 2016, Vol. 36
2016, Vol. 36文章信息
- 罗久富, 郑景明, 周金星, 张鑫, 崔明
- LUO Jiufu, ZHENG Jingming, ZHOU Jinxing, ZHANG Xin, CUI Ming.
- 青藏高原高寒草甸区铁路工程迹地植被恢复过程的种间关联性
- Analysis of the interspecific associations present in an alpine meadow community undergoing revegetation on the railway-construction affected land of the Qinghai-Tibet Plateau
- 生态学报[J]. 2016, 36(20): 6528-6537
- Acta Ecologica Sinica[J]. 2016, 36(20): 6528-6537
- http://dx.doi.org/10.5846/stxb201502030276
-
文章历史
- 收稿日期: 2015-02-03
- 网络出版日期: 2016-01-22





2. 北京林业大学水土保持学院, 北京 100083;
3. 西北农林科技大学林学院, 杨凌 712100;
4. 中国林业科学研究院荒漠化研究所, 北京 100091
2. School of Soil and Water Conservation, Beijing Forestry University, Beijing 100083, China;
3. College of Forestry, Northwest A & F University, Yangling 712100, China;
4. Institute of Dersertification, Chinese Academy of Forestry, Beijing 100091, China
植物群落的本质特征是整体性和差异性的统一, 也就是群落内的物种之间存在相互关联, 这种关联决定着群落的结构和动态[1-2]。种间关联包括联结性和相关性两个方面, 种间联结性是指不同物种在空间分布上的相互关联性, 通常是由于群落生境的差异影响了物种的分布而引起的[3], 是一种定性关系;种间相关性也是指不同种类在空间分布上的相互关联性, 但是不局限于物种存在与否的二元数据, 同时还与物种的数量数据(如盖度、高度等)有关, 是一种定量的关系。由于χ2检验是基于物种有/无的二元数据进行检验的, 只能定性的分析出物种间是否存在关联, 而不能检验出关联程度的大小[4-5]。因此, 在量化的盖度数据的基础上, 结合方差检验和Spearman秩相关系数检验来分析不同时期群落内主要物种之间的相关性, 能更精确、全面地分析一定时期内群落物种间的关联性变化动态[6-7]。一般认为, 在一个群落中一个物种依赖另一个物种或两个物种具有相似的生活习性会呈现出正关联, 而相互竞争或者生态位分离的物种之间则表现为负关联[4, 8-9]。因此, 研究一个群落内物种在不同时期的种间关联变化, 能加深对群落性质的理解, 正确认识生物与非生物因素在群落物种组配过程中的地位和作用[4, 10]。
国内外学者对森林、湿地、草原等各类植物群落的种间关联开展了大量研究[1-2, 8-9, 11-12]。针对高山草地群落, 国外学者研究了道路修建、干旱胁迫等对其组成、结构等的影响, 以及高山地区垫状植被与其他物种的相关关系等[9, 13-16], 如O′Connor和Aarssen认为, 植物群落物种间正负联结比例和联结强度会随着演替的推进而增强[9], Alex等对高山垫状植物的研究发现, 物种之间存在复杂的相互关联性, 并且受到生物环境因子的影响[16];国内多是基于一次性调查数据的物种关联性分析[5, 11, 17-22], 如邢福等对糙隐子草草原的物种关联性研究发现, 随着放牧压力增加, 物种间总体上趋向于无关联[11];房飞等发现, 在新疆亚高山草地中, 原始植被物种间多成正联结, 而退化植被的物种间则表现出无联结或负联结[22]。对大型工程或干扰后植被群落变化的研究相对较少。对汶川地震滑坡迹地植被恢复过程的研究发现, 震后5a的群落中正关联物种对数低于负关联, 达到显著关联的比例也较低[23];齐麟等研究发现, 加强采伐的干扰力度会使某些树种之间由正关联转变为负关联[24]。本研究采用长期样地监测数据, 对青藏高原高寒草甸区铁路工程迹地植物群落物种关联性动态变化进行研究。
青藏高原高寒草甸区自然地理和气候条件独特, 保存有相对完整的特殊生态系统及特有的珍稀动植物资源, 具有特殊的生态价值以及科研价值[25]。青藏铁路贯穿大部分高寒植被区, 受铁路工程迹地影响最大的植被类型之一就是高寒草甸植被[26], 而这种影响究竟如何, 本研究以该区域内青藏铁路工程迹地植物群落为研究对象, 对以高山嵩草(Kobresia pygmaea)等为主要优势种的高寒草甸区设置长期监测样地[26-28], 进行种间关联动态研究, 旨在揭示高寒草甸区工程迹地植被自然恢复过程规律, 探讨高寒脆弱环境条件下的自然植被受到大型工程干扰后能否通过自然恢复达到稳定状态, 为青藏高原高寒草甸区工程迹地植被恢复和多样性保护提供参考。
1 研究区域研究区域主要位于唐古拉山与当雄县之间(91°06′—91°44′E, 33°04′—30°28′N), 最高海拔5072m, 最低海拔4549m。年均温-7—0℃, 年降水300—400mm。根据《中国植被》相关分类和实地调查可知该区域主要是高寒草甸景观, 土壤以高寒草甸土为主[29], 土壤风化程度较低, 粗骨性强, 土层较薄, 土层下面多砾石, 透水性强, 保水型差。地下有永冻层, 阻碍植物根系生长和发育, 水分难下渗。主要植物物种有高山嵩草, 青藏薹草(Carex moorcroftti), 紫花针茅(Stipa purpurea)等。青藏铁路从2001年开始动工, 2005年基本完成路轨建设, 2006年铁路正式通车运行。
2 数据来源与研究方法 2.1 植被调查方法样地布设范围为唐古拉山口至乌玛塘, 由于青藏铁路护栏外工程迹地主要在距离护栏0—20m之间, 所以自唐古拉山口起, 每隔50km在护栏外距离护栏5m处设置一块样地, 共8块样地, 每块样地10m×40m, 长边与青藏铁路平行, 在样地中线上每隔5m布设一个1m×1m的样方, 每块样地共9个样方, 共72个样方。分别于铁路建设期(2005年8月)、植被恢复4年后(2009年8月)和植被恢复8年后(2013年8月)对每块固定样地进行群落学调查, 内容主要包括植物种类、盖度、高度、株数等, 记录海拔、经纬度等。本研究选取重要值大于0.01的物种作为群落主要物种进行物种关联性分析[4, 30]。
2.2 数据分析方法 2.2.1 物种重要值采用重要值综合指标来处理数据, 物种重要值(IV)用下式计算[4, 31]:

|
总体关联性是一个群落中所有物种的关系, 可以通过计算种间联结指数(VR)来测定所研究对象的总体关联性[32]。其计算公式为:
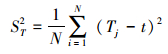
|
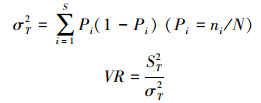
|
式中, S为调查地区总的物种数;N为总样方数;Tj为样方j内出现的物种总数;t为全部样方中种的平均数;σT为所有物种出现频度的方差;ST为所有样方物种数的方差;ni为物种i出现的样方数;Pi为物种出现的频度。独立性假设条件下群落内植物种间总体联结指数VR期望值为1。若VR=1, 接受零假设, 即种间总体无关联;若VR>1, 则否定零假设, 即物种间总体呈正关联;若VR < 1, 则物种总体呈负关联。为检验VR偏离1是否显著, 引入统计量W, W=VR×N。W服从χ2分布(df=N-1)。若W < χ0.95N2或W>χ0.05N2, 则物种间总体关联显著(P2 < 0.05);反之, 若χ0.95N2 < W < χ0.05N2, 则物种间总体关联不显著(P2 > 0.05)。
2.2.3 种对间联结性分析种间联结一般采用χ2检验进行定性研究, 根据基于2×2列联表的χ2统计量进行联结性分析[33], 其公式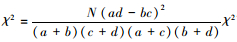
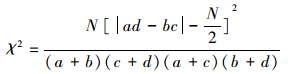
|
式中, N为样方总数;a为两物种均出现的样方数;b、c分别为只有1种物种出现的样方数;d为两物种均未出现的样方数。当ad>bc时, 两物种为正联结, adY bc时, 两物种为负联结。若χ2 < 3.841(P2>0.05), 则表示两个种独立分布, 为中性联结;若3.841 < χ2 < 6.635(0.01 < P < 0.05), 表示种对间联结显著;若χ2>6.635(P2 < 0.01), 则表示种对间联结极显著。
2.2.4 种对间相关性分析用定量数据(盖度)作为Spearman秩相关分析的数量指标[31]。
Spearman秩相关系数计算公式:
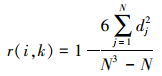
|
式中, N为总样方数, dj=(xij-xkj), xij和xkj分别为种i和种k在样方j中的秩。
3 结果与分析 3.1 受铁路工程干扰后植物群落的物种变化3次调查结果显示, 群落平均盖度2005年为(35.21±4.41)%、2009年为(33.42±3.01)%、2013年为(43.41±3.26)%;在72个样方内, 铁路建设期仅49个样方调查到植物, 共71种, 隶属18科42属;恢复4年后有69个样方调查到植物, 共78种, 隶属23科53属;恢复8年后有71个样方调查到植物, 共85种, 隶属23科57属。调查结果如表 1和图 1所示, 表明恢复过程中群落盖度和物种发生变化, 适生于湖边、沼泽草甸等湿生环境的毛茛科(高原毛茛(Ranunculus tanguticus))、大戟科(青藏大戟(Euphorbia altotibetica))、虎耳草科(西藏虎耳草(Saxifraga tibetica))等植物种逐渐减少甚至消失, 而常见于山坡、砂砾地等干旱环境中的景天科(大炮山景天(Sedum erici-magnusii)、藏布红景天(Rhodiola smithii)等)、百合科(镰叶韭(Allium carolinianum))、藜科(平卧轴藜(Axyris prostrata)、垫状驼绒藜(Ceratoides compacta)等)植物种数逐渐增加。
| 序号No. | 物种Species | 重要值IV | ||
| 2005 | 2009 | 2013 | ||
| 1 | 阿尔泰葶苈Draba altaica | 0.0137 | ||
| 2 | 矮生嵩草K. humilis | 0.0220 | 0.0192 | 0.0110 |
| 3 | 矮羊茅Festuca coelestis | 0.0220 | 0.0153 | 0.0130 |
| 4 | 白花枝子花Dracocephalum heterophyllum | 0.0114 | ||
| 5 | 波伐早熟禾Poa poophagorum | 0.0521 | 0.0754 | 0.0826 |
| 6 | 臭蒿Artemisia hedinii | 0.0185 | ||
| 7 | 丛生黄耆Astragalus confertus | 0.0134 | ||
| 8 | 粗壮嵩草K. robusta | 0.0124 | 0.0159 | |
| 9 | 垫状点地梅Androsace tapete | 0.0105 | ||
| 10 | 垫状金露梅Potentilla fruticosa var. pumila | 0.0201 | 0.0453 | 0.0255 |
| 11 | 叠裂银莲花Anemone imbricata | 0.0229 | 0.0291 | 0.0119 |
| 12 | 短穗兔耳草Lagotis brachystachya | 0.0115 | 0.0371 | 0.0243 |
| 13 | 多茎委陵菜P. multicaulis | 0.0143 | ||
| 14 | 多裂委陵菜P. multifida | 0.0118 | 0.0170 | |
| 15 | 二裂委陵菜P. bifurca | 0.0149 | 0.1015 | 0.1254 |
| 16 | 高山嵩草K. pygmaea | 0.3580 | 0.0906 | 0.1110 |
| 17 | 黑褐穗薹草C. atrofusca subsp. minor | 0.0179 | ||
| 18 | 卷鞘鸢尾Iris potaninii | 0.0114 | ||
| 19 | 鳞叶龙胆Gentiana squarrosa | 0.0105 | ||
| 20 | 密丛棘豆Oxytropis densa | 0.0127 | ||
| 21 | 平卧轴藜Axyris prostrata | 0.0200 | ||
| 22 | 草Koeleria cristata | 0.0127 | 0.0265 | 0.0400 |
| 23 | 青藏薹草C. moorcroftti | 0.0178 | 0.0343 | |
| 24 | 肉果草Lancea tibetica | 0.0184 | 0.0246 | 0.0312 |
| 25 | 弱小火绒草Leontopodium pusillum | 0.0357 | 0.0277 | 0.0433 |
| 26 | 沙生风毛菊Saussurea arenaria | 0.0123 | 0.0239 | |
| 27 | 梭罗草Roegneria thoroldiana | 0.0103 | 0.0194 | |
| 28 | 团垫黄耆A. arnoldii | 0.0114 | ||
| 29 | 萎软紫菀Aster flaccidus | 0.0192 | 0.0165 | |
| 30 | 无茎黄鹌菜Youngia simulatrix | 0.0124 | ||
| 31 | 西伯利亚蓼Polygonum sibiricum | 0.0105 | ||
| 32 | 纤杆蒿A. demissa | 0.0330 | ||
| 33 | 羊茅F. ovina | 0.0151 | 0.0195 | |
| 34 | 窄叶薹草C. montis-everestii | 0.0705 | 0.0446 | 0.0510 |
| 35 | 珠光香青Anaphalis margaritacea | 0.0150 | 0.0129 | |
| 36 | 紫花针茅S. purpurea | 0.0426 | 0.0457 | 0.0565 |
| 37 | 紫羊茅F. rubra | 0.0515 | ||
| —表示重要值小于0.01 | ||||

|
| 图 1 2005年、2009年和2013年固定样地中各科所含物种数的比例 Fig. 1 The proportions of species in families in 2005, 2009 and 2013 景天科Crassulaceae;藜科Chenopodiaceae;百合科Liliaceae;麻黄科Ephedraceae Dumortier;车前科Plantaginaceae;茜草科Rubiaceae;紫草科Boraginaceae;报春花科Primulaceae;唇形科Labiatae;鸢尾科Iridaceae;蔷薇科Rosaceae;伞形科Umbelliferae;十字花科Cruciferae;罂粟科Papaveraceae;龙胆科Gentianaceae;禾本科Gramineae;豆科Leguminosae;菊科Compositae;莎草科Cyperaceae;茄科Solanaceae;蓼科Polygonaceae;石竹科Caryophyllaceae;玄参科Scrophulariaceae;毛茛科Ranunculaceae;大戟科Euphorbiaceae;虎耳草科Saxifragaceae |
重要值大于0.01的主要物种名称及重要值见表 1。
随着时间的推移, 群落内主要物种的种类和数量也发生了变化, 恢复初期群落内IV大于0.01物种数为23种, 高山嵩草在群落内占据绝对优势, 其重要值(IV=0.3580)远远高于其它物种, 恢复4年后增加到27种, 恢复8年后, 降为21种。
3.2 物种间的总体关联性分析3次调查的物种间的总体关联性统计结果见表 2。可见历次调查的群落内物种的总体关联性都为显著正关联(P < 0.05), 联结指数(VR)大小关系为VR2005 > VR2009 > VR2013, 这表明受到铁路工程干扰后, 植物群落内物种总体关联虽保持显著正关联, 但是随着时间推移正关联强度呈现出减弱的趋势。表明铁路建设对该区域内高寒草甸植物物种间的总体关联性产生了一定的影响, 群落内各物种之间的依赖性可能随着时间的增加逐渐减弱。
| 年份 Year | 联结指数VR Association Index VR | 统计量W Statistic W | 总体关联性 Overall association |
| 2005 | 5.33 | 383.88 | 显著正关联 |
| 2009 | 3.10 | 223.11 | 显著正关联 |
| 2013 | 1.63 | 117.02 | 显著正关联 |
| χ0.095(72)2=53.47, χ0.05(72)2=92.81 | |||
2005年、2009年和2013年3次调查的主要物种种对间的联结性χ2检验(P=0.05)结果表明, 正联结种对数所占的百分比总体呈降低趋势, 负联结种对数比例增加(图 2), 3个阶段正负联结种对百分数之比分别为1.88、0.94和1.04。这说明, 最初恢复的4a时间里群落内种间关系经历了剧烈地变化, 后4a时间内群落变动较弱, 正联结比重微弱回升。

|
| 图 2 历次调查主要物种种间χ2检验正关联、负关联和无关联种对数百分比 Fig. 2 Percentage of dominant species pairs with positive, negative and non-association of three periods in alpine meadow |
在Spearman秩相关系数检验中(表 3, 图 3), 3次调查中大部分物种种对间都未达到显著相关, 种间联结较为松散, 各物种独立性较强, 群落稳定性较弱, 容易受到外界干扰而发生改变。2005年253对物种对中, 正相关种对数162对, 负相关91对, 2009年351对物种对中, 正相关种对数166对, 负相关185对, 2013年210对物种对中, 正相关种对102对, 负相关108对。负相关比重增加, 且达到显著负相关的种对数百分比增加, 尤其是最初4a恢复时期内物种负相关比重迅速增加, 后4a增加幅度则相对减弱, 8年间达到显著正相关种对数百分比从37.16%降低到18.57%。同时部分相同物种之间的关联程度也发生了变化, 如11-15(二裂委陵菜-叠裂银莲花)在3次调查结果中呈现不显著负相关-显著负相关-极显著负相关的变化, 11-16(叠裂银莲花-高山嵩草)种对在2005年和2009年均表现为不显著负相关, 2013年则为显著负相关, 10-15(垫状金露梅-二裂委陵菜)种对在2005年表现为不显著负相关, 2009年和2013年均表现为极显著负相关。
| 年份 Year | 正关联种对数百分比 Percentage of positive association/% | 负关联种对数百分比 Percentage of negative association/% | 无关联 Null | |||||
| 极显著 Highly significant | 显著 Significant | 不显著 Not significant | 极显著 Highly significant | 显著 Significant | 不显著 Not significant | |||
| 2005 | 28.46 | 8.70 | 26.88 | 0 | 0 | 35.97 | 0 | |
| 2009 | 13.96 | 5.98 | 27.35 | 0.85 | 1.42 | 50.43 | 0 | |
| 2013 | 11.90 | 6.67 | 30.00 | 1.90 | 6.19 | 43.33 | 0 | |
| 极显著P < 0.01, 显著P < 0.05, 不显著P≥0.05 | ||||||||

|
| 图 3 3次调查优势种群间的Spearman秩相关系数半矩阵图 Fig. 3 Semi-matrix of Spearman′s rank correlation coefficients of dominant population |
结合χ2检验和Spearman秩相关系数检验分析, 青藏高原高寒草甸区铁路工程迹地植被主要种在最初4a的恢复期间负相关种对数比例明显增加, 达到显著或极显著负相关的种对数所占的百分比也呈现上升趋势, 达到显著或极显著正相关的种对数所占的百分比降低, 但恢复第4年到第8年间正负相关比例变化不明显。
4 讨论植物群落中各物种的关系决定着群落结构特征和动态, 研究群落内物种的关联性是了解群落稳定性和分析群落动态的重要途径[35]。
4.1 高寒草甸区铁路工程迹地植被恢复过程中群落结构变化规律等级演替理论指出, 不同物种对裸地的利用能力存在差异, 对不同裸地的适应能力也不同, 裸地的可利用性与物种的繁殖体生产力、传播能力以及萌发和生长能力紧密相关[36]。本研究中, 工程迹地植被在8a的自然恢复过程中, 恢复时间较短, 物种组成不稳定, 群落盖度及群落内物种数总体上呈增加趋势, 群落内耐旱植物种类增加。能在极端干旱或寒冷环境下存活并繁殖的基本上是抗旱性或耐寒性最好的物种[37]。铁路工程迹地土壤物理结构遭到破坏, 影响植物根系生长以及水分下渗, 地表径流增加, 导致该区域内土壤含水量降低, 形成干旱环境[38];另外通车运行之后, 火车通过时小环境的气流扰动加速地表水分蒸发也可能是造成该地域趋向旱生环境的原因之一[15]。恢复初期, 群落优势种重要值最高的为高山嵩草(IV2005=0.3593), 二裂委陵菜地位中等, 4年后及8年后群落内重要值最高的均为二裂委陵菜;不适宜干扰后的干旱生境的植物如西藏虎耳草等逐渐消失, 适应恶劣环境能力强的藜科物种出现并占据较高的重要值, 如高寒荒漠及高寒荒漠草
原建群种平卧轴藜、垫状驼绒藜(IV2013=0.0083)出现, 且平卧轴藜与纤杆蒿、臭蒿等耐旱物种呈正相关关系, 说明受到干扰后, 由于高山嵩草和二裂委陵菜对环境适应能力不同, 高山嵩草的无性系分蘖数下降[39], 生态幅广的旱生植物二裂委陵菜等迅速扩张。李丹雄等对汶川地震滑坡迹地植被恢复研究也发现, 在自然恢复过程中, 滑坡迹地群落物种多以菊科和禾本科等耐旱耐瘠薄物种为主[23]。总体来说, 铁路工程迹地群落在8a恢复期间物种数增加, 耐寒耐旱植物种增多, 更多不同生态位的物种共存于群落中, 表明群落的非生物环境条件发生了某种程度的改变。
4.2 高寒草甸区铁路工程迹地植被恢复过程中种间关联变化规律在植物群落发展过程中, 有学者提出, 早期群落内物种大量侵入定居, 物种间激烈竞争, 群落变化剧烈, 当发展到一定的阶段, 群落变化速度减缓[37, 40]。本研究中群落内的植被恢复8年间, 群落内物种间的正负联结比例表现为降低趋势, 达到显著的种对数比重降低, 尤其是最初4a时间里, 正关联种对数比重明显减小, 变化剧烈, 后4a变化不明显, 该结果与O′Connor和Aarssen等预测的物种关联性变化与群落发展趋势的关系不同, 他们认为随演替的推进物种间的正负联结比例增加, 联结强度增强[9], 而在本研究中前4年群落内正负联结比例明显降低, 说明群落受到铁路工程干扰后, 在恢复阶段初期群落内发生剧烈变化, 物种迅速向裸地扩张, 处于大量侵入定居阶段, 并且各物种间发生生态位竞争, 之后各物种开始适应干扰的生境, 铁路工程迹地植被在经历剧烈变化之后开始恢复, 但仍未达到稳定阶段。导致差异的主要原因可能是各研究的时间尺度、干扰类型、非生物因子胁迫程度以及植被发展变化的初始状态等不同所致, O′Connor等研究针对的是不同恢复年限的撂荒地群落, 而本研究对象是环境极端严酷的青藏高原, 青藏铁路的修建和之后的运行对冻土、土壤和植被的影响方式等都不同于其他研究。
研究发现, 工程迹地群落中二裂委陵菜与叠裂银莲花等多对种对间的关联程度随时间发生变化, 负相关性增强, 说明这些种对间竞争强度增加。由竞争理论可知, 植物对资源的相似或者不相似的需求使各物种之间存在相关关系, 而不同物种对同一资源获取的难易程度影响着这些物种间的竞争强度, 因而即使是相同种对在不同的恢复阶段也可能表现出不同的相关关系[41-42]。齐麟等也指出种对间的关系会因外界条件的改变而发生改变[24]。这种物种间关系的变化可能与环境资源的可利用性有关, 更多物种利用相同资源, 导致物种间的竞争程度以及产生竞争的物种数增加[43]。
5 结论及建议青藏高原高寒草甸区铁路工程迹地植被8a恢复时间内, 群落盖度增加, 耐旱耐瘠薄物种增加, 群落内物种间的正关联种对数比例下降, 负关联种对数比例增加;经历了初期4a的剧烈变动后, 群落变化程度减弱, 工程迹地植物群落处于不稳定的恢复阶段。由于原生境被严重破坏, 恢复过程中虽然物种数和盖度增加, 但是土壤环境条件变化缓慢, 群落结构尚不稳定, 应该属于从逆向演替到正向演替的过渡时期。所以在植被自然恢复仍不稳定的状态下, 应尽量降低人为的二次干扰, 避免逆向演替的发生。
致谢: 本文所用数据源于青藏铁路沿线植被多次调查成果, 调查数据是所有参与青藏铁路沿线调查人员的成果, 8年来参与调查的人员共60余位。野外调查得到了青藏铁路公司、国家林业局、西藏林业厅、青海林业厅等单位的大力帮助, 特此致谢。| [1] | 王文进, 张明, 刘福德, 郑建伟, 王中生, 张世挺, 杨文杰, 安树青. 海南岛吊罗山热带山地雨林两个演替阶段的种间联结性. 生物多样性 , 2007, 15 (3) : 257–263. |
| [2] | 钟彦龙, 王银山, 徐敏, 何静, 吕光辉. 艾比湖湿地植物种间关系研究. 干旱区资源与环境 , 2010, 24 (5) : 153–157. |
| [3] | 彭少麟, 周厚诚, 郭少聪, 黄忠良. 鼎湖山地带性植被种间联结变化研究. 植物学报 , 1999, 41 (11) : 1239–1244. |
| [4] | 王慧敏, 庞春花, 张峰, 赵彩莉. 汾河流域中下游湿地植被优势种种间关联性分析. 生态学杂志 , 2012, 31 (10) : 2507–2512. |
| [5] | 王琳, 张金屯. 历山山地草甸优势种的种间关联和相关分析. 西北植物学报 , 2004, 24 (8) : 1435–1440. |
| [6] | 李秋玲, 范庆安, 马晓勇, 高昆, 张峰. 山西黄河湿地植被优势种群种间关系. 生态学杂志 , 2007, 26 (10) : 1516–1520. |
| [7] | 张峰, 张金屯, 韩广业. 历山自然保护区猪尾沟森林群落树种种间关系及环境解释. 植物生态学报 , 2002, 26 (增刊) : 52–56. |
| [8] | 张桂萍, 张峰, 茹文明. 旅游干扰对历山亚高山草甸优势种群种间相关性的影响. 生态学报 , 2005, 25 (11) : 2868–2874. |
| [9] | O'Connor I, Aarssen L W. Species association patterns in abandoned sand quarries. Vegetation , 1987, 73 (2) : 101–109. |
| [10] | 郭逍宇, 张金屯, 高洪文. 白羊草群落优势种种间联结性的分析. 草业学报 , 2003, 12 (2) : 14–19. |
| [11] | 邢福, 郭继勋. 糙隐子草草原3个放牧演替阶段的种间联结对比分析. 植物生态学报 , 2001, 25 (6) : 693–698. |
| [12] | 娄彦景, 赵魁义. 三江平原毛苔草群落近30年演替过程中的种间联结性分析. 生态学杂志 , 2008, 27 (4) : 509–513. |
| [13] | Rotholz E, Mandelik Y. Roadside habitats:effects on diversity and composition of plant, arthropod, and small mammal communities. Biodiversity and Conservation , 2013, 22 (4) : 1017–1031. DOI:10.1007/s10531-013-0465-9 |
| [14] | Caprez R, Spehn E, Nakhutsrishvili G, Körner C. Drought at erosion edges selects for a'hidden'keystone species. Plant Ecology and Diversity , 2011, 4 (4) : 303–311. DOI:10.1080/17550874.2011.600343 |
| [15] | Gutiérrez-Girón A, Gavilán R G. Spatial patterns and interspecific relations analysis help to better understand species distribution patterns in a Mediterranean high mountain grassland. Plant Ecology , 2010, 210 (1) : 137–151. DOI:10.1007/s11258-010-9745-6 |
| [16] | Alex F, Constanza L Q, Lohengrin A C. Spatial patterns in cushion-dominated plant communities of the high Andes of central Chile:How frequent are positive associations?. Journal of Vegetation Science , 2008, 19 (1) : 87–96. DOI:10.3170/2007-8-18332 |
| [17] | 钟宇, 张健, 杨万勤, 吴福忠, 冯茂松, 刘泉波. 巨桉人工林草本层优势种的种间关系及生态种组的划分. 草业学报 , 2010, 19 (3) : 56–62. |
| [18] | 何友均, 崔国发, 邹大林, 郑杰, 董建生, 李永波, 郝万成, 李长明. 三江源玛珂河林区寒温性针叶林优势草本种间联结研究. 北京林业大学学报 , 2008, 30 (1) : 148–153. |
| [19] | 周金星, YangJ, 董林水, 张旭东, 左力. 青藏铁路唐古拉山南段沿线植被多样性及盖度特征分析. 北京林业大学学报 , 2008, 30 (3) : 24–30. |
| [20] | 董林水, 张旭东, 周金星, 李冬雪. 青藏铁路沿线北段植被物种丰富度及盖度的动态变化. 长江流域资源与环境 , 2008, 17 (4) : 551–556. |
| [21] | 马涛, 周金星, 张旭东, 巴特尔·巴克, 李冬雪. 新建青藏铁路沿线各生态区植被分布特征研究初探. 水土保持研究 , 2007, 14 (3) : 150–154. |
| [22] | 房飞, 胡玉昆, 张伟, 公延明, 柳妍妍, 杨秀娟. 高寒草原植物群落种间关系的数量分析. 生态学报 , 2012, 32 (6) : 1898–1907. |
| [23] | 李丹雄, 杨建英, 史常青, 周进, 王兴宇, 王新, 伏凯, 赵廷宁. 汶川地震滑坡迹地植被恢复中优势种的种间关联性. 应用与环境生物学报 , 2014, 20 (5) : 938–943. |
| [24] | 齐麟, 赵福强. 不同采伐强度对阔叶红松林主要树种空间分布格局和物种空间关联性的影响. 生态学报 , 2015, 35 (1) : 46–55. |
| [25] | 孙士云. 青藏铁路沿线的生态环境特点及保护对策. 冰川冻土 , 2003, 25 (增刊1) : 181–185. |
| [26] | 祝广华, 陶玲, 任珺. 青藏铁路工程迹地对植被的影响评价. 草地学报 , 2006, 14 (2) : 160–180. |
| [27] | 周金星, 易作明, 李冬雪, 高甲荣. 青藏铁路沿线原生植被多样性分布格局研究. 水土保持学报 , 2007, 21 (3) : 173–187. |
| [28] | Zhou J X, Yang J, Peng G. Constructing a green railway on the Tibet Plateau:evaluating the effectiveness of mitigation measures. Transportation Research Part D:Transport and Environment , 2008, 13 (6) : 369–376. DOI:10.1016/j.trd.2008.06.002 |
| [29] | 吴征镒. 中国植被. 北京: 科学出版社, 1995642–653. |
| [30] | 高永恒, 曾晓阳, 周国英, 王根绪. 长江源区高寒湿地植物群落主要种群种间关系分析. 湿地科学 , 2011, 9 (1) : 1–7. |
| [31] | 张金屯. 数量生态学.2版. 北京: 科学出版社, 201120–109. |
| [32] | Schluter D. A variance test for detecting species associations, with some example applications. Ecology , 1984, 65 (3) : 998–1005. DOI:10.2307/1938071 |
| [33] | Dice L R. Measures of the amount of ecologic association between species. Ecology , 1945, 26 (3) : 297–302. DOI:10.2307/1932409 |
| [34] | 王伯荪, 彭少麟. 南亚热带常绿阔叶林种间联结测定技术研究. Ⅰ.种间联结测式的探讨与修订. 植物生态学与地植物学丛刊 , 1985, 9 (4) : 274–285. |
| [35] | 周先叶, 王伯荪, 李明光, 昝启杰. 广东黑石顶自然保护区森林次生演替过程中群落的种间联结性分析. 植物生态学报 , 2000, 24 (3) : 332–339. |
| [36] | Pickett S T A, Collins S L, Armesto J J. A hierarchical consideration of causes and mechanisms of succession. Vegetation , 1987, 69 (1) : 109–114. |
| [37] | 杜峰, 徐学选, 张兴昌, 邵明安, 梁宗锁, 山仑. 陕北黄土丘陵区撂荒群落排序及演替. 生态学报 , 2008, 28 (11) : 5418–5427. |
| [38] | 鄢燕, 张锦华, 张建国. 青藏公路沿线高寒草地次生群落特征及生态修复. 草地学报 , 2006, 14 (2) : 156–159. |
| [39] | 杨元武, 李希来. 不同退化程度高寒草甸高山嵩草的构件变化. 西北植物学报 , 2011, 31 (1) : 167–171. |
| [40] | 张继义, 赵哈林. 植被(植物群落)稳定性研究评述. 生态学杂志 , 2003, 22 (4) : 42–48. |
| [41] | 刘秀珍, 张峰, 张金屯, 王琰. 管涔山撂荒地植物群落演替过程中优势种种间关系分析. 山西大学学报:自然科学版 , 2010, 33 (1) : 142–146. |
| [42] | 王伯荪. 植物种群学. 广州: 中山大学出版社, 1989115–120. |
| [43] | 靳瑰丽, 董莉莉, 安沙舟, 何龙, 梁娜, 张梦妮. 天山北坡醉马草群落种间关联分析. 草地学报 , 2014, 22 (6) : 1179–1185. |