 2016, Vol. 36
2016, Vol. 36文章信息
- 马青青, 刘建军, 余鸽, 刘伟, 马亦生
- MA Qingqing, LIU Jianjun, YU Ge, LIU Wei, MA Yisheng.
- 佛坪国家级自然保护区秦岭箭竹克隆结构的SSR分析
- Clonal structure of a Fargesia qinlingensis population inferred using simple sequence repeat fingerprints in Foping National Nature Reserve
- 生态学报[J]. 2016, 36(20): 6496-6505
- Acta Ecologica Sinica[J]. 2016, 36(20): 6496-6505
- http://dx.doi.org/10.5846/stxb201503070443
-
文章历史
- 收稿日期: 2015-03-07
- 网络出版日期: 2016-01-22





2. 西北农林科技大学风景园林艺术学院, 杨凌 712100;
3. 陕西佛坪国家级自然保护区管理局, 佛坪 723400
2. College of Landscape Architecture and Arts, Northwest Agriculture & Forestry University, Yangling 712100, China;
3. Shaanxi Foping National Nature Reserve, Foping 723400, China
在被子植物中, 克隆繁殖是一种极其普遍的繁殖方式[1], 导致母本与亲本产生相同的基因, 对植物的遗传结构有重要影响[2]。克隆植物的克隆大小及空间分布结构已经成为克隆植物生态学和群体遗传学研究的重要组成部分[3]。居群克隆多样性和遗传结构的空间分布是物种长期进化过程中自然选择等多种因素共同作用的结果, 因此对克隆多样性和克隆结构的研究有助于了解各种进化因素的作用[4]以及克隆植物的定居、侵殖和演替机理。空间自相关分析是研究小尺度范围内种群遗传结构的有效工具[5], 已被广泛的运用于种群遗传结构领域中[6]。
秦岭箭竹(Fargesia qinlingensis)属禾本科竹亚科箭竹属, 具有地下茎合轴丛生的克隆生长方式[7], 是一种典型的克隆植物[8]。秦岭箭竹耐寒性强, 较耐干旱贫瘠土壤, 在海拔2000 m上下有大面积纯林, 有的绵延数公里至数十公里[9], 对水土保持、气候调节和维护竹林野生动物的生物多样性等方面有重要的生态服务功能[10]。已有研究表明竹子以基株为单位开花, 基株交错或镶嵌生长, 开花时间不同导致一片区域开花时间可以持续好几年[11-12], 在没有区分无性系分株间的遗传结构时, 不能确定竹林是否同时开花。秦岭箭竹是大熊猫夏季食物的主要来源[13], 开花周期一般为50a[14], 其大面积开花或死亡会直接造成大熊猫食物短缺。例如, 1970年岷山缺苞箭竹和糙花箭竹的开花事件就导致了138只大熊猫的死亡[15]。
为准确了解竹类的遗传结构, 分子标记法已经被广泛地运用于竹类克隆多样性和克隆结构的研究中。如Suyama等通过AFLP分析了高山地带日本矮竹Sasa senanensis的基株分布, 发现在10hm2范围内至少有22个克隆, 最大的克隆约有300 m, 陡坡、岩石、溪流等微环境对克隆多样性和克隆结构都有较大影响[16]。Isagi等利用AFLP分子标记法对同一个居群毛竹开花前和开花后的克隆结构进行了研究, 发现起源于上次开花的不同的基株都有各自的开花时间, 开花现象和克隆结构之间的关系以及小面积开花都可能影响野生毛竹居群克隆的镶嵌性结构[17]。Franklin等通过SSR分子标记对澳大利亚丛生竹Bambusa arnhemica进行研究, 推断出:野外观察到的一些野生的丛生竹类, 一丛不只有一种克隆, 而且生长过于密集的分株会导致竞争的加剧[18]。Ma等利用AFLP分子标记法发现四川卧龙保护区内小于30岁龄级和大于70岁龄级的两个年龄段的冷箭竹Bashania fangiana都是多克隆的, 最大的克隆可达30m, 克隆多样性水平很高[19]。
目前对于秦岭箭竹的研究主要集中在分类与分布、生物量、种子萌芽、幼苗生长、叶片光谱特性变化、无性系构件形态等方面[7, 20-22], 利用分子标记法对其克隆多样性和克隆结构的研究还未见报导。SSR分子标记法是生物学里最有效的遗传标记法之一, 是由1—6个核苷酸为单位组成的多次串联重复的DNA序列, 这种序列广泛地存在于真核生物中[23]。与其他分子标记法相比, SSR分子标记法表现出诸如PCR筛选过程简单、能够提供相对丰富的潜在共显性变异等优点[24], 被认为是更适合用于遗传多样性分析的分子标记法[25], 并且已经被运用于多种植物克隆多样性和克隆结构研究中[26-28]。因此本研究利用SSR分子标记, 旨在初步探讨佛坪国家级自然保护区内秦岭箭竹的克隆多样性和克隆结构及其形成原因, 以期为该种开花特性、高山地区生态环境维护和大熊猫的保护提供重要依据。
1 材料与方法 1.1 实验材料样地设在佛坪国家级自然保护区野猪荡的山脊上, 地理位置E107°50′40.4″、N33°41′50.2″, 海拔2530 m左右, 此处秦岭箭竹密集地分布于巴山冷杉-牛皮桦混交林下。秦岭箭竹地下茎合轴丛生, 依据McClure以丛为单位分单株取样的方法采集样品[29]。为避免对某一丛竹子重复取样, 根据野外观察到的每丛竹子大小确定取样单株的最小间距为1 m。取样分两步进行:第一步, 设立5 m×5 m的两个样方a1、a2, 以1 m×1 m为单位在交叉点上共取样46个;第二步, 为了得到清晰的克隆结构, 设立两个样方A(30 m×10 m)、B(40 m×40 m), 根据a1、a2实验结果最终确定以5 m×5 m为单位在交叉点上取样。两个样地由长约120 m宽约40 m的裸露岩石带阻隔, 其中样地A由a1、a2扩大得到, 共计取样61个, 样地B共取样81个(图 1)。所有个体均经GPS定位以做小尺度空间自相关分析。将所取新鲜幼嫩叶片放入装有变色硅胶的塑封袋中于-80℃保存。

|
| 图 1 秦岭箭竹两个样方的克隆空间分布 Fig. 1 Clonal structure in two plots of Fargesia qinlingensis inferred from Simple Sequence Repeat (SSR) fingerprints |
用植物基因组DNA提取试剂盒(北京天根生化科技有限公司)并按试剂盒提供的流程提取秦岭箭竹的总DNA。用1%的琼脂糖凝胶电泳检测DNA片段大小, 用紫外分光光度计检测DNA的纯度和浓度。所提取的DNA用ddH2O稀释至50 ng/μL, 于-20℃保存。
1.2.2 SSR引物设计和合成从NCBI上下载箭竹属的DNA序列信息(http://www.ncbi.nlm.nih.gov/), 利用SSRHunter[30]软件对所得序列进行SSR搜索, 选取二、三、四、五和六核苷酸重复基序长度大于等于12 bp的序列, 运用OLIGO软件设计引物, 引物长度为18—25 bp, 扩增产物预期片段为130—400 bp。同时选取文献中近缘物种的SSR引物(引物1和7)[31-32], 共合成引物51对(生工生物工程(上海)股份有限公司)。
1.2.3 SSR引物筛选和PCR扩增随机选取4个样品对上述合成的51对引物进行首轮筛选, PCR产物用3%的琼脂糖凝胶检测。从剩下的样品中随机选取10个进一步筛选扩增条带在目的片段内的引物, 用8%的非变性聚丙烯酰胺凝胶检测PCR产物, 共筛选出7对引物(表 1)。
| 引物编号 Primer |
登录号 GenBank Accesion no. |
重复基序 Repeat motif |
5′-3′端引物序列 5′-3 Primer sequence |
退火温度 Annealing temperature/℃ |
扩增产物 Production size/ bp |
| 1 | GQ267718 | (GA)11 | F:GAGTCCTCCGCTCTGTGCAAT R:AATTAATCACCCCATCTCCAAGC |
58 | 166—232 |
| 2 | GQ281375 | (AG)9 | F:TTACACCGCGAGCCGTCCATC R:AATGCACTGTCATGATCCGCAAC |
59 | 169—226 |
| 3 | GQ281370 | (TG)9 | F:AGAAAGATAGGGATAGTGATTGTGT R:TCCGAGGTGAAAAAGGAGGAACT |
57 | 182—201 |
| 4 | GQ281366 | (AG)9 | F:GAGAGTGCCAAGAGACCACTG R:CTCACACACGCACACAACACAAA |
58 | 135—180 |
| 5 | GQ281356 | (GA)11 | F:ACGTGTTAGCTCGGTTCGACTG R:TTCTCTTCCTCCCTACATCGTCT |
59 | 168—226 |
| 6 | JN131934 | (TA)6 | F:ACGGAACGGTACAATATATGCT R:TCTTGCCTTCTATGATGGTGTC |
55 | 162—172 |
| 7 | EDO18306 | (TC)11 | F:GCCGTCCAAACGCTCCTT R:CACCCATCCATCTTATGCTAT |
52 | 257—279 |
上述筛选的条件如下:PCR反应体系为20 μL反应体系, 包括2 × PCR缓冲液、50 ng模板DNA、0.16 mmol/L dNTP、1.2 mmol/L Mg2+、0.2 μmol/L引物和1.0 U Taq DNA聚合酶。PCR反应在美国Bio-RAD公司生产的MyCycler Thermal Cycler #170—9701EDU型PCR仪上进行。PCR反应程序为, 94℃预变性5 min, 各引物最佳退火温度退火45 s, 72 ℃延伸45 s, 进行30个循环, 72 ℃延伸5 min后在4 ℃保存。将筛选出7对多态性好且稳定的引物在ABI3730XL9(Applied Biosystems, USA)毛细管电泳仪上检测所有样品, 用GeneMapper 4.0 (ABI, USA)软件读取扩增片段大小。
1.2.4 数据分析毛细管电泳得到的结果为扩增产物的bp值, 所有引物扩增bp值相同的样品为同一个基因型, 将原始bp数据转化为0、1矩阵。
运用下列指数对克隆多样性进行分析[2]:
(1) 平均克隆大小(N/G) 即所有样本中基因型相同的为同一基株, G是居群中基株总数, N是样本总数。
(2) 不同基因型比率(G/N) N为样本大小, G为基株总数。
(3) Simpson多样性指数(D) D=1-{[∑Ni(Ni-1)]/[N(N-1)]}
式中, Ni表示第i个基因型的总数, N表示样本大小。D从0到1变化, D为0表示整个居群是相同的基因型;D为1表示居群内样品的基因型各不相同。
(4) Fager指数(E)[33] 表示种群内不同基因型分布的均匀度。

|
式中, Dmin=[(G-1)(2N-G)]/[N(N-1)],
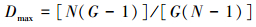
|
式中, E从0到1变化, E为0表示整个种群只有一个基因型或有一个基因型占据主要优势而其他基因型各包含一个样品;E为1表示种群内不同基因型有相同的样本。
用POPGENE v1.31软件[34]计算多态位点百分率(PPB)和Nei′s遗传距离。运用PowerMaker 3.25[35]对所有基因型进行UPGMA非加权算术平均聚类分析(Unweighted pair-group method analysis)。运用GenAlEx 6.4[36]软件计算秦岭箭竹所有个体和克隆遗传距离矩阵在相应距离等级下(本研究中距离等级间相隔4 m)的空间自相关系数r, 分析其小尺度克隆结构。
2 结果与分析 2.1 秦岭箭竹的克隆多样性利用筛选出的7对引物对秦岭箭竹的142个样品进行PCR扩增, 共检测到79个位点, 其中多态性位点77个, 多态位点百分率(PPB)为97.47%, 扩增片段大小为135—279 bp。供试的142个秦岭箭竹个体包含107个克隆(图 1), 基因型比率(G/N)为0.7535, Simpson多样性指数为0.9680, 平均克隆大小(N/G)为1.3271, Fager均匀度指数(E)为0.5109, 显示出较高的克隆多样性。
秦岭箭竹居群的克隆空间分布见图 1。A、B两个样地均由多克隆组成, 分别有30和77个克隆, 并且没有相同的基因型。在较小的取样尺度(a1、a2, 1 m×1 m)和较大的取样尺度下(5 m×5 m)分别有16个和91个克隆。克隆1由25个分株组成, 克隆9、30、43、102、106和107各由2个分株组成, 克隆12包括6个分株, 其余克隆都只有一个分株。秦岭箭竹同一克隆的分株排列紧密(如克隆1和12), 相邻分株属于同一克隆的可能性更大, 各克隆呈镶嵌性分布, 为密集型构型。
UPMGA对所有克隆聚类如图 2所示。各克隆之间聚类不明显, 表现出较高的遗传相似性, 总体上来自同一样地的克隆被聚为一类, 但样地A的克隆5、18、19和23与样地B的克隆71、39、75、96、107、33和104聚在一起, 起源更为相似。

|
| 图 2 107个基因型UPGMA聚类图 Fig. 2 UPGMA clustering based on dice genetic similarities calculated from SSR data between the 107 clones of Fargesia qinlingensis accessions 蓝色的数字代表来自样地A的克隆, 黑色的数字代表来自样地B的克隆 |
秦岭箭竹居群所有分株和克隆的空间自相关分析结果如图 3所示。当距离等级间隔为4 m时, 所有分株在空间距离小于44 m时存在显著的正相关空间遗传结构, 存在聚集的格局, 在56—116 m之间存在显著的负相关空间遗传结构, 而在大于120 m时不存在显著的空间遗传结构, 说明个体在56 m外个体差别较大, X-轴截距为52.280, 表示不规则克隆的平均大小, 空间自相关系数r的取值范围为0.084—0.626, 在4 m处个体最为相似, 表现出最大的正相关性(r=0.626)。所有克隆在空间距离小于36 m时表现出显著的正相关空间遗传结构, 在56—104 m之间存在显著的负相关空间遗传结构, 而在36—56 m之间和108 m以外不存在显著的空间遗传结构, 空间自相关系数r的取值范围分别为0.024—0.288。

|
| 图 3 秦岭箭竹居群所有个体和克隆空间自相关曲线图 Fig. 3 Correlograms showing the spatial autocorrelation coefficient r of all individuals and clones with Fargesia qinlingensis population r为空间自相关系数;U和L分别表示不存在显著性空间自相关遗传结构的95%置信区间的上限和下限 |
本研究中秦岭箭竹的Simpson多样性指数(D)为0.9680, 不同基因型比率(G/N)平均值为0.7535, 高于Ellstrand和Roose[2]总结的克隆植物的平均值(D=0.62, G/N=0.17, E=0.68)表现出较高的克隆多样性。Fager均匀度指数(E=0.5109)与之相比略低, 说明秦岭箭竹居群内基因型分布均匀度处于中等水平, 每个克隆包含的分株数不完全相同。取样策略(比如网格的规模和大小)和采样个体的选择都会影响秦岭箭竹克隆多样性水平。如Peng等对蔷薇科植物(Ploylepis reticulate)的克隆多样性研究发现较大取样尺度比较小取样尺度的克隆多样性更高, 克隆多样性受取样大小和空间范围影响[37]。
克隆多样性由基株的死亡和补充比率来确定[38], 在最初定居后的短时期内只有一些幼苗能够存活下来以维持居群的克隆多样性[39], 随之分株和基株间的竞争排斥或死亡会导致克隆多样性的降低[40], 如果没有种苗重复补充, 基株就会急剧减少, 最后导致几个大的克隆的出现[41]。秦岭箭竹无性繁殖期很长, 大约每50a开1次花结1次籽[14], 因此在建群后发生实生苗补充的机率很低, 此外, Ma等发现同为箭竹属的冷箭竹(Bashania fangiana)很可能是初始苗补充物种[19], 据此推测初始苗补充对秦岭箭竹的克隆多样性有极大贡献。
聚类分析表明来自同一样地的克隆先聚在一起, 但也有部分来自不同样地的克隆被聚为一类, 这可能是由于采样点位于山脊, 地势开阔平坦, 在初期种苗补充时, 有利于风力、动物等传播花粉或种子, 从而使这些克隆有了更近的起源。
Lovett Doust把克隆植物的生长型分为为游击型和密集型, 认为典型的密集型物种, 克隆镶嵌分布而单个克隆中的分株紧密聚合;典型的游击型物种, 克隆混合生长而单个克隆里的分株排列松散[42]。实验结果表明秦岭箭竹居群内不同克隆镶嵌分布, 在较小的取样尺度下(克隆1、12)单个克隆内的分株紧密聚合, 属于密集型空间分布格局。这可能是因为本实验恰好处于该地区箭竹人工造林较理想的生境, 当环境适宜、养分充足时, 秦岭箭竹人工林表现为节间变短, 隔离者长度减小, 分株增多[8], 因此分株呈现出密集型克隆结构。
微环境对克隆结构有很大影响。Suyama等对日本矮竹(Sasa senanensis)的克隆结构进行分析发现生长在地势平坦区域的克隆分布范围较广, 大多数生长在地势陡峭区域的克隆生长范围受到限制, 一个克隆只包含1个或2个分株[16]。Wang等发现交错的岩石阻碍了华西箭竹(Fargesia nitida)地下茎的延伸, 在贫瘠的土壤里无法形成较为密集的种群[43]。根据野外观测, 样地A边缘分布大片的裸露岩石, 分布过于密集的岩石限制了秦岭箭竹的生长空间, 种苗只能在较小的空间内进行无性繁殖, 地下茎很难延展, 无法形成较大克隆, 加之秦岭箭竹地下茎合轴丛生, 因而此处出现了多个克隆(克隆1、2、3、6、7、8、9、10、11), 而居群内部障碍物较少, 地下茎能有足够的生长空间, 克隆1和克隆12有较多分株。此外, 秦岭箭竹在较小的取样尺度下存在由单个分株组成的克隆, 这可能是因为在长寿命的克隆植物中, 体细胞突变是维持其遗传变异的持续来源[2], 可以使物种更好的适应环境变化。李钧敏等对小尺度范围内蛇莓(Duchesnea indica)克隆多样性的研究也得到类似结果[44]。在较大尺度下大多数个体基因型各不相同, 体现出取样尺度对克隆结构的影响。
物种遗传变异的空间分布格局与繁育系统、花粉和种子散布方式等生殖生物学特性以及种群生境异质性等有着密切联系[45-46]。空间自相关分析表明, 在36 m范围内, 可能是由于存在广泛的克隆繁殖, 分株比基株表现出更为显著的正相关空间遗传结构, 反映出秦岭箭竹呈密集型分布, 在其他克隆植物的小尺度克隆结构研究中也有类似结果[37, 47], 本研究以1 m×1 m为单位进行预实验, 以初步确定克隆大小, 结果发现最大克隆为5 m, 继而以此为单位扩大样地以较大的样本量验证秦岭箭竹的最大克隆能否超过5 m, 结果发现大多数克隆都在5 m范围内, 而空间自相关分析表明所有分株在4 m处表现出最大的空间自相关系数(r=0.626), 可能是因为无性繁殖可以导致短距离内正的空间自相关[48], 说明相距4 m内的个体最有可能属于同一克隆, 在以后的研究中可以以此为单位取样, 以便得到更清晰的克隆结构。在1 m×1 m和5 m×5 m的取样尺度下, 秦岭箭竹最大克隆分别包括25(克隆1)和2(克隆30、43、102、106、107)个分株, 可见单个克隆在较小的取样尺度下包含更多分株。正如Harada[49]的观点:在一定范围内较小的取样尺度更能清晰地阐明克隆结构, 而过大的取样尺度会错过小尺度的空间遗传结构。
竹子花粉较轻, 无粘性, 为风媒传粉植物[50], 空旷的地带有利于花粉和种子的长距离扩散。秦岭箭竹野猪荡居群位于山脊, 地势开阔, 有利于花粉和种子传播到更远区域, 但由于其开花结籽周期很长, 通过地下茎合轴丛生的方式进行克隆繁殖的生长习性占据了整个生命周期的绝大部分, 因此克隆繁殖在一定程度上限制了空间遗传结构的范围。
X-轴截距为52.280 m, 代表了秦岭箭竹不规则克隆的平均最小长度, 明显小于沈晓婷得出的毛竹X-轴截距平均值63.36 m[51]。这可能是因为毛竹“游击型”构型的分株之间的距离可以很大, 而秦岭箭竹“密集型”克隆结构的分株排列更为紧密, 各基株呈镶嵌性分布。综上所述, 秦岭箭竹的克隆多样性和克隆结构与初始苗补充、花粉散播方式和微环境差异有关。本文仅用SSR分子标记法初步探讨了秦岭箭竹的克隆多样性和克隆结构, 不同分子标记法和不同取样策略下的相关研究以及克隆结构的动态变化有待深入开展。
| [1] | Albert T, Raspé O, Jacquemart A L. Clonal structure in Vaccinium myrtillus L. revealed by RAPD and AFLP markers. International Journal of Plant Sciences , 2003, 164 (4) : 649–655. DOI:10.1086/375373 |
| [2] | Ellstrand N C, Roose M L. Patterns of genotypic diversity in clonal plant species. American Journal of Botany , 1987, 74 (1) : 123–131. DOI:10.2307/2444338 |
| [3] | 夏立群, 李建强, 李伟. 论克隆植物的遗传多样性. 植物学通报 , 2002, 19 (4) : 425–431. |
| [4] | Epperson B K. Recent advances in correlation studies of spatial patterns of genetic variation. Evolutionary Biology , 1993, 27 : 95–155. |
| [5] | Sokal R R, Oden N L, Thomson B A. Local spatial autocorrelation in biological variables. Biological Journal of the Linnean Society , 1998, 65 (1) : 41–62. DOI:10.1111/bij.1998.65.issue-1 |
| [6] | Epperson B K, Chung M G. Spatial genetic structure of allozyme polymorphisms within populations of Pinus Strobus (Pinaceae). American Journal of Botany , 2001, 88 (6) : 1006–1010. DOI:10.2307/2657081 |
| [7] | 李云, 任毅, 贾辉. 秦岭大熊猫主食竹的分类学研究(I). 西北植物学报 , 2003, 23 (1) : 127–129. |
| [8] | 王瑞, 周建云, 曹庆, 赵凯辉, 何景峰, 张文辉. 不同生境秦岭箭竹人工种群的生长特性研究. 西北农林科技大学学报:自然科学版 , 2011, 39 (5) : 110–118. |
| [9] | 陕西森林编辑委员会. 陕西森林. 西安: 中国林业出版社, 19891–263. |
| [10] | 李睿, 章笕, 章珠娥. 中国竹类植物生物多样性的价值及保护进展. 竹子研究汇刊 , 2003, 22 (4) : 7–12. |
| [11] | Miyazaki Y, Ohnishi N, Takafumi H, Hiura T. Genets of dwarf bamboo do not die after one flowering event:evidence from genetic structure and flowering pattern. Journal of Plant Research , 2009, 122 (5) : 523–528. DOI:10.1007/s10265-009-0241-9 |
| [12] | Kitamura K, Kawahara T. Clonal identification by microsatellite loci in sporadic flowering of a dwarf bamboo species, Sasa cernua. Journal of Plant Research , 2009, 122 (3) : 299–304. DOI:10.1007/s10265-009-0220-1 |
| [13] | Liu X H, Toxopeus A G, Skidmore A K, Shao X M, Dang G D, Wang T J, Prins H H T. Giant panda habitat selection in Foping nature reserve, China. Journal of Wildlife Management , 2005, 69 (4) : 1623–1632. DOI:10.2193/0022-541X(2005)69[1623:GPHSIF]2.0.CO;2 |
| [14] | Wang W, Franklin S B, Ouellette J R. Clonal regeneration of an arrow bamboo, Fargesia qinlingensis, following giant panda herbivory. Plant Ecology , 2007, 192 (1) : 97–106. DOI:10.1007/s11258-006-9229-x |
| [15] | 秦自生. 四川大熊猫的生态环境及主食竹种更新. 竹子研究汇刊 , 1985, 4 (1) : 1–10. |
| [16] |
Suyama Y, Obayashi K, Hayashi I.
Clonal structure in a dwarf bamboo (Sasa senanensis) population inferred from amplified fragment length |
| [17] | Isagi Y, Shimada K, Kushima H, Tanaka N, Nagao A, Ishikawa T, OnoDera H, Watanabe S. Clonal structure and flowering traits of a bamboo[Phyllostachys pubescens (Mazel) Ohwi] stand grown from a simultaneous flowering as revealed by AFLP analysis. Molecular Ecology , 2004, 13 (7) : 2017–2021. DOI:10.1111/mec.2004.13.issue-7 |
| [18] | Franklin D C, Kaneko S, Yamasaki N, Isagi Y. Short communication. Some wild bamboo clumps contain more than one genet. Australian Journal of Botany , 2008, 56 (5) : 433–436. DOI:10.1071/BT08026 |
| [19] | Ma Q Q, Song H X, Zhou S Q, Yang W Q, Li D S, Chen J S. Genetic structure in Dwarf Bamboo (Bashania fangiana) clonal populations with different genet ages. PLoS One , 2013, 8 (11) : e78784. DOI:10.1371/journal.pone.0078784 |
| [20] | 冯永辉, 冯鲁田, 雍严格, 党高弟, 任毅. 秦岭大熊猫主食竹的分类学研究(Ⅱ). 西北农林科技大学学报:自然科学版 , 2006, 36 (1) : 101–102. |
| [21] | 刘雪华, 吴燕. 大熊猫主食竹开花后叶片光谱特性的变化. 光谱学与光谱分析 , 2012, 32 (12) : 3341–3346. |
| [22] | 钟伟伟, 刘益军, 史冬梅. 大熊猫主食竹研究进展. 中国农学通报 , 2006, 22 (5) : 141–145. |
| [23] | Jurka J, Pethiyagoda C. Simple repetitive DNA sequences from primates:compilation and analysis. Journal Molecular Evolution , 1995, 40 (2) : 120–126. DOI:10.1007/BF00167107 |
| [24] | Zhao H S, Yang L, Peng Z H, Sun H Y, Yue X H, Lou Y F, Dong L L, Wang L L, Gao Z M. Developing genome-wide microsatellite markers of bamboo and their applications on molecular marker assisted taxonomy for accessions in the genusPhyllostachys. Scientific Reports , 2015, 5 : 8018–8018. DOI:10.1038/srep08018 |
| [25] | De Carvalho Moretzsohn M, Hopkins M S, Mitchell S E, Kresovich S, Valls J F M, Ferreira M E. Genetic diversity of peanut (Arachis hypogaea L.) and its wild relatives based on the analysis of hypervariable regions of the genome. BioMed Central Plant Biology , 2004, 4 : 11. |
| [26] | Araki K, Shimatani K, Ohara M. Dynamics of distribution and performance of ramets constructing genets:a demographic-genetic study in a clonal plant, Convallaria keiskei. Annals of Botany , 2009, 104 (1) : 71–79. DOI:10.1093/aob/mcp092 |
| [27] | 郑书星, 张建国, 段爱国, 何彩云, 保尔江, 王健. 额尔齐斯河流域银白杨克隆结构及多样性研究. 林业科学研究 , 2013, 26 (4) : 426–432. |
| [28] | Zalapa J E, Bougie T C, Bougie T A, Schlautman B J, Wiesman E, Guzman A, Fajardo D A, Steffan S, Smith T. Clonal diversity and genetic differentiation revealed by SSR markers in wild Vaccinium macrocarpon and Vaccinium oxycoccos. Annals of Applied Biology , 2015, 166 (2) : 196–207. DOI:10.1111/aab.2015.166.issue-2 |
| [29] | McClure F A. The Bamboos:A Fresh Perspective. Cambridge, USA:Harvard University Press , 1966 : 1–347. |
| [30] | 李强, 万建民. SSRHunter, 一个本地化的SSR位点搜索软件的开发. 遗传 , 2005, 27 (5) : 808–810. |
| [31] | Yu G, Liu J J, Ma Q Q, Ma Y S. Development polymorphic microsatellite primers for the giant panda's staple food bamboo Bashania fargesii and B. aristata (Poaceae:Bambusoideae). Conservation Genetics Resources , 2015, 7 (3) : 739–741. DOI:10.1007/s12686-015-0450-6 |
| [32] | 卢江杰, 吉永胜彦, 方伟, 汤定钦. 3种竹类植物杂种的分子鉴定. 林业科学 , 2009, 45 (3) : 29–34. |
| [33] | Fager E W. Diversity:a sampling study. The American Naturalist , 1972, 106 (949) : 293–310. DOI:10.1086/282772 |
| [34] | Yeh F C, Yang R C, Boyle T. POPGENE:The user friendly shareware or population genetic analysis. Edmonton, Canada:Molecular Biology and Biotechnology Center, University of Alberta., 1997. |
| [35] | Liu K J, Muse S V. PowerMarker:an integrated analysis environment for genetic marker analysis. Bioinformatics , 2005, 21 (9) : 2128–2129. DOI:10.1093/bioinformatics/bti282 |
| [36] | Peakall R, Smouse P E. GenALEx 6:genetic analysis in Excel. Population genetic software for teaching and research. Molecular Ecology Notes , 2006, 6 (1) : 288–295. DOI:10.1111/men.2006.6.issue-1 |
| [37] | Peng Y L, Macek P, Macková J, Romoleroux K, Hensen I. Clonal diversity and fine-scale genetic structure in a high Andean treeline population. Biotropica , 2015, 47 (1) : 59–65. DOI:10.1111/btp.2015.47.issue-1 |
| [38] | Verburg R, Maas J, During H J. Clonal diversity in differently-aged populations of the pseudo-annual clonal plantCircaea lutetiana L. Plant Biology , 2000, 2 (6) : 646–652. DOI:10.1055/s-2000-16633 |
| [39] | Soane I D, Watkinson A R. Clonal variation in populations of Ranunculus repens. New Phytologist , 1979, 82 (2) : 557–573. DOI:10.1111/nph.1979.82.issue-2 |
| [40] | Simmonds N W. Monocarpy, calendars and flowering cycles in Angiosperms. Kew Bulletin , 1980, 35 (2) : 235–245. DOI:10.2307/4114566 |
| [41] | Watkinson A R, Powell J C. Seedling recruitment and the maintenance of clonal diversity in plant populations:a computer simulation of Ranunculus repens. Journal of Ecology , 1993, 81 (4) : 707–717. DOI:10.2307/2261668 |
| [42] | Doust L L. Population dynamics and local specialization in a clonal perennial (Ranunculus repens):Ⅰ. The dynamics of ramets in contrasting habitats. Journal of Ecology , 1981, 69 (3) : 743–755. DOI:10.2307/2259633 |
| [43] | Wang Y J, Tao J P, Zhong Z C. Factors influencing the distribution and growth of dwarf bamboo, Fargesia nitida, in a subalpine forest in WoLong Nature Reserve, southwest China. Ecological Research , 2009, 24 (5) : 1013–1021. DOI:10.1007/s11284-008-0573-2 |
| [44] | 李钧敏, 金则新. 匍匐茎草本植物蛇莓小尺度克隆结构. 生态学报 , 2009, 29 (7) : 3540–3548. |
| [45] | Chung M G, Chung J M, Chung M Y, Epperson B K. Spatial distribution of allozyme polymorphisms following clonal and sexual reproduction in populations of Rhus javanica (Anacardiaceae). Heredity , 2000, 84 (2) : 178–185. DOI:10.1046/j.1365-2540.2000.00660.x |
| [46] | Marquardt P E, Epperson B K. Spatial and population genetic structure of microsatellites in white pine. Molecular Ecology , 2004, 13 (11) : 3305–3315. DOI:10.1111/j.1365-294X.2004.02341.x |
| [47] | Ohsako T. Clonal and spatial genetic structure within populations of a coastal plant, Carex kobomugi (Cypceaceae). American Journal Botany , 2010, 97 (3) : 458–470. DOI:10.3732/ajb.0900262 |
| [48] | Geburek T. Are genes randomly distributed over space in mature populations of sugar maple (Acer saccharum Marsh.)?. Annals of Botany, , 1993, 71 (3) : 217–222. DOI:10.1006/anbo.1993.1027 |
| [49] | Harada Y, Kawano S, Iwasa Y. Probability of clonal identity:inferring the relative success of sexual versus clonal reproduction from spatial genetic patterns. Journal of Ecology , 1997, 85 (5) : 951–600. |
| [50] | 林树艳, 石文文, 缪彬彬, 丁雨龙. 竹类植物生殖生物学研究进展. 世界竹藤通讯 , 2010, 8 (2) : 1–6. |
| [51] | 沈晓婷.毛竹小尺度遗传多样性及取样策略的初步研究[D].南京:南京林业大学, 2012. |