间大两头小的纺锤型(图 1), 表现为增长型的种群;采用陈晓德的方法[27]量化的幼苗种群数量化动态指数为正值(图 6), 进一步肯定岷江冷杉幼苗为增长型种群[2, 27]。可见, 在王朗自然保护区, 储量丰富的更新幼苗和增长型的种群动态能保障岷江冷杉-箭竹林的天然更新。
对种群生命表和存活曲线的分析可以反映种群现实状况、展现种群对环境的适应机制[1, 4, 33]。本研究中岷江冷杉幼苗种群的Deevey-Ⅱ型的存活曲线(表 6)明显不同于程伟等[16]、康东伟等[17]、谭留夷等[18]、曹广侠等[24]发现的岷江冷杉种群Deevey-Ⅲ型的存活曲线。Deevey-Ⅲ型曲线的种群具有早期死亡率较高, 一旦活到某一年龄, 死亡率就降低的特点[2, 4], 这些Deevey-Ⅲ型的生活曲线均反映出岷江冷杉幼苗期高死亡率的事实[16-18, 24]。而就岷江冷杉幼苗的存活曲线而言, 本研究结果仅与李媛等[20]报道的低密度箭竹环境中的相似, 与中、高密度箭竹环境中的相异。可见, 种群的存活曲线并非一类种群或某一生长阶段所固有[6, 20], 可能取决于外界因素的影响。
岷江冷杉幼苗的死亡率(图 3)、消失率(图 3)、累计死亡率(图 4)和危险率(图 4)也呈现出随苗龄的增大而增加的趋势, 生存率(图 4)和死亡密度(图 4)却随苗龄的增大而下降。这是由于岷江冷杉幼苗更新的集群效应明显、更新幼苗以聚集分布为主[32, 34], 随着个体的长大, 个体间的竞争增强, 自疏作用更强烈, 出现死亡率较高的现象[26]。进入8龄级, 表征种群动态的参数均发生明显的波动, 说明岷江冷杉幼苗已处于最关键和敏感的时期[24], 当不能适应环境条件的变化时, 种群的增长型将改变[27]。到10龄级后, 存活的个体数已较少, 自疏效应减缓, 但个体一旦死亡又将引起各项指数的明显变化, 造成种群的波动。经过高死亡率的幼苗阶段进入幼树阶段后, 岷江冷杉幼苗种群的生命期望将明显提高[20], 种群会呈长时期稳定的状态[24]。谱分析表明岷江冷杉种群的数量动态变化明显受整个生命周期生物学特征的控制[28-30], 由于所调查幼苗种群的年龄还不够长(最大仅龄级13), 仅幼苗阶段的种群动态尚不足以揭示岷江冷杉整个生长周期的固有波动[28-30]。
4.2 林窗影响岷江冷杉的林窗更新特色明显[13-14], 林窗的发生可以提高其种子的发芽率、降低虫害侵染率[12, 14], 倒木腐朽还能提供较多营养, 促进幼苗的扎根和生长等[13, 15]。在本研究中, 林窗的发生扩大了幼苗种群的规模。尽管林窗面积仅占样地面积的20.67%, 但补充的幼苗数量占总体种群规模的62.5%。相对于林下, 林窗可以获得和贮存更多的太阳辐射能[9], 使土壤温度、倒木腐朽状况、灌草密度、苔藓厚度等微环境更为复杂多变[8, 13], 林窗幼苗在适应多变环境的过程中表现出更强的敏感性和增长型(图 6), 抵御外界随机事件干扰的能力也更优秀, 且在经受随机事件干扰后增长型的种群较林下突出。
林窗内幼苗密度可达1.29×104株/hm2, 约为林下幼苗密度的6.45倍, 因而对资源的竞争更为强烈[35], 随龄级增加, 密度制约效应比林下种群更明显, 其累计死亡率和危险率要高于林内种群(图 4)。林窗内种群的死亡率、消失率和生存函数在7龄级后就出现明显的波动, 而林下种群的波动则相对较晚, 说明林窗内种群比林下种群更早地经受着环境的筛选。而从相邻龄级间个体数量变化指数也表明林窗内种群在8龄级、林下种群在9龄级时为负增长种群(图 5)。此外, 谱分析的结果也表明除受整个生命周期生物学特性的控制外, 林窗内岷江冷杉幼苗的小周期波动还受其他因素影响[13, 29]。
无论是林下还是林窗环境, 岷江冷杉幼苗的发生尚取决于其它因素(如箭竹、杜鹃、苔藓等)的制约程度[14, 34], 尤其箭竹密度对岷江冷杉种群更新阻碍最为明显[13, 19, 21]。即便是单位面积上岷江冷杉的落种甚多、且以林窗内的特多, 由于箭竹的密集丛生, 仍无幼苗的发生[14];但李媛等[20, 32]也指出, 密集的箭竹会严重抑制岷江冷杉幼苗的定居, 而小密度的箭竹可能促进岷江冷杉幼苗的更新。在本研究中缺苞箭竹密度可达9546株/hm2, 多呈块状分布, 对岷江冷杉幼苗发生的制约也表现在箭竹密集生长的地段。
本研究采用多种方法分析了林窗对川西亚高山岷江冷杉幼苗种群存活过程的影响, 年龄结构金字塔、存活曲线均未发现林窗内和林下幼苗种群存活过程的明显差别;而死亡率曲线、消失率曲线、生存函数均表明林窗对岷江冷杉幼苗存活过程的影响主要表现在中期阶段;种群动态数量化分析表明林窗内的岷江冷杉幼苗具有更强的抵御外界随机干扰的能力和增长潜力;谱分析则表明岷江冷杉幼苗的动态变化受整个生命周期生物学特征的控制, 林窗内种群波动还受其他因素影响。可见, 仅使用种群年龄结构、静态生命表、存活曲线和生存函数来比较林窗内和林下岷江冷杉幼苗种群的生存过程还尚有不足, 而增加数量化动态特征分析才能更好地反映不同种群增长型的差异。综合多种分析方法可以看出, 林窗尽管未改变岷江冷杉幼苗种群的年龄结构和存活曲线、还提高了幼苗的死亡率和危险率;但由于扩大了幼苗的数量规模、提高了种群抵御外界随机干扰的能力, 使种群的增长型更为突出;因而, 在岷江冷杉-箭竹林的天然更新过程中应重视林窗的作用。此外, 本研究关注了株高1.0 m内岷江冷杉幼苗的种群动态, 能否保障幼苗安全过渡到幼树、小树的阶段, 其种群波动特征尚需持续关注。
| [1] | Williams C E, Johnson W C. Age structure and the maintenance of Pinus pungens in pine-oak forests of southwestern Virginia. American Midland Naturalist , 1990 (1) : 124:130–141. |
| [2] | 向悟生, 王斌, 丁涛, 黄俞淞, 农重刚, 刘晟源, 李先琨. 喀斯特季节性雨林蚬木种群结构和数量动态. 生态学杂志 , 2013, 32 (4) : 825–831. |
| [3] | Bierzychudek P. Plant biodiversity and population dynamics. Ecology and the Environment , 2014, 8 : 29–65. |
| [4] | Hett J M, Loucks O L. Age structure models of balsam fir and eastern hemlock. Journal of Ecology , 1976, 64 (3) : 1029–1044. DOI:10.2307/2258822 |
| [5] | 申仕康, 马海英, 王跃华, 王博轶, 申国柱. 濒危植物猪血木(Euryodendron excelsum H.T.Chang)自然种群结构及动态. 生态学报 , 2008, 5 : 2404–2412. |
| [6] | 张婕, 上官铁梁, 段毅豪, 郭微, 刘卫华, 郭东罡. 灵空山辽东栎种群年龄结构与动态. 应用生态学报 , 2014, 25 (11) : 3125–3130. |
| [7] | Carthy J M. Gap dynamics of forest trees:A review with particular attention to boreal forests. Environmental Reviews , 2001, 9 (1) : 1–59. DOI:10.1139/a00-012 |
| [8] | 谭辉, 朱教君, 康宏樟, 胡理乐. 林窗干扰研究. 生态学杂志 , 2007, 26 (4) : 587–594. |
| [9] | 张远彬, 王开运, 鲜骏仁. 岷江冷杉林林窗小气候及其对不同龄级岷江冷杉幼苗生长的影响. 植物生态学报 , 2006, 30 (6) : 941–946. |
| [10] | 韩文娟, 袁晓青, 张文辉. 油松人工林林窗对幼苗天然更新的影响. 应用生态学报 , 2012, 23 (11) : 2940–2948. |
| [11] | 鲜骏仁, 胡庭兴, 张远彬, 王开运. 林窗对川西亚高山岷江冷杉幼苗生物量及其分配格局的影响. 应用生态学报 , 2007, 18 (4) : 721–727. |
| [12] | 刘庆. 林窗对长苞冷杉自然更新幼苗存活和生长的影响. 植物生态学报 , 2004, 28 (2) : 204–209. |
| [13] | Kang W, Tian C, Kang D W, Wang M J, Li Y X, Wang X R, Li J Q. Effects of gap microsites and bamboo on Abies faxoniana regeneration in a subalpine forest, China. Journal of Plant Interactions , 2015, 10 (1) : 59–64. DOI:10.1080/17429145.2015.1015059 |
| [14] | 杨玉坡, 叶兆庆, 钱国禧. 西南高山地区冷杉、云杉林冠下天然更新的初步观察. 林业科学 , 1956 (4) : 337–354. |
| [15] | 马姜明, 刘世荣, 史作民, 张远东, 缪宁. 川西亚高山暗针叶林恢复过程中岷江冷杉天然更新状况及其影响因子. 植物生态学报 , 2009, 33 (4) : 646–657. |
| [16] | 程伟, 吴宁, 罗鹏. 岷江上游林线附近岷江冷杉种群的生存分析. 植物生态学报 , 2005, 29 (3) : 349–353. |
| [17] | 康东伟, 刘夏明, 谭留夷, 康文. 四川省王朗自然保护区岷江冷杉种群结构特征研究. 河北林业科技 , 2010 (3) : 15–17. |
| [18] | 谭留夷, 赵志江, 康东伟, 康文, 李俊清. 王朗自然保护区岷江冷杉林(Abies faxoniana)更新及其种群结构分析. 四川林业科技 , 2011, 32 (1) : 31–37. |
| [19] | Taylor AH, Qin Z S. Regeneration patterns in old-growth Abies-Betula forests in the Wolong Natural Reserve, Sichuan, China. Journal of Ecology , 1988, 76 (4) : 1204–1218. DOI:10.2307/2260643 |
| [20] | 李媛, 陶建平, 王永健, 余小红, 席一. 暗针叶林下华西箭竹(Fargesia nitida)对岷江冷杉(Abies faxoniana)幼龄植株种群动态的影响. 生态学报 , 2007, 27 (3) : 1041–1049. |
| [21] | Taylor AH, Qin Z S. Tree regeneration after bamboo die-back in Chinese Abies-Betula forests. Journal of Vegetation Science , 1992, 3 (2) : 253–260. DOI:10.2307/3235687 |
| [22] | 王开运. 川西亚高山森林群落生态系统过程. 成都: 四川科学技术出版社, 200453–55. |
| [23] | 解婷婷, 苏培玺, 周紫鹃, 张海娜, 李善家. 荒漠绿洲过渡带沙拐枣种群结构及动态特征. 生态学报 , 2014, 34 (15) : 4272–4279. |
| [24] | 曹广侠, 林璋德, 张联敏. 甘南地区紫果云杉、岷江泠杉生命表. 生态学报 , 1991, 11 (3) : 286–288. |
| [25] | 谢宗强, 陈伟烈, 路鹏, 胡东. 濒危植物银杉的种群统计与年龄结构. 生态学报 , 1999, 19 (4) : 523–528. |
| [26] | 杨凤翔, 王顺庆, 徐海根, 李邦庆. 生存分析理论及其在研究生命表中的应用. 生态学报 , 1991, 11 (2) : 153–158. |
| [27] | 陈晓德. 植物种群与群落结构动态量化分析方法研究. 生态学报 , 1998, 18 (2) : 214–217. |
| [28] | 伍业钢, 韩进轩. 阔叶红松林红松种群动态的谱分析. 生态学杂志 , 1988, 7 (1) : 19–23. |
| [29] | 周资行, 焦健, 李毅, 李永兵, 张世虎. 腾格里沙漠南缘唐古特白刺无性系种群分株年龄结构及其动态. 林业科学 , 2013, 49 (12) : 1–9. |
| [30] | 洪伟, 王新功, 吴承祯, 何东进, 廖成章, 程煜, 封磊. 濒危植物南方红豆杉种群生命表及谱分析. 应用生态学报 , 2004, 15 (6) : 1109–1112. |
| [31] | Comita L S, Uriarte M, Thompson J, Jonckheere I, Canham C D, Zimmerman J K. Abiotic and biotic drivers of seedling survival in a hurricane-impacted tropical forest. Journal of Ecology , 2009, 97 (6) : 1346–1359. DOI:10.1111/jec.2009.97.issue-6 |
| [32] | 李媛, 陶建平, 王永健, 余小红, 席一. 亚高山暗针叶林林缘华西箭竹对岷江冷杉幼苗更新的影响. 植物生态学报 , 2007, 31 (2) : 283–290. |
| [33] | Taylor AH, Qin Z S, Liu J. Tree regeneration in an Abiesfaxoniana forest after bamboo dieback, Wang Lang Natural Reserve, China. Canadian Journal of Forest Research , 1995, 25 (12) : 2034–2039. DOI:10.1139/x95-220 |
| [34] | 王微, 陶建平, 胡凯, 李宗峰, 宋利霞. 华西箭竹对岷江冷杉林主要乔木树种幼苗结构及分布格局的影响. 林业科学 , 2007, 43 (1) : 1–7. |
| [35] | 黎磊, 周道玮. 红葱种群地上和地下构件的密度制约调节. 植物生态学报 , 2011, 35 (3) : 284–293. |