 2016, Vol. 36
2016, Vol. 36文章信息
- 王渊, 初红军, 韩丽丽, 陶永善, 布兰, 刘钊, 蒋志刚
- WANG Yuan, CHU Hongjun, HAN Lili, TAO Yongshan, BU Lan, LIU Zhao, JIANG Zhigang
- 野放普氏野马(Equus przewalskii)家域面积及其影响因素
- Factors affecting the home range of reintroduced Equus przewalskii in the Mt. Kalamaili Ungulate Nature Reserve
- 生态学报, 2016, 36(2): 545-553
- Acta Ecologica Sinica, 2016, 36(2): 545-553
- http://dx.doi.org/10.5846/stxb201402090224
-
文章历史
- 收稿日期: 2014-02-09
- 网络出版日期: 2015-06-09






2. 新疆卡拉麦里山有蹄类自然保护区阿勒泰管理站, 阿勒泰市 836500;
3. 中国科学院动物研究所, 北京 100101
2. Altay Management Station, Mt. Kalamaili Ungulate Nature Reserve, Altay 836500, China;
3. Institute of Zoology, Chinese Academy of Science, Beijing 100101, China
普氏野马是准格尔盆地生物多样性的旗舰物种之一,普氏野马重引入中国的工作受到国内外广泛关注[1]。普氏野马群体可分为家族群和公马群,家族群由1匹成年公马、1—6匹甚至更多成年母马和幼驹组成;公马群则由成年公马及被驱逐出家族的亚成年公马组成,群体数量变化较大。普氏野马是家族性动物,家族成员在相对固定的家域(Home range)中活动。家域大小与季节性水源、食物丰富度和温度等因素有关[2, 3, 4, 5, 6]。家域的形状、面积及其变化规律以及家域与种群密度之间的关系是动物生态学研究的重要内容[7]。群体年龄组成及与年龄有关的社群行为是影响家域大小的主要因素[8]。
2001年8月28日,在新疆卡拉麦里山有蹄类自然保护区(简称卡山保护区)北缘国道216线310km西侧吉列库都克区域首次进行了27匹普氏野马野化放归自然试验的软放归。随后,人们相继开展了野放状态下普氏野马野化过程及食性、应激和社群等研究[9, 10, 11, 12]。由于吉列库都克区域距离国道216线较近,自野马野放后到2009年发生了多起野马被车撞的事故[13]。为了避免事故再次发生,通过考察论证,选定卡拉麦里山西南部乔木希拜区域为新野放点。于2009年初将野马迁移至乔木希拜野放点。目前,放归自然的普氏野马基本适应了卡山保护区生境,成功的在野外产下了第2代。种群数量不断增加,至2012年底野放野马数目增加到96匹。
家域(Home range)一词由Burt于1943提出,指动物日常摄食、休息、繁殖、哺育幼体和群体成员互作的区域,是生物因素和非生物因素综合影响的综合体。家域的形状、面积及其变化规律,以及家域与种群密度之间的关系是动物生态学研究的重要内容[7]。对于主要营群体生活的野放普氏野马来说,影响家域面积大小不断变化的原因复杂,因此,有必要对野放普氏野马的家域变化进行详细的研究,以期为卡山保护区乃至新疆荒漠区域的野生动物保护工作提供科学的指导[12]。
本研究检验了野马家族群数量影响其家域面积,期望通过本研究回答如下问题:(1)年度间和季节间野放野马家域有何变化?(2)影响野放野马家域有哪些因子?
1 研究区域概况卡山保护区乔木希拜区域(88°40′—89°10′E,45°5′—45°20′N)是目前野放普氏野马的主要栖息区,面积约2500 km2,位于卡山保护区西南缘,海拔高度在800—1200 m之间。野放点北部为砾漠和固定沙丘,植被盖度约为30%左右,东面与216国道相邻,西面与彩南油气田相望,南靠卡拉麦里山北缘。地形复杂多变,荒漠植被呈斑块状分布,其中石质山丘植被盖度接近于0,而低洼处的黄泥滩周边植被盖度可达30%—40%。全年降水量159.1 mm,而蒸发量为2090 mm,气候异常干燥。6—9月降水量少且气候炎热,日均气温20—30 ℃,最高温度可达50 ℃以上;9—10月为秋季,气温逐渐下降,到11月上旬即可降至0 ℃以下,最冷月份平均气温在-20 ℃左右,最低气温可达-38 ℃,次年3月底4月初气温逐步上升。
野放区有国家I级重点保护野生动物13种,国家Ⅱ级重点保护野生动物36种,如蒙古野驴(E. hemionus)、盘羊(Ovis ammon)、鹅喉羚(Gazella subgutturosa)、猞猁(Felis lynx)、狼(Canis lupus)以及多种猛禽等。优势植物类群有藜科(Chenopodiaceae)、菊科(Asteraceae)、禾本科(Poaceae)、豆科(Leguminosae)、莎草科(Cyperaceae)、蓼科(Polygonaceaee)、麻黄科(Ephedraceaee)等共计31科139种[14]。
2 研究方法 2.1 调查方法因为长期野外监测,野放普氏野马熟悉监测人员,故监测人员在野外能靠近并辨识野马臀部的烙印编号和个体特征,实测野放野马的GPS点位,记录每个家族群的头马和带驹的母马体况、活动现况以及附近的其它野生有蹄类动物如蒙古野驴、鹅喉羚等。同时记录海拔、温度、湿度、风速、水源的盐碱度、与最近水源地的距离、与人类活动地的距离、草本和灌木类型、草本覆盖度和灌木覆盖度等参数。
本文使用的野放普氏野马行为名词定义如下:(1)夺群,成年公马对家族群的头马(公马)发起挑战,通常表现为向头马鸣叫、追打、撕咬、踢,如果胜利则夺取了该家庭群头马控制的所有母马和幼驹,组成新的家庭群,而失败的头马则被驱逐,或夺取家庭群头马的一部分母马和幼驹并形成两个家庭群。(2)圈群,家庭群中公马围绕家庭群低头快速跑动,圈拢群体成员并同时警示其他群体的公马,在夏季水源地饮水期间此种行为较常见。(3)护群,头马为保护自己的群体成员,而与其它家庭群的头马或公马群成员的打斗行为。
利用GIS中的Intersect analysis模块分别计算了2011年与2012年、年度间及季节间不同野放野马群体的年际间及季节间家域重叠。
2.2 数据处理采用最小凸多边形法(MCP)[15]计算不同年份、不同季节和不同群体野放普氏野马家域面积:
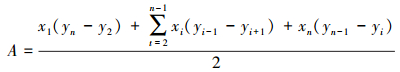
式中,A为家域面积,xi和yi分别是第i个野放野马群体位点的经度和纬度。利用ArcGIS9.3的Spatial Analyst Tools模块进行分析。
2.3 统计分析利用SPSS17.0 for Windows处理数据,采用单因素方差分析野放野马群体的家域及家域重叠;以家族群大小为协变量进行了野放野马家域的协方差分析。对野放野马不同季节家域面积差异、2011年与2012年不同群体的家域重叠以及不同年份不同季节之间的家域重叠值进行LSD多重比较。对影响野放野马家域的生境因子进行了主成分分析。
3 结果 3.1 野放普氏野马的群体及构成根据2011年与2012年的跟踪观测,记录到放归普氏野马的9个群体及各群体的构成(图 1)。野放成马和幼驹比例为分别为5.23∶1和4.53∶1,2011年与2012年野放野马雄雌性比分别为0.65∶1和1∶1.02。

|
| 图 1 2012年野放野马家族群体结构 Fig.1 Family population structures of released Przewalski′s horses in 2012 |
通过2011至2012年每年3月到11月的长时间观测,得出每个野马群均有较为固定的活动区域,但各群野马的活动范围存在年际间的较大变化。
以年度为重复样本,单因素方差分析(One-Way ANOVA)表明,不同野马群的家域面积差异显著(F=4.235,P=0.036<0.05)(表 1)。家域面积与群体大小显著相关(r=0.608,P=0.019<0.05),与线性回归模型y=12.168x+225.161吻合(R2=0.370,F=8.217,P=0.012<0.05)(图 2)。2011年,每匹野放野马平均家域面积为(20±2) km2,2012年上升为(30±2) km2,家域面积与家族大小显著相关(P<0.05)。
| 群 Families | 家域面积Home range sizes/km2 | |||||||
| 春季Spring | 夏季Summer | 秋季Autumn | 全年Whole year | |||||
| 2011 | 2012 | 2011 | 2012 | 2011 | 2012 | 2011 | 2012 | |
| *公马群 | ||||||||
| No.1 | 217 | 291 | 95 | 70 | 137 | 180 | 332 | 383 |
| No. 2 | 385 | 305 | 93 | 63 | 227 | 181 | 387 | 424 |
| No. 3 | 243 | 210 | 56 | 52 | 141 | 131 | 286 | 285 |
| No. 4 | 362 | 172 | 89 | 67 | 213 | 99 | 376 | 248 |
| No. 5* | 463 | 342 | 139 | 100 | 303 | 236 | 481 | 463 |
| No. 6 | 254 | 230 | 68 | 57 | 157 | 135 | 289 | 339 |
| No. 7 | 222 | 266 | 47 | 56 | 130 | 151 | 297 | 401 |
| No. 8 | 135 | 68 | 84 | 189 | ||||
| No. 9 | 169 | 65 | 90 | 260 | ||||
| 均值Mean | 307 | 219 | 84 | 66 | 187 | 136 | 335 | 310 |
| 标准差 Standard Deviation | 96 | 70 | 31 | 14 | 64 | 50 | 71 | 92 |

|
| 图 2 野放普氏野马家族群大小与家域面积关系 Fig.2 Relationship between family sizes and home range areas of the released Przewalski′s horses in Qiaomuxibai |
多因素嵌套设计方差分析表明,不同季节间野马家域面积差异显著(F=15.875,df=2,P=0.025<0.05);同一季节不同年份间野马家域面积差异显著(F=2.730,df=3,P=0.056>0.05)。单因素方差分析表明,2011年不同季节野马家域面积差异显著(F=18.283,df=2,P<0.001),2012年野马不同季节家域面积差异显著(F=25.417,df=2,P<0.001)。LSD多重比较表明,无论是2011年还是2012年,春季野马家域面积显著大于其他季节(P<0.001),秋季显著大于夏季(P<0.001)。
3.4 野放普氏野马家域重叠分析单因素方差分析 (One-Way ANOVA) 表明,2011年各野马群两两之间家域重叠面积无显著差异(F=1.238,df=6,P=0.311>0.05)。相关性分析表明,家域重叠面积与群大小不相关(r=0.256,P=0.580>0.05)。2012年各个野马群两两之间的家域重叠面积有显著差异(F=4.521,df=8,P<0.001)。家域重叠面积与群大小显著相关(r=0.706,P=0.033<0.05),与线性回归模型 y=3.826x+23.429吻合(R2=0.499,F=6.973,P=0.033<0.05)。独立样本T检验表明,2011年各个野马群两两之间的家域重叠面积显著大于2012年(t=2.543,P=0.012<0.05)(表 2,表 3)。
| 群号 Population number | 2 | 3 | 4 | 5 | 6 | 7 |
| 1 | 92 | 40 | 21 | 93 | 53 | 66 |
| 2 | 52 | 60 | 206 | 101 | 85 | |
| 3 | 18 | 523 | 44 | 39 | ||
| 4 | 61 | 37 | 41 | |||
| 5 | 101 | 90 | ||||
| 6 | 70 |
| 群号 Population number | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
| 1 | 143 | 71 | 34 | 127 | 77 | 81 | 36 | 60 |
| 2 | 68 | 34 | 140 | 77 | 82 | 36 | 59 | |
| 3 | 33 | 65 | 65 | 53 | 34 | 30 | ||
| 4 | 51 | 38 | 24 | 12 | 3 | |||
| 5 | 81 | 79 | 35 | 58 | ||||
| 6 | 56 | 34 | 32 | |||||
| 7 | 35 | 55 | ||||||
| 8 | 24 |
以年份为重复样本,单因素方差分析表明(One-Way ANOVA),单个野马群季节间家域重叠面积差异显著(F=5.695,df=8,P<0.001)。LSD多重比较表明,5、7和8号群的家域季节间重叠面积较大(P<0.05),3、6和9号群较小(P<0.05)。Pearson相关性分析表明,不同野马群年均家域重叠面积与群大小不相关(r=0.162,P=0.548>0.05)。单因素方差分析表明,野马2011年不同季节间家域重叠面积无显著差异(F=0.575,df=2,P=0.573>0.05);而2012年不同季节有显著差异(F=4.148,df=2,P=0.028>0.05)。独立样本T检验表明,2012年夏秋季野马家域重叠面积显著小于春秋季的(t=-2.401,df=16,P=0.029<0.05),2012年春夏季与夏秋季(t=0.464,df=16,P=0.649>0.05)、春夏季与春秋季(t=-2.038,df=16,P=0.058>0.05)均无显著差异(表 4)。
| 群号 Families | 家域Home ranges/km2 | |||||
| 春-夏Spring-Summer | 夏-秋Summer-Autumn | 春-秋Spring-Autumn | ||||
| 2011年 | 2012年 | 2011年 | 2012年 | 2011年 | 2012年 | |
| 1 | 24 | 29 | 61 | 33 | 27 | 106 |
| 2 | 63 | 43 | 63 | 45 | 109 | 149 |
| 3 | 9 | 24 | 40 | 24 | 17 | 49 |
| 4 | 13 | 80 | 27 | 57 | 20 | 81 |
| 5 | 119 | 104 | 130 | 92 | 166 | 109 |
| 6 | 36 | 36 | 41 | 23 | 54 | 30 |
| 7 | 76 | 79 | 76 | 73 | 132 | 90 |
| 8 | 47 | 46 | 196 | |||
| 9 | 20 | 15 | 32 | |||
2011年,影响野放野马家域生境因子第一主成分特征值为1.614,贡献率为14.672%(表 5)。第一个主成分中春季绝对值较大的权系数是草本盖度、温度、湿度等生态因子,夏季绝对值较大的权系数是湿度、最近水源地距离和草本盖度等生态因子,秋季绝对值较大的权系数是草本盖度、风速和最近水源地距离等生态因子。说明野放野马家域的大小与温度、风速、湿度、草本盖度及最近水源地距离5个因子有着密切的关系。因此,影响2011年度野放野马家域面积的生物因子包括草本盖度,而非生物因子包括温度、风速、湿度、最近水源地距离等。
| 季节Season | 变量Variable | 主成份Main component | ||||
| 1 | 2 | 3 | 4 | 5 | ||
| 春季Spring | 草本盖度/% | 0.493 | 0.524 | 0.363 | 0.184 | 0.389 |
| 温度/℃ | 0.627 | 0.218 | -0.182 | -0.283 | 0.176 | |
| 湿度/ | 0.422 | 0.465 | 0.238 | -0.197 | 0.234 | |
| 夏季Summer | 温度/℃ | 0.627 | 0.218 | -0.182 | -0.283 | 0.176 |
| 湿度/ | 0.479 | 0.445 | 0.138 | -0.097 | 0.289 | |
| 最近水源距离/km | 0.677 | 0.463 | 0.372 | 0.523 | 0.345 | |
| 秋季Autumn | 草本盖度/% | 0.673 | 0.524 | 0.363 | 0.184 | 0.389 |
| 风速/(m/s) | 0.452 | 0.401 | 0.123 | 0.107 | 0.220 | |
| 最近水源距离/km | 0.472 | 0.412 | 0.362 | 0.503 | 0.188 | |
| 特征值 | 1.614 | 1.443 | 1.180 | 1.120 | 1.048 | |
| 贡献率/% | 14.672 | 13.119 | 20.90 | 18.380 | 8.314 | |
| 累计贡献率/% | 14.672 | 27.791 | 48.695 | 67.075 | 75.389 | |
2012年影响野放野马家域的主成分特征值结果与2011年的主成分分析结果相一致(表 6)。成对样本相关性分析发现温度与草本盖度对家域面积大小的影响呈显著性差异(P=0.0037<0.01)。因此,影响2012年度野放野马家域面积的生物因子包括草本盖度和灌木盖度,而非生物因子包括温度、风速、湿度、最近水源地距离等。
| 季节Season | 变量Variable | 主成份Main component | ||||
| 1 | 2 | 3 | 4 | 5 | ||
| 春季Spring | 草本盖度/% | 0.619 | 0.335 | 0.299 | 0.070 | 0.039 |
| 温度/℃ | 0.523 | -0.465 | -0.393 | -0.399 | -0.223 | |
| 湿度/ | 0.540 | 0.278 | -0.116 | 0.260 | 0.157 | |
| 夏季Summer | 温度/℃ | 0.527 | 0.334 | -0.482 | -0.345 | 0.256 |
| 最近水源距离/km | 0.521 | 0.422 | 0.413 | 0.332 | 0.315 | |
| 湿度/ | 0.540 | 0.278 | -0.116 | 0.260 | 0.163 | |
| 秋季Autumn | 草本盖度/% | 0.611 | 0.427 | 0.383 | 0.229 | 0.336 |
| 最近水源距离/km | 0.502 | 0.645 | 0.296 | -0.279 | -0.237 | |
| 灌木盖度/% | 0.549 | 0.517 | -0.101 | -0.162 | 0.107 | |
| 特征值 | 2.180 | 1.576 | 1.350 | 1.243 | 1.038 | |
| 贡献率/% | 18.163 | 13.136 | 21.612 | 16.246 | 6.805 | |
| 累计贡献率/% | 18.163 | 31.299 | 52.910 | 69.156 | 75.961 | |
Pearson相关性双侧检验,2011年的家域面积与最近隐蔽物的距离及湿度呈现正相关,表明两者与家域面积的大小呈相关的线性变化,随着湿度的增加家域面积表现为不断增大。而与最近居民点距离、风速、温度呈现负相关,且最近居民点距离和温度在(P<0.01)水平上呈显著性负相关,表明最近居民点距离和温度是野马家域的限制性因子。其次,Pearson相关性双侧检验得,2012年家域面积与草本盖度、最近居民点距离、温度成显著性负相关,其中草本盖度在(P<0.01)水平上呈显著性相关。表明2012年草本盖度是影响家域大小的主要因素之一。此外,最近居民点距离和温度也是影响家域大小的主要因素。
4 讨论通过对卡山保护区野放野马家域的研究,有助于进一步了解大中型荒漠有蹄类的生境要求,在规划恢复灭绝物种重引入种群时充分考虑各种限制因子。
本研究支持野马群体数量决定其家域面积的假说。随着野放马种群密度的增加,气象、植被盖度和水源地等环境因素的年际变化,为了维持群体成员的生存,野放野马采食等活动地域增大,竞争强度增大,家域也随之变大。此外,家域的形成还是多因素的组合体或是多因素的妥协。研究中1号群、2号群和3号群家域面积变化并没有呈现出较强规律性,但群体数量的增加导致其家域面积的增加是显而易见的,如:2号群在2012年比2011年多产3驹,其家域面积从387 km2增加到424 km2。可能的解释为随着栖息地内家族成员的增加,其家族需要的采食等活动面积增大,因此,家域面积扩大。而Krasinska[16]等人的研究结果发现偶蹄类家族群中幼体及亚成体数目的增加,导致了家族群体稳定性的降低,从而导致了家域面积的增加或减少。
Miller[17]发现美国野化家马家域面积在73km2至303km2之间,本研究中野马的家域面积则为47km2至463km2。Miller的研究结果未考虑公马群、野马群体性别比例和组成。卡山保护区野放野马公马群家域面积无论是在2011年还是在2012年都显著大于家族群。究其原因,其一,卡山保护区野放野马公马群成员都是成年个体,个体大,移动速度快。其二,野放野马公马群需要的采食等活动面积大,而家族群的幼驹可能影响了整个野放群体的移动速度。其三,公马群中社会关系较家族群单一,在繁殖期没有“圈群”、“护群”、“头马之间的打斗”等社会行为。有蹄类、鸟类、鼠类、猫科和犬科等雄性家域面积往往比雌性的大,乔木西拜区域的5号雄性群的家域比其它野马家族群的家域大。研究发现在繁殖期雄性家域较大[18, 19, 20],野放雄性野马需要通过扩大家域寻找繁殖机会。
季节对动物家域面积有一定的影响[21]。不同季节家域的变化体现了野放野马对环境变化的适应。一般来讲,春季野放野马家域面积最大,其次为秋季,夏季最小。春季,野放野马搜索采食嫩草导致其家域面积增大,而夏秋季该区域的高温和水是制约野放普氏野马活动的主要因素,与本文的主成分分析的研究结果一致。在荒漠草原温度是影响野放野马家域大小的最主要因素[22, 23]。6—9月酷热的夏季和夏末秋初期间,野放野马和蒙古野驴等争夺水源;而春季野放野马则与蒙古野驴等争夺草场[12],这可能是春季和秋季野放野马家域较夏季大的一个原因。由于荒漠区夏季极度干旱少雨且温度的持续升高,导致动物需水量的增加,减少了动物活动范围和采食时间,在一定意义上导致了野放野马家域面积的减少[24, 25]。而野放野马选择性采食[26],且喜欢在针茅等密度较大区域采食,这又是导致野放野马家域面积减小的另一个原因[27]。
卡山保护区的生态环境季节性变化强烈。家域变化规律与家域内食物质量和数量的季节性变化相关,因此野放野马家域随季节的变化。本研究与Berger[28]对野马的研究有很大差别,Berger的研究证明野放野马在野放地附近活动,其家域面积不到50 km2。并且水源充足时四季野马家域面积变化较小。
早先人们很少认识到动物的个性。现在研究表明,由于进化的连续性,所有动物都有个性。人们在许多动物中研究了动物个性特征,如胆量、活动量和攻击性的个体差异[29]。由于生活史特征是常见的生物学特征,并可能促进个体间的行为差异。Biro和Stamps[30]推测,如果动物个性特征的稳定差异能够使动物个体的产生稳定生产力性状(生长/繁殖力)个体差异的话,那么动物个性特征的稳定差异将在自然选择中被固定下来。头马个性的差异是否影响其进化适合度(Evolutionary fitness)值得进一步研究。
由于野放初期野放区域野马密度低,野马与同域分布的蒙古野驴等有蹄类竞争较小。小的野放野马家族群在食物充足的情况下没有必要增加家域面积[31]。随着野放野马种群数量的增加,水、食物和隐蔽场所以及空间再分配等各种因素的影响,野放普氏群体之间竞争加剧,导致了野放野马家域扩大,这与King[31]在蒙古国Hustai National Park的研究报道相一致。
Kaczensky等[24]在蒙古国戈壁B保护区的研究发现野放野马和蒙古野驴等有蹄类同域分布,导致这种同域分布的主要原因是食物和水资源的变化。这与同域原麝和斑羚种群的研究结果相一致[32]。可以推测,随着野放野马种群数量增加,单位面积马匹密度增大,导致对荒漠草原和水源地等资源的过度利用,加大了野放野马与蒙古野驴、盘羊和鹅喉羚的竞争。
地形地貌可能对不同季节野放野马家域有一定的影响。研究表明,提高所在环境的郁闭度是增高野生动物安全感的途径之一[33, 34, 35]。选择高郁闭度的生境是影响野放野马家域的一个因素,例如:生产不久的母马,可能利用该方法来提高对幼驹的保护,从而导致活动区域的减少或增加。
没有证据表明野放普氏野马具有严格的领地性[36, 37],野放野马不能或无法限制其它家族群进入其家域活动,多项观察研究表明不同野放野马群体在同一区域采食[38, 39, 40]。然而,不同家族群体的组成差异可能会导致不同群体家域重叠程度差异,而各家族群头马个性差异也可能导致这种差异。目前,尚不能得出是哪一方面因素占主导地位。
随着区域内野放野马数量增加,原先家域重叠度较高的家族向外扩大家域。此外,随着野放区域野马家族群和个体数量增加,各野马群的的活动范围及重叠变化较大。2012年,随着群体增多、个体数量增加,野放野马活动范围不断增大。家域重叠面积与群体的大小显著相关。水源地可能影响家域面积,在野放区只有少数几处永久水源地的前提下,群体大小与家域的重叠有显著性差异。
致谢:艾代、蒋峰、阿尔成、叶尔江、木合买提、哈力等参加野外工作,曹青博士和张永军博士提供帮助,特此致谢。
| [1] | 蒋志刚. 普氏野马(Equus przewalskii). 动物学杂志, 2004, 39(2):100-101. |
| [2] | McIntyre N E, Wiens J A. Interactions between landscape structure and animal behavior:the roles of heterogeneously distributed resources and food deprivation on movement patterns. Landscape Ecology, 1999, 14(5):437-447. |
| [3] | Prange S, Gehrt S D and Wiggers E P. Influences of anthropogenic resources on Raccoon(Procyon Lotor) movements and spatial distribution. Journal of Mammalogy, 2004, 85(3):483-490. |
| [4] | Eide N E, Jepsen J U, Prestrud P. Spatial organization of reproductive Arctic foxes Alopex lagopus:responses to changes in spatial and temporal availability of prey. Journal of Animal Ecology, 2004, 73(6):1056-1068. |
| [5] | Devault T L, Reinhart B D, Brisbin I L, Rhodes O E Jr. Home ranges of sympatric Black and Turkey Vultures in South Carolina. Condor, 2004, 106(3):706-711. |
| [6] | Wauters L A, Bertolino S, Adamo M, Dongen S V, Tosi G. Food shortage disrupts social organization:The case of red squirrels in conifer forests. Evolutionary Ecology, 2005, 19(4):375-404. |
| [7] | Kaczensky P, Ganbaatar O, von Wehrden H, Walzer C. Resource selection by sympatric wild equids in the Mongolian Gobi. Journal of Applied Ecology, 2008, 45(6):1762-1769. |
| [8] | 刘群秀, 王小明, 王正寰. 藏狐家域功能分区及其生物学特性比较. 兽类学报, 2009, 29(4):432-438. |
| [9] | 曹婷婷, 胡德夫, 于小杰, 曹杰, 李凯, 张峰. 冬季圈养和放归普氏野马成年雄性应激状态研究. 干旱区研究, 2009, 26(2):234-237. |
| [10] | 张峰. 普氏野马行为节律及其影响因子研究[D]. 北京:北京林业大学, 2010. |
| [11] | 黄艳, 初红军, 兰文旭, 时坤, 陶永善, 邵长亮. 重引入普氏野马、蒙古野驴和鹅喉羚的秋季营养生态位. 干旱区研究, 2011, 28(6):1045-1050. |
| [12] | 刘姝, 初红军, 王渊, 陶永善, 邵长亮, 刘冬志. 普氏野马(Equus przewalskii)重引入区域的社区保护意识调查分析. 干旱区研究, 2013, 30(1):135-143. |
| [13] | 张峰, 胡德夫, 陈金良, 曹婷婷. 为放归野马开辟安全通道. 大自然, 2008,(3):14-16. |
| [14] | 初红军, 蒋志刚, 兰文旭, 王臣, 陶永善, 蒋峰. 蒙古野驴、鹅喉羚和家畜的食物重叠. 动物学报, 2008, 54(6):941-954. |
| [15] | Hayne D W. Calculation of size of home range. Journal of Mammalogy, 1949, 30(1):1-18. |
| [16] | Kaczensky P, Enkhsaihan N, Ganbaatar O, Walzer C. Identification of herder-wild equid conflicts in the Great Gobi B Strictly Protected Area in SW Mongolia. Exploration into the Biological Resources of Mongolia, 2007, 10:99-116. |
| [17] | Miller R. Seasonal movements and home ranges of feral horse bands in Wyoming's Red Desert. Journal of Range Management, 1983, 36(2):199-201. |
| [18] | Goltz D M, Hess S C, Brinck K W, Danner R M. Home range and movements of feral cats on Mauna Kea, Hawai'i. Pacific Conservation Biology, 2008, 14:177-184. |
| [19] | Swingle J K, Foreman E D. Home range areas and activity patterns of red tree voles(Arborimus longicaudus) in Western Oregon. Northwest Science, 2009, 83(3):273-286. |
| [20] | Morris A D, Miller D A. Conner L M. Home-range size of evening bats(Nycticeius humeralis) in Southwestern Georgia. Southeastern Naturalist, 2011, 10(1):85-94. |
| [21] | 周江, 李小成, 周照骊, 韩文涛, 陈升华. GIS技术在海南长臂猿保护中的运用. 贵州师范大学学报:自然科学版, 2011, 24(4):439-442. |
| [22] | Kaczensky P, Ganbaatar O, Altansukh N. Status of the Przewalski's horse population in the Great Gobi B Strictly Protected Area in SW Mongolia in 2012. |
| [23] | Zweifel-Schielly B, Kreuzer M, Ewald K C, Suter W. Habitat selection by an Alpine ungulate:the significance of forage characteristics varies with scale and season. Ecography, 2009, 32(1):103-113. |
| [24] | Kaczensky P, Sheehy D P, Johnson D E, Walzer C, Lkhagvasuren D, Sheehy C M. Room to Roam? The threat to Khulan(Wild Ass) from Human Intrusion. Mongolia Discussion Papers, East Asia and Pacific Environment and Social Development Department. World Bank, Washington, D. C. 2006. |
| [25] | Kaczensky P, Ganbaatar O, Altansukh N. Status of the Przewalski's horse population in the Great Gobi B Strictly Protected Area in SW Mongolia in 2012. |
| [26] | Duncan P. Horses and Grasses:The Nutritional Ecology of Equids and Their Impact on the Camargue. New York:Springer, 1992. |
| [27] | Salter R, Hudson R. Feeding ecology of feral horses in Western Alberta. Journal of Range Management, 1979, 32(3):221-225. |
| [28] | Berger J. Wild Horses of the Great Basin:Social Competition and Population Size(Wildlife Behavior and Ecology). Chicago:University of Chicago Press, 1986. |
| [29] | Dall S R X, Houston A I, McNamara J M. The behavioural ecology of personality:consistent individual differences from an adaptive perspective. Ecology Letters , 2004, 7(8):734-739. |
| [30] | Biro P A, Stamps J A. Are animal personality traits linked to life-history productivity?. Trends in Ecology and Evolution, 2008, 23(7):361-368. |
| [31] | King S R B. Home range and habitat use of free-ranging Przewalski horses at Hustai National Park, Mongolia. Applied Animal Behaviour Science, 2002, 78(2/4):103-113. |
| [32] | 王文, 张静, 马建章, 刘海波. 小兴安岭南坡野猪家域分析. 兽类学报, 2007, 27(3):257-262. |
| [33] | Keiper R. Social organization of feral ponies. Proc Penn Acad Sci, 1976, 50:69-70. |
| [34] | Keiper R R, Berger J. Refuge-seeking and pest avoidance by feral horses in desert and island environments. Applied Animal Ethology, 1982, 9(2):111-120. |
| [35] | Wallis de Vries M F, Manibazar N, Dügerlham S. The vegetation of the forest-steppe region of Hustain Nuruu, Mongolia. Vegetatio, 1996, 122(2):111-127. |
| [36] | Catt D C, Staines B W. Home range use and habitat selection by red deer(Cerrus elaphus) in a sitka spruce plantation as determined by radio-tracking. Journal of Zoology, 1987, 211(4):681-693. |
| [37] | Cederlund G N, Okarma H. Home range and habitat use of adult female moose. Journal of Wildlife Management, 1988, 52(2):336-343. |
| [38] | Gates S. A study of the home ranges of free-ranging Exmoor ponies. Mammal Review, 1979, 9(1):3-18. |
| [39] | Rubenstein D I. Behavioural ecology of island feral horses. Equine Veterinary Journal, 1981, 13(1):27-34. |
| [40] | Linklater W L. Adaptive explanation in socio-ecology:lessons from the Equidae. Biological Reviews, 2000, 75(1):1-20. |