 2016, Vol. 36
2016, Vol. 36文章信息
- 王烨, 秦浩, 董刚, 秦晓娟, 赵小娜, 张峰
- WANG Ye, QIN Hao, DONG Gang, QIN Xiaojuan, ZHAO Xiaona, ZHANG Feng.
- 山西油松林分类学多样性
- Taxonomic diversity of Pinus tabuliformis communities in Shanxi
- 生态学报[J]. 2016, 36(20): 6520-6527
- Acta Ecologica Sinica[J]. 2016, 36(20): 6520-6527
- http://dx.doi.org/10.5846/stxb201502040284
-
文章历史
- 收稿日期: 2015-02-04
- 网络出版日期: 2016-01-22





2. 山西大学生命科学学院, 太原 030006
2. School of Life Science, Shanxi University, Taiyuan 030006, China
生物多样性是评价物种数量、结构类型[1]、分布格局[2]、时空变化的有力工具[3]。传统的生物多样性指数大都基于物种个体数及相关信息, 如Margalef丰富度指数、Pielou均匀度指数及Simpson多样性指数等, 这些指数对取样的时间、地点、样地大小与方法极其敏感[4], 同时也不能直接反映种间的分类等级关系。
分类等级关系是群落物种多样性的重要方面, 当两个群落物种丰富度相同时, 分类关系较远的群落比分类关系较近的有更大的多样性[5]。为了研究群落分类学关系的变化, Clarke等提出了考虑物种间的分类关系的指数[6], 主要包括分类多样性指数Δ、分类差异指数Δ*、平均分类学差异指数Δ+和分类学差异变异指数Λ+[7]。平均分类学差异指数Δ+和分类学差异变异指数Λ+是基于物种间权重, 根据分类等级的路径长度, 将群落分类学多样性特征量化的方法[8-9]。其平均值不依赖于取样性质和样本大小, 可用于大尺度的研究, 避免取样性质未知或不一致对取样数据的影响[10], 且易于计算[11]。Δ+反映群落种间亲缘关系[12], Δ+值越高表示物种间亲缘关系越远, 多样性越大;Λ+可以反映群落物种分类关系的均匀程度[13], Λ+值越大表明物种分布均一性越差。分类学多样性研究最初集中在动物区系和鱼类群落[6, 14]。如Leonard等[15]应用不同的分类指标评估英国海洋生物多样性;Ellingsen等[16]研究了挪威大陆架底栖生物的分类学多样性。近十余年来, 在植被生态研究中分类学多样性方法逐渐得到应用, 如李景文等[17]对海南主要热带森林植物的分类学多样性的研究;Da Silva等[18]对巴西中部稀树草原的研究;秦晓娟等[3]检验了分类学多样性对湿地植物群落的适用性;赵小娜等[19]对庞泉沟自然保护区植物群落分类学多样性进行研究, 并分析了分类学多样性指数与环境因子的关系。
油松(Pinus tabuliformis)是中国北方地区重要的用材和生态树种, 以油松为建群种形成的森林群落是华北山地优势植被类型之一, 山西是我国油松林分布的集中区之一, 具有广泛的代表性。本文在对山西油松群落进行野外调查的基础上, 运用分类学多样性指数(Δ+和Λ+)对山西油松群落的分类学多样性进行研究, 探讨山西油松林物种组成特征和分类学多样性及与其有关的物种进化关系、物种共存和群落构建等问题, 分析地形因子和群落因子与分类学多样性指数的相关性, 分析大尺度上影响物种分类学差异性的因子, 为群落构建等问题提供一定的理论依据;同时, 检验分类学多样性分析方法在森林群落生态学中的应用效果, 全面了解区域尺度上油松林的分类学多样性, 为山西油松林资源保护与管理提供科学依据。
1 研究区概况油松林在山西北起恒山, 至五台山折向西, 在岢岚、兴县以南分布极为广泛。以太行山的盂县、平定、和顺、左权、榆社、黎城、陵川, 太岳山的沁源、霍州、介休、灵石、洪洞, 吕梁山的交城、中阳、蒲县、乡宁, 五台山的繁峙、五台、代县, 恒山, 中条山的沁水、阳城、翼城[20-21], 黑茶山的岚县、兴县、方山,管涔山的宁武、五寨、岢岚等地分布较为集中。其中太岳山是我国油松林分布的代表性区域[22], 而大同天然油松很少成林[23]。
野外调查的油松林(山西)分布范围:110°45′—113°45′ E, 34°48′—38°57′ N, 海拔744—1867 m。研究区属温带大陆性季风气候, 全年日照时数2808 h, 无霜期120 d[24]。年平均气温为4—14 ℃, 1月平均温为-6.8 ℃, 7月平均温为23.6℃。年降水量为400—600 mm。
2 研究方法 2.1 野外调查2012年6月—2014年9月, 在山西共设置113个样地进行植被调查(图 1), 几乎遍及山西省油松林分布区。样地面积为20×30 m2, 记录乔木层树种名、高度、冠幅、多度和郁闭度等;将样地划分为6个10×10 m2的样方, 选取对角线的2个样方, 记录灌木层种名、多度、平均高和盖度;在6个样方中各选取1个1×1 m2的小样方, 记录草本层种名、平均高和盖度[25-26], 采集每个样方的表层土, 密封保存, 分析土壤属性。记录的环境信息包括经纬度、海拔、坡度、坡向、坡位和干扰情况等。

|
| 图 1 调查地点的基本信息 Fig. 1 Dsitribution of stands of Pinus tabulaeformis in Shanxi |
坡向采用等级制, 即阳坡(1)、半阳坡(2)、半阴坡(3)和阴坡(4), 数字越大, 表示水分条件越好。坡位同样采用等级制, 即下坡位(1)、中下坡位(2)、中坡(3)、中上坡(4)和上坡位(5);干扰情况可分为无干扰(4)、轻微(3)、中度(2)和重度(1), 数字越小, 表示人为干扰相对越强。将调查的环境因子划分为宏观地形因子(经度、纬度和海拔)、微地形因子(坡度、坡向和坡位)、土壤属性因子(全氮、全磷、pH和有机碳)和群落结构因子(胸径、乔木密度和群落物种数), 以便进行比较。
2.2 数据分析综合野外调查和历史资料整理山西油松林物种总名录。根据林奈分类系统, 将物种分类等级确定为门、纲、目、科、属、种6个等级。定义分类等级上最远的2个种路径长度为100(即最长路径)[6], 根据分类等级确定每一等级水平上的权重[27-28]:同属的不同种ωij=16.67[3];同科不同属的种ωij=33.33;同目不同科的种ωij=50.00;同纲不同目的种ωij=66.67;同门不同纲的种ωij=83.33;不同门的种ωij=100。
Clarke等[29]提出了计算分类学多样性测度的方法, 公式如下:
平均分类学差异指数

|
(1) |
分类学差异变异指数
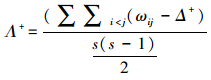
|
(2) |
式中, ωij为第i和第j个物种在分类等级中的路径长度, S为样方中出现的种数。
对各样地的多样性指数值与理论平均值进行统计性检验, 建立95%的置信漏斗曲线[5, 30]。由PRIMER6软件可求得Λ+与Δ+的值并完成置信漏斗的绘制。
用R软件完成Λ+和Δ+与地形因子、土壤属性和群落结构的Spearman秩相关分析。
3 结果与分析 3.1 群落结构与组成山西北部油松林多为人工油松纯林, 林下灌木较少, 草本植物优势种为长芒草(Stipa capillata)等。中西部吕梁山、太岳山和太行山油松林成熟林较少, 其中太岳山中幼龄林占优势;关帝林区的油松多为中幼龄林, 且多为纯林, 同时分布有少量的松、栎类混交林;太行山区幼龄林约占2/3, 其余大部分为中龄林, 常与栎类混交;灌木层主要有黄刺玫(Rosa xanthina)、美蔷薇(Rosa bella)、三裂绣线菊(Spiraea trilobata)、土庄绣线菊(Spiraea pubescens)、虎榛子(Ostryopsis davidiana)等;草本层以披针薹草(Carex lancifolia)为主。南部中条山幼龄林居多, 中龄林约占l/3, 常与白桦(Betula platyphylla)、辽东栋(Quercus wutaishanica)、元宝槭(Acer truncatum)、山杨(Populus davidiana)等混交, 灌木种类和数量较多, 常见的有胡枝子(Lespedeza bicolo)、黄刺玫(Rosa xanthina)、金银忍冬(Lonicera maackii)、荆条(Vitex negund var. heterophylla)、西北栒子(Cotoneaster multiflorus)、甘肃山楂(Crataegus kansuensis)、土庄绣线菊、黄栌(Cotinus coggygria var. coggygria)、暴马丁香(Syringa reticulata)等, 草本层主要以羊胡子草(Carex rigescens)、华北米蒿(Artemisia giraldii)等为主。
野外共记录357种植物, 隶属于3门4纲36目71科227属。3个门分别为被子植物门(包含2纲350种)、裸子植物门(包含1纲5种)和蕨类植物门(包含1纲2种)。被子植物中双子叶植物纲(30目)占优势地位。从目水平来看, 管状花目含科最多, 有6科。含属最多的为桔梗目(34属), 含种最多的为蔷薇目(71种)。植物科内种、属组成差异较大, 其中菊科(31属)、蔷薇科(18属)、豆科(15属)禾本科(15属)、唇形科(11属)等含属较多。菊科含种最多(50种), 占总种数的14.01%, 其次为豆科34种(9.52%)、蔷薇科33种(9.24%)和禾本科21种(5.88%)。含种较多的属为蒿属(Artemisia 11种)、堇菜属(Viola 9种)、委陵菜属(Potentilla 7种)、沙参属(Adenophora 7种)和薹草属(Carex 5种)等。
由此可见, 山西油松林群落大都属于中幼龄林, 林下的物种组成和分类单元相对比较丰富, 但主要以几个大的分类单元为主, 如在科水平上集中于菊科、蔷薇科、豆科和禾本科。
3.2 分类学多样性指数根据山西油松林的Δ+和Λ+理论平均值建立95%的置信漏斗曲线(图 2)。图 2中置信漏斗的上下置信限分别为相应种数在最大阶元和最小阶元的分类学多样性理论值。图 2Δ+中粗线是Δ+的理论平均值(约为53), 所有样地Δ+理论平均值与种数的变化基本独立。Δ+阈值在种数为4—7时明显减小, 之后随着种数的增加, 逐渐趋近于稳定, 与Δ+理论平均值平行。

|
| 图 2 Δ+和Λ+ 95%的置信漏斗曲线及各调查点在曲线中的位置 Fig. 2 Δ+ and Λ+ for the plant communities in 113sites, plotted against the number of species on the 95% confidence funnel |
Δ+值表示群落中所有种间路径长度的理论平均值, 置信漏斗范围内样地Δ+值越大, 表示样地内种间分类学多样性越大, 种间亲缘关系越远。Δ+值大都处于研究区Δ+的理论平均值之上, 表明所调查的样地内种间的分类学多样性比较丰富。其中位于置信漏斗上包迹线上方的样地占总样地数的11.5%, 包括样地S6(永济五老峰)、S78(晋源区天龙山)、S85(阳城庄沟)等, 这些样地的Δ+值显著高于区域的Δ+理论平均值(P < 0.05)。位于理论平均值下方的样地数远远小于其上方样地数, 仅有样地S10(运城盐湖区)、S12(安泽草峪岭)、S30(襄垣古韩)、S47(左权程家村)、S108(交城双家寨)等10个;位于下包迹线之外的样地并未出现。样地S78平均分类差异指数Δ+值(64.86)高于其余样地, 即样地S78种间亲缘关系较其他样地远。样地S12的Δ+值(47.23)最低, 平均每科含3.33种, 每属含3种, 表明其物种组成间的亲缘关系较近。
图 2置信漏斗的上下置信限分别为Λ+值理论的最大曲线与最小曲线, Λ+值可表征物种分类单元分布的均一性水平。置信漏斗范围内样地Λ+值越小, 表明油松群落间的分类地位关系程度越均匀。虚线是Λ+的理论平均值, 反映区域内物种分类等级间的差异变异情况。由图 2可以看出, Λ+理论平均值也几乎与种数变化无关, 基本保持在280左右。只有当种数很少时( < 5), Λ+的最大和最小曲线才会出现急剧变化;之后随着种数的增加逐渐趋于稳定, 且逐渐平行于Λ+理论平均值。
所有样地的Λ+值均位于理论平均值上方, 表明油松林种间的分类等级均匀性低于区域内的理论平均值。其中有25个样地位于置信漏斗最大值曲线上方, 分别为样地S5(平陆东郭)、S6、S12、S17(永和芝河)、S20(乡宁台头)等。这些样地的Λ+值显著高于区域内的理论平均值(P < 0.05), 表现出种间分类单元显著的不均匀性。样地S30分类差异变异指数Λ+(917.35)最高, 表明其物种的等级分布最不均匀。样地S30共有13种植物分属2门3纲6目7科11属, 其中菊科含种最多(7种), 其他皆为单属单种的科。相反, 样地S8(垣曲毛家弯)分类差异变异指数Λ+(304.66)低于其余样地, 基本接近区域内的理论平均值, 这说明S8物种等级分布相对均匀。样地S8共有28种植物, 隶属2门3纲16目20科26属, 其中毛茛目、蔷薇目、管状花目和中央种子目各有两个科, 其他各目仅包含一个科。豆科、菊科、堇菜科含物种数最多, 各有3种植物。
由Δ+和Λ+与种的分布可以看出, Δ+和Λ+随着物种的增加基本呈现出下降的趋势。样地S102(宁武接官亭)种数最多, 共43种, Δ+和Λ+值相对较低, 分别为54.4和434.47。样地S32种数最少, 虽然仅有7种植物但它们分属2门3纲5目5科6属, 平均分类差异指数(63.04)较高, 分类差异变异指数(874.28)较高。
3.3 分类学多样性指数与环境因子和群落结构的相关分析由表 1可知, 宏观地形因子与Δ+相关不显著(P>0.05), 与Λ+相关亦不显著(P>0.05), 其中纬度和海拔与Δ+和Λ+相关性大于经度, 主要原因是山西纬度跨度大于经度和海拔梯度且变化明显。微地形因子中, 坡向与Δ+极显著正相关(P < 0.01), 坡位与Δ+显著正相关(P < 0.05), 与Λ+极显著正相关(P < 0.01)。
| 指数 Index |
纬度 Latitude |
经度 Latitude. |
海拔 Altitude |
坡度 Slope |
坡向 Aspect |
坡位 SP |
pH pH |
全氮 TN |
全磷 TP |
有机碳 SOC |
胸径 DBH |
密度 Density |
物种数 S |
| Δ+ | 0.070 | 0.005 | 0.117 | 0.015 | -0.240** | 0.175* | 0.012 | 0.317** | -0.147 | 0.315** | 0.214* | -0.028 | -0.540** |
| Λ+ | -0.103 | -0.031 | -0.050 | -0.095 | 0.025 | 0.225** | 0.048 | 0.132 | 0.010 | 0.131 | 0.048 | 0.210* | -0.579** |
| SP:坡位Slope position;TN:全氮Total nitrogen;TP:全磷Total phosphorus;SOC:有机碳Soil organic carbon;DBH:胸径Diameter at breast height;S:物种数Number of species | |||||||||||||
土壤属性因子与Λ+相关不显著(P > 0.05);全氮和有机碳与Δ+呈极显著正相关, pH和全磷与Δ+相关不显著(P > 0.05)。
群落结构因子中, 物种数与Δ+呈极显著负相关(P < 0.01), 与Λ+呈极显著负相关(P < 0.01);乔木密度与Δ+相关不显著(P > 0.05), 与Λ+显著正相关(P < 0.05);胸径与Δ+显著正相关(P < 0.05), 与Λ+相关不显著(P>0.05)。由此可以看出, 影响油松林Δ+和Λ+分布的最直接的因子主要是微地形因子和群落结构因子。
4 讨论山西油松林的植物主要集中于菊科、豆科、禾本科、莎草科。多数样地的Λ+值高于理论平均值, 表明山西油松群落物种分布均一性较弱。Pienkowski等研究表明, Δ+值落入95%的置信漏斗最小值曲线以下时, 表示该样地的生境发生退化或受到严重污染[13, 31]。山西油松林所有样地Δ+值没有位于置信漏斗最小值曲线下方的, 表明山西油松林分布区域的生态环境整体情况良好, 受到的各种干扰较少。这主要得益于1998年以来的退耕还林政策的实施和山西省生态文明建设各种管理措施的全面落实, 包括停止对油松林的采伐、加强对盗伐油松林的处罚和监管等。五老峰和天龙山为旅游胜地, 油松林处于中幼林阶段, 林隙间较为充足的光照有利于林下灌木和草本的生长, 使得林下物种多样性更丰富, 群落分类学多样性增高;同时, 旅游对油松林有轻度干扰, 加之一些植物繁殖体也可能随游人进入松林, 其中某些种可能在油松林定居并成功完成生活史, 使得样地S6和S78具有较高的分类学多样性。样地S85、S86、S87、S90、S112, 属于国有林场, 油松林发育良好, 群落结构完整, 乔木层可分为两个亚层, 其中第一亚层为建群种——油松, 第二亚层物种较为丰富, 常见的有辽东栎、山杨、白桦等, 灌木层和草本层发育也较为完整, 从而使得这些样地具有较高的分类学多样性。这一结果与其他研究者的结论一致[3], 即结构复杂的群落具有更高的分类学多样性指数。样地S12位于安泽草峪岭乡道附近, 垦荒、出行等人类日常活动严重干扰了林下和林缘植被的生长和种子的传播;油松林处于幼林阶段, 且位于阳坡, 水分条件较差, 不利于林下植物的生长, 分布的草本大多为菊科和禾本科等抗逆性强的种。因此, 样地S12的Δ+值最小。
相关性分析(表 1)表明, 尽管山西油松林分布的纬度和海拔变异明显, 但纬度和海拔并不是导致Δ+和Λ+分布的差异的决定性因子, 亦即平均分类差异指数Δ+并没有表现出明显的纬度和海拔变化规律, 与李永振等研究结果类似[11, 32], 这说明Δ+指数在区域尺度上反映群落的分类学多样性具有较强的稳健性;同时, 也表明山西是油松林分布相对适宜的区域, 林下物种组成主要以菊科、禾本科等为主, 它们具有较广的生态幅和适应性。微地形因子中, 坡向与Δ+极显著正相关(P < 0.01), 即阴坡的油松林具有较高的分类学多样性。坡向的差异会影响光照和水分的空间异质性[20]。在山西油松林分布区光照充足, 限制植物生长和分布的主要是水分因子。在阳坡由于光照时间长、强度大, 分布的物种往往以耐旱性较强的植物为主;由于土壤持水量较低, 种间对水分的竞争强度相对较大, 物种组成往往集中于某一科或某一属。如样地S30位于襄垣古韩乡土桥村海拔1122 m的阳坡, 充分的光照使得喜阳的油松长势良好, 乔木层郁闭度较大(0.85)不利于林下灌木和草本的生存;S30共有13种植物分属2门3纲6目7科11属, 其中含种最多的科为菊科(7种), 其他科皆为单属单种, 如展枝沙参(Adenophora divaricata)、披针薹草、角盘兰(Herminium monorchis)、油松、茜草(Rubia cordifolia)、鸡腿堇菜(Viola acuminata)。位于交城庞泉沟阴坡的样地S90, 由于水分条件好于阳坡, 乔木层郁闭度较小(0.55), 林下灌木层和草本层组成丰富多样, 共有22种植物分属2门3纲13目16科21属, 包括侧柏(Platycladus orientalis)、灰栒子(Cotoneaster acutifolius)、美蔷薇、野青茅(Deyeuxia arundinacea)、筋骨草(Ajuga ciliata)、北柴胡(Bupleurum chinense)、异鳞薹草(Carex heterolepis)等不同生活型种, 使得样地分类多样性指数较大。
坡位主要反映的是立地条件、水分状况以及人为干扰等情况;坡位与Δ+显著正相关(P < 0.05), 与Λ+极显著正相关(P < 0.01)。赵小娜等[19]研究结果表明坡位与Δ+正相关(P > 0.05), 与Λ+负相关(P > 0.05), 是由于研究区域位于庞泉沟自然保护区, 海拔1600—2400 m, 温度变化明显, 且受人为活动干扰较少, Δ+和Λ+主要受海拔梯度的影响。本文研究的油松林海拔分布相对较低(744—1800 m), 热量条件相对较好, 人为活动干扰较多, 林下的物种组成相对简单。群落物种数与Δ+和Λ+呈极显著负相关(P < 0.01);密度与Λ+显著正相关(P < 0.05);胸径与Δ+显著正相关(P < 0.05)。如位于宁武接官亭林场的样地S102物种数最多, 共43种, 属2门3纲19目24科38属, 乔木层密度较小郁闭度低(0.60), 林下种类丰富, 灌木层优势种为土庄绣线菊, 草本优层优势种为披针薹草和东方草莓(Fragaria orientalis), Δ+和Λ+值相对较低。位于武乡县石门乡石门村的样地S32物种数最少, 仅出现油松、披针薹草、茜草、小红菊(Dendranthema chanetii)、白莲蒿(Artemisia sacrorum)、牛尾蒿(Artemisia dubia)等, 植物但它们分属2门3纲5目5科6属, 因此表现出较高的Δ+和Λ+值。
微地形因子和群落结构因子导致物种的微生境差异。已有研究表明,与微生境相比地形因子对分类学多样性影响较小[11], 本文的结果与此类似。群落结构因子对分类学多样性影响最大可能是因为群落演替早期具有若干生态位相似或对环境有相似响应的近缘种; 随着演替的进行, 物种对水分、光照等环境因子的需求增多, 种间对相似资源的激烈竞争使近缘种逐渐减少, 而使亲缘关系较远的物种得以实现稳定共存, 群落趋向成熟, 并最终达到顶级群落阶段。
致谢: 感谢山西大学黄土高原研究所苏智娇、赵蓉、张淼淼、李旭华、邓永利、毛空、王洁等同学参与野外调查。| [1] | Neeson T M, Van Rijn I, Mandelik Y. How taxonomic diversity, community structure, and sample size determine the reliability of higher taxon surrogates. Ecological Applications , 2013, 23 (5) : 1216–1225. DOI:10.1890/12-1167.1 |
| [2] | Jucker T, Carboni M, Acosta A T R. Going beyond taxonomic diversity:Deconstructing biodiversity patterns reveals the true cost of iceplant invasion. Diversity and Distributions , 2013, 19 (12) : 1566–1577. DOI:10.1111/ddi.2013.19.issue-12 |
| [3] | 秦晓娟, 董刚, 邓永利, 毛空, 李旭华, 张峰. 山西平陆黄河湿地植物分类学多样性研究. 生态学报 , 2015, 35 (2) : 409–415. |
| [4] | 蒋志刚, 纪力强. 鸟兽物种多样性测度的G-F指数方法. 生物多样性 , 1999, 7 (3) : 220–225. |
| [5] | Moreno C E, Castillo-Campos G, Verdú J R. Taxonomic diversity as complementary information to assess plant species diversity in secondary vegetation and primary tropical deciduous forest. Journal of Vegetation Science , 2009, 20 (5) : 935–943. DOI:10.1111/jvs.2009.20.issue-5 |
| [6] | Clarke K R, Warwick R M. A taxonomic distinctness index and its statistical properties. Journal of Applied Ecology , 1998, 35 (4) : 523–531. DOI:10.1046/j.1365-2664.1998.3540523.x |
| [7] | Graham N A J, McClanahan T R, Letourneur Y, Galzin R. Anthropogenic stressors, inter-specific competition and ENSO effects on a Mauritian coral reef. Environmental Biology of Fishes , 2007, 78 (1) : 57–69. |
| [8] | Rogers S I, Clarke K R, Reynolds J D. The taxonomic distinctness of coastal bottom-dwelling fish communities of the North-east Atlantic. Journal of Animal Ecology , 1999, 68 (4) : 769–782. DOI:10.1046/j.1365-2656.1999.00327.x |
| [9] | Ceschia C, Falace A, Warwick R. Biodiversity evaluation of the macroalgal flora of the Gulf of Trieste (Northern Adriatic Sea) using taxonomic distinctness indices. Hydrobiologia , 2007, 580 (1) : 43–56. DOI:10.1007/s10750-006-0466-8 |
| [10] | 徐宾铎, 金显仕, 梁振林. 黄海鱼类群落分类学多样性的研究. 中国海洋大学学报:自然科学版 , 2005, 35 (4) : 629–634. |
| [11] | 李永振, 史赟荣, 艾红, 董丽娜, 李娜娜, 李夏, 高天翔. 南海珊瑚礁海域鱼类分类多样性大尺度分布格局. 中国水产科学 , 2011, 18 (3) : 619–628. |
| [12] | Richard M. The use of taxonomic distinctness to assess environmental disturbance of insect communities from running water. Freshwater Biology , 2007, 52 (8) : 1634–1645. DOI:10.1111/fwb.2007.52.issue-8 |
| [13] | Mouillot D, Laune J, Tomasini J A, Aliaume C, Brehmer P, Dutrieux E, Do Chi T. Assessment of coastal lagoon quality with taxonomic diversity indices of fish, zoobenthos and macrophyte communities. Hydrobiologia , 2005, 550 (1) : 121–130. DOI:10.1007/s10750-005-4368-y |
| [14] | 丁兰平. 两个新的群落多样性指数. 海洋科学 , 1999, 21 (1) : 25–28. |
| [15] | Leonard D R P, Clarke K R, Somerfield P J, Warwick R M. The application of an indicator based on taxonomic distinctness for UK marine biodiversity assessments. Journal of Environmental Management , 2006, 78 (1) : 52–62. DOI:10.1016/j.jenvman.2005.04.008 |
| [16] | Ellingsen K E, Clarke K R, Somerfield P J, Warwick R M. Taxonomic distinctness as a measure of diversity applied over a large scale:the benthos of the Norwegian continental shelf. Journal of Animal Ecology , 2005, 74 (6) : 1069–1079. DOI:10.1111/jae.2005.74.issue-6 |
| [17] | 李景文, 李俊清, 任艳林. 海南主要热带森林生态系统类型自然保护区物种分类多样性研究. 北京林业大学学报 , 2002, 24 (5/6) : 108–114. |
| [18] | Da Silva I A, Batalha M A. Taxonomic distinctness and diversity of a hyperseasonal savanna in central Brazil. Diversity and Distributions , 2006, 12 (6) : 725–730. DOI:10.1111/ddi.2006.12.issue-6 |
| [19] | 赵小娜, 秦晓娟, 董刚, 张峰. 庞泉沟自然保护区植物群落分类学多样性. 应用生态学报 , 2014, 25 (12) : 3437–3442. |
| [20] | 陈姣, 廉凯敏, 张峰, 王建军, 史荣耀. 山西历山保护区野生种子植物区系研究. 山西大学学报:自然科学版 , 2012, 35 (1) : 151–157. |
| [21] | 刘蕾, 张峰. 历山白云洞景区植物群落物种多样性研究. 山西大学学报:自然科学版 , 2010, 33 (3) : 468–472. |
| [22] | 狄晓艳.油松遗传多样性与光合生理生态特性研究[D].太原:山西大学, 2014. http://cdmd.cnki.com.cn/article/cdmd-10108-1015370279.htm |
| [23] | 张峰. 山西油松林分布区气候因素的排序研究. 山西大学学报:自然科学版 , 1990, 13 (3) : 322–327. |
| [24] | 张新波, 李悦, 袁虎威, 李伟, 富裕华, 刘致远, 朱松林. 山西油松天然林分21年子代生长性状遗传变异研究. 北京林业大学学报 , 2014, 36 (3) : 104–109. |
| [25] | 方精云, 王襄平, 沈泽昊, 唐志尧, 贺金生, 于丹, 江源, 王志恒, 郑成洋, 朱江玲, 郭兆迪. 植物群落清查的主要内容、方法和技术规范. 生物多样性 , 2009, 17 (6) : 533–548. |
| [26] | Schaminée J H J, Hennekens S M, Chytrý M, Rodwell J S. Vegetation-plot data and databases in Europe:an overview. Preslia , 2009, 81 (3) : 173–185. |
| [27] | Clarke K R, Warwick R M. The taxonomic distinctness measure of biodiversity:weighting of step lengths between hierarchical levels. Marine Ecology Progress Series , 1999, 184 : 21–29. DOI:10.3354/meps184021 |
| [28] | 曾燏. 嘉陵江干流鱼类物种分类多样性研究. 西华师范大学学报:自然科学版 , 2012, 33 (3) : 246–260. |
| [29] | Clarke K R, Warwick R M. A further biodiversity index applicable to species lists:variation in taxonomic distinctness. Marine Ecology Progress Series , 2001, 216 (2) : 265–278. |
| [30] | Ricotta C, Avena G C. An information-theoretical measure of taxonomic diversity. Acta Biotheoretica , 2003, 51 (1) : 35–41. DOI:10.1023/A:1023000322071 |
| [31] | Pienkowski M W, Watkinson A R, Kerby G, Warwick R M, Clarke K R. Taxonomic distinctness and environmental assessment. Journal of Applied Ecology , 1998, 35 (4) : 532–543. DOI:10.1046/j.1365-2664.1998.3540532.x |
| [32] | 李圣法.东海大陆架鱼类群落生态学研究-空间格局及其多样性[D].上海:华东师范大学, 2005. http://cdmd.cnki.com.cn/article/cdmd-10269-2005086168.htm |