 2016, Vol. 36
2016, Vol. 36文章信息
- 杨惠滨, 国庆喜
- YANG Huibin, GUO Qingxi.
- 地形与竞争因子对红松胸径与年龄关系的影响
- Influence of topography and competitive factors on the relationship between DBH and age of Korean pine
- 生态学报[J]. 2016, 36(20): 6487-6495
- Acta Ecologica Sinica[J]. 2016, 36(20): 6487-6495
- http://dx.doi.org/10.5846/stxb201503090457
-
文章历史
- 收稿日期: 2015-03-09
- 网络出版日期: 2016-01-22





林分的年龄结构一直是森林生态学研究的重要内容之一。它是林木更新过程长短和更新速度快慢的反映[1-2]。红松(Pinus koraiensis)作为东北东部地带性顶极植被-阔叶红松林的建群种, 了解其年龄结构对于研究其在气候变化下生物群落演变、形成与维持机制, 制订合理的经营方案, 以及提高林分生产力等, 都有一定的实际意义。但是由于空间分布复杂和发育阶段各异等的影响, 天然林中树木年龄的测定难度很大[3], 并容易对其造成伤害, 因此对年龄的确定一般采用间接的推测方法。许多研究表明林木的直径生长与年龄呈正相关, 并可建立一定的生长模型[4-6]。通过建立树龄与胸径的回归关系, 能够简化测定方法, 大大提高工作效率。在20世纪80年代, 李克志[7]、孙玉军[8]等利用伊春林区红松解析木数据, 对红松的胸径、树高生长过程以及各生长阶段状态作了系统的研究。同一时期, 邵国凡[9]、蒋伊尹[10]则利用人工红松林解析木数据建立了胸径等生长的数学模型。近年来关于红松胸径与年龄关系的研究集中在长白山地区[1, 3], 并且考虑到了竞争因子在红松胸径与年龄关系中的影响[11-13]。目前对林木胸径和年龄的相互关系研究, 均从单因子出发, 建立简单的一元回归统计模型。但由于数据难以获得, 迄今为止在这方面开展的研究很少, 多数集中于红松小径木。从以往的研究和野外调查实践都可以看出, 相同径级的林木, 其年龄存在一定的差异, 究其原因, 除了本身的生物学特性, 还有林木周围的生长环境、周围林木的相对空间位置以及相互之间的距离等因素[3]。但对这些影响因素的了解仍十分模糊, 更缺乏影响因子的定量研究。有鉴于此, 本文通过搜集黑龙江小兴安岭的树龄在100a以上的成熟红松解析木数据进行分析, 利用解析木数据中红松生长的立地条件以及周围邻近木的信息, 探讨竞争、地形等有关因子对胸径与年龄之间关系的影响, 为阔叶红松林年龄结构的研究提供理论参考。
1 研究区概况小兴安岭位于黑龙江省东北部, 地理位置大约在46°28′—49°21′N, 127°42′—130°14′E之间。该区域属北温带大陆季风气候区, 四季分明, 最冷月1月平均气温-20—-25℃, 最低温度可达-40℃, 最热月7月平均气温19—21℃左右, 最高温度可达31℃, 极端最高气温为35℃, 全年平均气温-1—1℃;年降水量为500—700 mm;土壤类型为典型暗棕壤。小兴安岭在中国植被区划上属于温带针阔混交林区域内的北部地区, 原生地带性植被是红松为主的温带阔叶红松林, 是我国主要林区之一, 拥有丰富的珍贵用材树种, 木材蓄积量大。木本植物种类多样, 乔木树种主要包括红松(Pinus koraiensis)、紫椴(Tilia amurensis)、枫桦(Betula costata)、白桦(Betula platyphylla)、臭冷杉(Abies nephrolepis)、兴安落叶松(Larix gmelini)、蒙古栎(Quercus mongolica)、裂叶榆(Ulmus laciniata)、红皮云杉(Picea koraiensis)、鱼鳞云杉(Picea jezoensis)、水曲柳(Fraxinus mandshurica)、黄菠萝(Phellodendron amurense)、胡桃楸(Juglans mandshurica)、色木槭(Acer mono)和大青杨(Populus ussuriensis)等, 灌木主要包括暴马丁香(Syringa reticulata)、毛榛子(Corylus mandshurica)、瘤枝卫矛(Euonymus pauciflorus)、和东北鼠李(Rhamnus davurica)等。
2 数据来源及处理数据来自黑龙江省小兴安岭林区各地红松解析木数据资料且全部为林分平均木, 搜集的地区其南北纬度范围在46°56′—48°06′N之间, 北自丰林国家级自然保护区, 南至朗乡、铁力林业局其主要分布如下:五营、红星、汤旺河、朗乡、美溪、翠峦、南岔、铁力、新清、乌伊岭等林业局。整理每株红松解析木的胸径、年龄、生长状况、周围邻近木的胸径以及与解析木的距离, 海拔、坡度、坡向、坡位等地形因子的数据信息。所有的解析木都为天然林红松针阔叶混交林, 个体海拔主要分布在300—700 m之间共计156株, 周围邻近木共计566株, 其中红松占总株数的44.9%, 云冷杉占29.8%, 其他阔叶树占25.3%。解析木与周围邻近木生长状况及解析木胸径与年龄分布散点图分别见表 1和图 1。
| 指标 Index |
解析木 Analysis wood |
竞争木 Competitive wood |
解析木与竞争木距离统计量 Distance statistics of analysis wood and Competitive wood |
||||
| 年龄/a Age |
胸径/cm DBH |
树高/m Height |
年龄/a Age |
胸径/cm DBH |
树高/m Height |
||
| 最大值Maximum value | 296 | 66 | 31.6 | 300 | 81.3 | 29 | 8.0 |
| 最小值Minimum value | 100 | 3.0 | 3.13 | 12.0 | 6.0 | 4.0 | 0.5 |
| 均值mean value | 160 | 37.9 | 23.1 | 90 | 27.88 | 18 | 4.7 |

|
| 图 1 解析木胸径与年龄关系散点图 Fig. 1 Scatter diagram of the relatoinship between the DBH and age of analysis wood |
林木间的竞争对林木生长的影响是很难量化的[14], 学者们通常采用竞争指标来度量林木所遭受的竞争强弱, 竞争指标可分为与距离有关的竞争指标和与距离无关的竞争指标[15], 一般认为与距离有关的竞争指标能更好的反应竞争关系。为探讨每株解析木受到的竞争影响, 采用了Hegyi简单竞争指数[16], 它包括了树木大小、距离和空间分布等多方面的信息, 并且简单直观, 测算容易。Hegyi简单竞争指数是我国研究林木种群竞争关系最多的与距离有关的竞争指数。表达式为[17]:
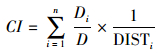
|
(1) |
式中, CI为对象木的简单竞争指数, 对象木指计算竞争指标时所针对的树木;D为对象木的胸径;Dj为竞争木i的胸径;DISTi为对象木与竞争木i的距离;n为竞争木的数量。
关于竞争木的距离范围界定, 国内有采用处于对象木5、6 m或8 m邻体半径范围内的林木作为竞争木[18-19], 也有采用与对象木最近的4株竞争木计算竞争指数[20]。考虑到竞争木树冠半径一般为2—4 m, 因此为了包含更多的竞争信息, 本文将邻体半径设置为8 m, 以尽量选取更多竞争木。参考有关研究报道[12], 如果邻体半径内林木胸径小于对象木胸径, 则不作为竞争木。
2.2 大树断面积竞争指标的改进大树断面积(BAL)为大于对象木胸径的所有林木的断面积之和[11, 21]。目前其在国际上应用广泛, 是最好的竞争指标之一[13, 22-23], 国内则研究较少, 是一种与距离无关的竞争指标。Busing认为, 某株林木周围邻体对其遮蔽和拥挤效应的大小, 均与它们之间距离的远近有关, 距离越远的邻体对这株林木生长的影响越小;反之则影响越大[24]。因此采用林分大树断面积作为竞争指标时, 可以只计算对象木周围邻体树木的大树断面积。有的研究根据树木周围邻体半径内其他林木的生物量来计算该树木所受到的竞争[25]。本文据此对BAL加以改进, 采用邻体半径内所有大于对象木胸径的林木胸高断面积的距离加权之和作为对象木所受到的竞争压力WBAL。

|
(2) |

|
(3) |
式中, Ai为邻体半径内第i株大于对象木胸径的林木个体的胸高断面积;PAi为对象木与其第i株邻体之间的距离加权系数取值范围为[0, 1]。当邻体与对象木距离等于0时, PA=1, 即邻体对于对象木的遮蔽和拥挤效应达到最大;当邻体距离等于邻体半径时, PA=0, 邻体对对象木生长无影响;加权系数PA的计算公式是根据图 2中角度与距离的关系所提出, 从图 2中可以看出, 邻体C和C′与中心树木O的距离越近, 其夹角AOB越大, 距离越远, 夹角越小。当邻体距离等于邻体半径时, 夹角等于0;当邻体与中心树木在一起时, 夹角等于180度按弧度制则等于π。THEi为一树木与其第i个邻体之间的夹角, 单位为弧度。

|
| 图 2 中心树木与其邻体间距离与夹角THE之间的关系 Fig. 2 The relationship between the distance and intersection angle of the center tree and its neighbous OC、OC′为邻体与中心树木之间的距离, 这一角度可以由下式计算, 式中DISTi为一树木与其邻体之间的距离;RADUS为邻体半径 |
目前, 国内外常用的竞争指标包括与距离有关的竞争指标和与距离无关的竞争指标两种, 每个竞争指标都有其各自的优势, 大树断面积(BAL)竞争指标的改进使两类计算方式相结合, 能更加有效的描述对象木所受的竞争压力。
2.3 回归方程的建立应用SPSS 19.0软件, 以林木年龄为因变量, 以胸径、竞争指标CI或WBAL、地形等因子为自变量建立回归方程。采用逐步回归分析法选取方程中所需的自变量。以F概率作为引入和剔除变量的判据, 当一个变量的双侧检验的显著性水平≤0.05时, 该变量被引入方程;当显著性水平≥0.10时, 该变量被剔除。为消除共线性, 方差膨胀因子(VIF)大于5的自变量也被排除在外[26]。
3 结果与分析 3.1 红松胸径与年龄的关系胸径随年龄的生长一般呈S型曲线[9], 林木一般在幼年时期胸径生长较慢, 而在中期, 尤其是在150a以后生长量大大提高[5], 直到生长后期又将呈现出生长缓慢趋势。反过来说, 本文利用胸径信息估算红松年龄大小, 考虑到生物学意义, 其总的拟合曲线应呈现出快—慢—快的变化趋势。又考虑到本文所选取的数据均为年龄在100a以上的近成熟红松, 变化趋势应表现为先慢后快, 所以用指数模型进行拟合更符合其生物学意义。并采用线性回归模型作为对比分析, 红松个体胸径与年龄的相关性分析结果表明Pearson相关系数为0.562, 表明对于所搜集的所有解析木, 二者之间线性相关程度并不高。随着年龄增加, 红松的胸径随之增大, 二者必然存在着紧密的相关性。然而由于各种影响因素的存在, 如竞争、地形、气候、甚至红松自身的生长节律等都会降低胸径与年龄之间的相关程度。
采用指数、线性两类模型, 以年龄为因变量, 胸径为自变量进行拟合(表 2)。从拟合结果看出, 在胸径与年龄变量均取对数的前提下, 指数模型决定系数达到0.507。即对于所搜集的解析木资料来说, 胸径变量通过指数模型对年龄的解释程度可以达到50.7%。而蒋伊尹对黑龙江省东部林区红松人工林胸径生长所做的分析表明, 胸径生长曲线用幂函数模型拟合比较理想, 模型的相关系数基本都在0.99以上[10]。胡云云等[3]对吉林省汪清林业局红松所做的胸径生长方程, 其幂函数模型决定系数也达到了0.97以上, 但所涉及的红松个体均为小径木。尽管上述报道是以胸径为因变量, 以年龄为自变量建立的回归方程, 但仍然说明对于本研究所涉及的天然林红松个体, 其胸径与年龄关系更加复杂, 受到的影响也更多。
| 序号 No. |
模型名称 Model name |
回归方程 Regression equation |
修正R2 Adjusted R2 |
| 1 | 线性模型 | lnA=1.834+0.884lnD | 0.473 |
| 2 | 指数模型 | lnA=2.478e0.195lnD | 0.507 |
| A:年龄, D:胸径 | |||
那么是否由于所搜集的解析木数据分布空间较大而影响了胸径年龄回归方程的拟合效果?比如研究地区南北相差约1个纬度, 年均温相差1.7℃, 可能会对红松胸径与年龄关系产生影响。我们挑选包含解析木数量最多(共计63个)的伊春地区五营林业局, 对其解析木进行胸径与年龄(均取对数)关系的曲线拟合, 其结果以指数模型拟合的决定系数只有0.402。说明即使消除了纬度变化因子, 红松胸径-年龄之间的拟合效果也并没有得到改善。
3.2 竞争因子对红松胸径与年龄关系的影响林木在生长过程中会受到种内或者种间竞争的影响。赵俊卉等人在对长白山主要针叶树种胸径和树高随年龄的变异系数与竞争因子的关系研究表明, 竞争是引起胸径变动的主要因子, 推断如果将竞争因子添加到传统的生长方程将会大幅度提高模型的精度[13]。因此本研究分别将CI、WBAL等竞争指标加入胸径-年龄回归方程, 探讨竞争因子对红松胸径与年龄关系的影响。以年龄为因变量, 以胸径、竞争因子为自变量, 采用多元逐步回归模型进行年龄的预测(表 3)。
| 序号 No. |
回归方程 Regression equation |
修正R2 Adjusted R2 |
| 1 | lnA=1.132+1.011lnD+0.184CL | 0.542 |
| 2 | lnA=1.704+0.899lnD+1.134WBAL | 0.583 |
从表 3可以看出, 不同的竞争指标对红松胸径-年龄回归关系的影响是不同的。引入简单竞争指数CI之后, 回归模型的修正决定系数增加为0.542;而引入WBAL指标的结果则使胸径-年龄拟合效果有所改善达到0.583。但是这种改善效果也是很有限的, 没有大幅度提高拟合精度, 至少对于所搜集的解析木数据是这样。因此进一步探讨其他因子的影响是十分必要的。
3.3 地形因子对红松胸径与年龄关系的影响在前面所做的多元线性回归模型基础上, 进一步加入坡度、海拔等地形因子。结果表明坡度与海拔因子均被剔除, 说明对于所搜集的全部解析木, 坡度与海拔因子没有显示出影响。本文拟将坡向与坡位因子分别加以限定, 继续开展相关探讨。
3.3.1 坡向对红松胸径与年龄关系的影响参照有关文献[27], 将方位角0°—180°范围内的山坡定义为东坡向;90°—270°为南坡向;180°—360°为西坡向;270°—90°为北坡向。各坡向范围之间存在一定的重叠。
各坡向红松解析木胸径-年龄最佳回归方程拟合效果依次为南坡>西坡>东坡>北坡。在各个坡向中, 处于南坡的回归方程以指数模型拟合效果最佳, 达到了0.594, 而以北坡的拟合效果最差。从这一现象来看, 不同坡向对红松胸径-年龄之间关系的影响存在差异, 尤其在北坡受到的影响最大。
分别用各个坡向的数据建立胸径与年龄、竞争指标、坡度、海拔等之间的多元回归方程(表 5), 并与表 4中相对应坡向的单因子回归方程决定系数进行比较。
| 序号 No. |
坡向 aspect |
样本数 Sample number |
回归方程 Regression equation |
修正R2 Adjusted R2 |
| 1 | 东 | 34 | lnA=0.618lnD+2.830 | 0.187 |
| 2 | 南 | 76 | lnA=1.176+1.044lnD+0.190CI | 0.588 |
| lnA=1.617+0.913lnD+1.350WBAL | 0.648 | |||
| lnA=0.948lnD+1.578WBAL+0.001ALT | 0.673 | |||
| 3 | 西 | 53 | A=4.421D+59.012CI-19.309 | 0.523 |
| A=3.382D+265.318WBAL+13.692 | 0.542 | |||
| 4 | 北 | 46 | A=23.777+3.208D+36.222CI | 0.337 |
| A=31.876+2.888D+253.629WBAL | 0.465 |
| 序号 No. |
坡向 aspect |
样本数 Sample number |
回归方程 Regression equation |
修正R2 Adjusted R2 |
| 1 | 东 | 34 | lnA=3.26e0.121lnD | 0.214 |
| 2 | 南 | 76 | lnA=2.331e0.121lnD | 0.594 |
| 3 | 西 | 53 | lnA=2.866e0.856lnD | 0.273 |
| 4 | 北 | 46 | lnA=3.65e0.089lnD | 0.110 |
由表 5可知, 采用东坡向数据建立的方程, 竞争因子、海拔等变量皆被剔除, 说明对分布于东坡的红松解析木, 在其胸径-年龄关系中加进入竞争因子和海拔变量对提高拟合优度没有贡献。在南坡, 基于表 4中胸径-年龄最佳线性拟合曲线(指数模型)效果(R2=0.594), 加入WBAL与海拔指标的回归方程, 其拟合效果有所改善(修正R2=0.673)。在各坡向中, 加入竞争指标之后对提高回归方程拟合优度最明显的是北坡与西坡, 例如加入WBAL指标, 决定系数分别提高了0.355和0.269。说明竞争因子对北坡与西坡的红松解析木胸径-年龄关系有较大的影响。
3.3.2 坡位对红松胸径与年龄关系的影响从表 6可以看出分布于中坡位的红松解析木胸径-年龄回归方程其修正系数达到了0.717。而上坡位与下坡位的拟合效果较差。
| 序号 No. |
坡位 Position of slope |
样本数 Sample number |
模型类型 Model type |
回归方程 Regression equation |
修正R2 Adjusted R2 |
| 1 | 上 | 39 | 指数模型 | lnA=3.141e0.131lnD | 0.214 |
| 2 | 中 | 69 | 指数模型 | lnA=2.183e0.231lnD | 0.717 |
| 3 | 下 | 48 | 指数模型 | lnA=3.202e0.124lnD | 0.183 |
分别用各个坡位的数据建立年龄与胸径、竞争指标、坡度、海拔等之间的多元回归方程(表 7)。并与表 6中相对应坡位的单因子回归方程决定系数进行比较。
| 序号 No. |
坡位 Position of slope |
样本数 Sample number |
回归方程 Regression equation |
修正R2 Adjusted R2 |
| 1 | 上 | 39 | lnA=0.75lnD+0.163CI+2.283 | 0.261 |
| lnA=0.698lnD+0.765WBAL+2.473 | 0.264 | |||
| 2 | 中 | 69 | lnA=1.136lnD+1.310CI+0.878 | 0.732 |
| lnA=0.990lnD+1.310WBAL+1.37 | 0.775 | |||
| lnA=1.011lnD+1.173WBAL+0.001ALT+1.028 | 0.788 | |||
| 3 | 下 | 48 | lnA=0.794lnD+0.251CI+2.073 | 0.297 |
| lnA=0.727lnD+1.238WBAL+2.312 | 0.345 |
在各坡位, 采用CI与WBAL竞争指标的回归方程, 其拟合优度都得到了改善, 其中以采用WBAL指标的效果更好一些。在中坡位, 加入WBAL与海拔因子之后, 回归方程修正决定系数达到了0.788, 是拟合效果最好的一组方程。而加入WBAL对改善下坡位的胸径-年龄回归方程效果最为明显, 这也说明下坡位的红松解析木的胸径生长受到竞争因子的影响最大。
本文将分布于南坡向与中坡位的解析木筛选出来, 共计39株, 所做的胸径-年龄回归方程以指数函数模型拟合效果最佳, 方程式为lnA=2.115e0.239lnD, 决定系数达到0.809, 进一步建立年龄与胸径、竞争指标、坡度、海拔等之间的多元回归方程(表 8)。结果表明采用WBAL、胸径变量对年龄的拟合效果有所改善;进一步引入海拔因子, 修正决定系数提高到了0.858。
| 序号 No. |
回归方程 Regression equation |
修正R2 Adjusted R2 |
| 1 | lnA=1.009lnD+1.137WBAL+1.297 | 0.842 |
| 2 | lnA=1.028lnD+1.324WBAL+0.001ALT+0.923 | 0.858 |
影响红松个体生长的因素是多方面的。立地条件、生长节律、周围林木竞争等等, 都会造成个体之间差异, 导致胸径-年龄关系复杂化, 增加依靠胸径推断年龄的困难。本文利用所搜集的小兴安岭红松解析木资料, 对影响成熟红松个体胸径-年龄关系的因素作了初步探讨。针对所搜集的全部解析木估计其胸径与年龄的最佳拟合曲线。结果表明在胸径与年龄变量均取对数的前提下, 以指数模型进行拟合, 其决定系数也只有0.507。侯向阳认为, 红松林中林木年轮径向生长受其周围其他林木的空间竞争影响是巨大的。这种生存空间竞争对林木生长的影响往往超过任何其他因素的影响[28]。但是在引入竞争因子后, 并没有像预计的那样大幅度提高对年龄的预测精度, 尽管采用距离加权的大树断面积指标(WBAL)比简单竞争指数(CI)更好地体现了竞争因子的影响。李克志对小兴安岭红松生长的分析表明, 年龄在100a以后, 林木已摆脱被压状态[7]。孙玉军将小兴安岭红松的生长划分为4个阶段, 在90a以下为树木生长缓慢阶段, 100—130a为生长加速阶段, 在此阶段, 竞争减弱, 胸径生长明显加快[8]。而本研究所涉及的红松解析木个体, 年龄都在100a以上, 胸径基本都大于20 cm, 所受到的竞争应该是比较小的。以往有研究表明, 红松胸径小于25 cm时, 所承受的竞争压力较大;胸径大于25 cm以后, 竞争指数变化不大, 并维持较低的水平[13]。
为了探讨地形因子对成熟红松胸径-年龄关系的影响, 分别按照坡向与坡位对红松解析木数据进行分类, 建立胸径-年龄拟合方程。结果表明南坡向或中坡位的红松解析木, 其胸径-年龄单因子回归方程的拟合优度大幅提高, 而南坡向与中坡位的地形因子组合使红松胸径-年龄单因子回归方程的拟合效果达到了最优, 引入竞争因子与海拔之后进一步改善了拟合效果。在其他坡向或坡位建立的方程虽然拟合效果较差, 但在引入竞争因子之后, 则明显提高了对年龄的预测精度。由以上模型得出红松胸径生长是十分复杂的过程, 在不同坡向、坡位其生长规律有所不同, 在南坡向、中坡位其生长规律较为一致, 这可能与其充足光照和相对较高的温度有关。而在西、北坡向其生长差异较大, 但是引入竞争指标后其拟合精度虽然不是很理想但是却有了较大的提高, 可能由于某些生长因子缺乏如光、热量, 而不能满足所有个体的生长需求从而凸显竞争优势的重要性, 使得量化的竞争指标对其拟合优度有了大为促进作用。
通过本文分析可知, 对于所搜集的小兴安岭地区红松解析木个体, 坡向、坡位、海拔, 以及竞争指标等都影响着红松胸径与年龄的关系, 其中对红松胸径-年龄关系影响最明显的是地形因子中的坡向与坡位, 在南坡向与中坡位的红松个体, 其胸径与年龄之间表现出了最佳的拟合效果。而加入竞争因子却没有大幅度提高红松胸径-年龄回归方程的拟合优度, 并且竞争因子作用的发挥可能受到地形因子尤其是坡向与坡位的制约, 坡向、坡位能够控制太阳辐射和降水的空间再分配, 营造局部小气候[29], 但二者又是通过哪些具体机制来影响红松胸径与年龄的关系还有待进一步研究。并且现有的分析方法只能体现地上部分的竞争, 并不能体现地下部分根系的竞争情况和林木对光、温度的利用情况[30]。
| [1] | 代力民, 孙伟中, 邓红兵, 肖以华. 长白山北坡椴树阔叶红松林群落主要树种的年龄结构研究. 林业科学 , 2002, 38 (3) : 73–77. |
| [2] | Zhang Y, Drobyshev I, Gao L S, Zhao X H, Bergeron Y. Disturbance and regeneration dynamics of a mixed Korean pine dominated forest on Changbai Mountain, North-Eastern China. Dendrochronologia , 2014, 32 (1) : 21–31. DOI:10.1016/j.dendro.2013.06.003 |
| [3] | 胡云云, 亢新刚, 赵俊卉. 长白山地区天然林林木年龄与胸径的变动关系. 东北林业大学学报 , 2009, 37 (11) : 38–42. |
| [4] | 徐德应. 林分和林木生长预测模型. 世界林业研究 , 1988 (3) : 42–49. |
| [5] | 邓坤枚, 邵彬, 李飞. 长白山北坡云冷杉林胸径、树高结构及其生长规律的分析. 资源科学 , 1999, 21 (1) : 77–84. |
| [6] | Wimberly M C, Bare B B. Distance-dependent and distance-independent models of Douglas-fir and western hemlock basal area growth following silvicultural treatment. Forest Ecology and Management , 1996, 89 (1/3) : 1–11. |
| [7] | 李克志. 红松生长过程的研究. 林业科学 , 1983, 19 (2) : 126–136. |
| [8] | 孙玉军, 刘吉春, 陈霖生, 张鹏, 刘传照, 戴兴安, 李海奎. 天然红松单木生长阶段的多元定量化研究. 东北林业大学学报 , 1993, 21 (6) : 1–5. |
| [9] | 邵国凡. 红松人工林单木生长模型的研究. 东北林业大学学报 , 1985, 13 (3) : 38–46. |
| [10] | 蒋伊尹. 红松人工林生长与生长模型. 东北林学院学报 , 1985, 13 (2) : 5–15. |
| [11] | 黄新峰, 亢新刚, 孙玲, 冯启祥, 姚景春. 红松单木断面积生长模型. 西北林学院学报 , 2011, 26 (3) : 143–146. |
| [12] | 刘红润, 李凤日. 红松天然林种内和种间竞争关系的研究. 植物研究 , 2010, 30 (4) : 479–484. |
| [13] | 赵俊卉, 亢新刚, 张慧东, 刘燕. 长白山主要针叶树种胸径和树高变异系数与竞争因子的关系. 应用生态学报 , 2009, 20 (8) : 1832–1837. |
| [14] | Holmes M J, Reed D D. Competition indices for mixed species Northern Hardwoods. Forest Science , 1991, 37 (5) : 1338–1349. |
| [15] | Avery T E, Burkhart H E. Forest Measurements. New York: McGraw-Hill, 19831–331. |
| [16] | Radtke P J, Westfall J A, Burkhart H E. Conditioning a distance-dependent competition index to indicate the onset of inter-tree competition. Forest Ecology and Management , 2003, 175 (1/3) : 17–30. |
| [17] | 沈琛琛, 雷相东, 王褔有, 马武, 沈剑波. 金苍林场蒙古栎天然中龄林竞争关系研究. 林业科学研究 , 2012, 25 (3) : 339–345. |
| [18] | 张思玉, 郑世群. 笔架山常绿阔叶林优势种群种内种间竞争的数量研究. 林业科学 , 2001, 7 (1) : 185–188. |
| [19] | 邹春静, 王庆礼, 韩士杰. 长白山暗针叶林建群种竞争关系的研究. 应用与环境生物学报 , 2001, 7 (2) : 101–105. |
| [20] | 李建军, 李际平, 刘素青, 赵春燕, 张宏伟, 冯湘兰. 基于Hegyi改进模型的红树林空间结构竞争分析. 中南林业科技大学学报 , 2010, 30 (12) : 23–27. |
| [21] | Wykoff W R. A basal area increment model for individual conifers in the northern Rocky Mountains. Forest Science , 1990, 36 (4) : 1077–1104. |
| [22] | Stage A R, Ledermann T. Effects of competitor spacing in a new class of individual-tree indices of competition:semi-distance-independent indices computed for Bitterlich versus fixed-area plots. Canadian Journal of Forest Research , 2008, 38 (4) : 890–898. DOI:10.1139/X07-192 |
| [23] | Roberts S D, Harrington C A. Individual tree growth response to variable-density thinning in coastal Pacific Northwest forests. Forest Ecology and Management , 2008, 255 (7) : 2771–2781. DOI:10.1016/j.foreco.2008.01.043 |
| [24] | Busing R T. A spatial model of forest dynamics. Vegetatio , 1991, 92 : 167–179. |
| [25] | 葛剑平. 森林生态学建模与仿真. 哈尔滨: 东北林业大学出版社, 1996. |
| [26] | 雷相东, 张则路, 陈晓光. 长白落叶松等几个树种冠幅预测模型的研究. 北京林业大学学报 , 2006, 28 (6) : 75–79. |
| [27] | 童跃伟, 项文化, 王正文, DurkaW, FischerM. 地形、邻株植物及自身大小对红楠幼树生长与存活的影响. 生物多样性 , 2013, 21 (3) : 269–277. |
| [28] | 侯向阳, 韩进轩, 阳含熙. 长白山红松阔叶林林冠木竞争生长及林冠空隙动态研究. 生态学报 , 2000, 20 (1) : 68–72. |
| [29] | Cantón Y, Del Barrio G, Solé-Benet A, Lázaro R. Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain. Catena , 2004, 55 (3) : 341–365. DOI:10.1016/S0341-8162(03)00108-5 |
| [30] | Bugmann H. A review of forest gap models. Climatic Change , 2001, 51 (3/4) : 259–305. DOI:10.1023/A:1012525626267 |