 2016, Vol. 36
2016, Vol. 36文章信息
- 文欣, 刘宝宁, 周成旭, 蒋莹, 陈航霞
- WEN Xin, LIU Baoning, ZHOU Chengxu, JIANG Ying, CHEN Hangxia
- 颗石藻Pleurochrysis carterae抗捕食特征
- Characterization of anti-predator abilities of Pleurochrysis carterae
- 生态学报, 2016, 36(2): 525-534
- Acta Ecologica Sinica, 2016, 36(2): 525-534
- http://dx.doi.org/10.5846/stxb201401170133
-
文章历史
- 收稿日期: 2014-01-17
- 网络出版日期: 2015-06-10






颗石藻(Coccolithophores)是一类单细胞的光合自养钙化微藻,属定鞭藻,是现代海洋中继硅藻和甲藻之后生物量最大的浮游植物,成为海洋初级生产力的重要组成部分。广泛分布于沿海、大洋甚至内陆咸水区域,可形成大面积高密度水华[1]。其细胞外被CaCO3晶体构成了深海碳酸盐重要来源[2],在海洋生态环境、生物地化循环、生物地层学和古海洋学等方面有重要意义[3]。
Pleurocrysis carterae是一类常发于沿海和围塘养殖环境的一种颗石藻,易快速增殖至高度密集状态,可形成浮沫状恶性水质。实验研究表明,P.carterae是显著引起卤虫死亡的颗石藻种类之一[4, 5, 6]。
随着研究技术进步,浮游植物的抗捕食防御现象及其机制研究得以深入开展[7]。捕食是微藻种群增殖的主要压力之一,为了抵抗捕食或调整因捕食而造成的种群数量减少,微藻进化出多种防御捕食策略。例如,被捕食时,微藻主动聚群形成多细胞链或藻落,以物理方式增加捕食者的捕食难度[8];微藻还可迅速启动针对捕食者的伤激防御机制[9]、产生毒素致死捕食者[10]、产生化学他感物质使捕食者转移捕食方向等[11];微藻也可以通过提高大量增殖能力以应对捕食压力下种群数量的减少[12]。
颗石藻P.carterae种群增殖过程中都处于游动单细胞,且捕食压力存在时不发生物理聚集。研究表明,颗石粒(coccolith)并不是其致死卤虫或防御捕食的原因[4, 5]。Moheimani等认为,人工培养颗石藻P.carterae时不易受原生动物捕食,很大程度上可能源于其产生的抗捕食防御物质[13]。目前尚未发现颗石藻P.carterae具有明显的毒素。研究得最多的颗石藻化学防御物质是DMSP [14-15]。颗石藻P.carterae属DMSP高产种类,我们早期研究也显示,DMSP可能在颗石藻P.carterae致害卤虫中发挥作用[16]。但是,由于DMSP在微藻中意义多种多样,且发挥作用时与群落组成、环境条件、生化代谢过程等多种因素相关[17],这个领域尚需大量研究。
研究微藻抗捕食机制,最理想的是建立捕食者与被捕食者模式系统。卤虫(Artemia salina)是一个理想的模式生物,易于获取及保存,方便培养,个体小且捕食能力强,常被用于毒理学研究及作为捕食者进行生物种间关系的研究[18]。培养卤虫用的微藻多种多样,其中球等鞭金藻(Isochrysis galbana)是一种卤虫适宜饵料,与颗石藻同属于定鞭藻,但两者对卤虫而言显然差异显著。
叶绿素荧光参数的变化是迅速表征微藻受到胁迫的重要参数[19]。脂类代谢与微藻重要的生物化学过程密切相关,特别是微藻质膜糖脂可能在微藻抗捕食防御中发挥作用[20, 21]。
为了研究颗石藻P.carterae抗捕食防御能力、特征及其机制,本文以卤虫为捕食者,以卤虫饵料金藻I.galbana为对照微藻,分析了颗石藻P.carterae对卤虫捕食能力的影响、捕食压力下微藻的叶绿素荧光以及脂组响应特征变化。由于磷对颗石藻的生理及其脂类生化代谢有重要影响[22],本文还研究了磷元素影响下,指标物质DMSP的变化以及是否因此对卤虫存活能力产生影响。
1 材料与方法 1.1 藻种培养与卤虫孵化颗石藻(P.carterae,NMBjih026)、球等鞭金藻(I.galbana,NMBjih-021-2)均来自宁波大学微藻种质库。实验前用f/2培养液预培养在温度20 ℃、光强 2200 lx(D∶L=12 h∶12 h)培养室中。
卤虫卵为本实验室4 ℃冰箱中低温保存种(< 1a),孵化前将卤虫卵置于常温下2 h。在盛有150 mL过滤海水的烧杯中进行孵化。温度30 ℃,盐度25‰,pH为8.0±0.2,光照孵化。
1.2 颗石藻P.carterae对卤虫摄食率的影响实验在6孔组织培养板上进行。所用卤虫为饵料金藻喂养的15日龄成虫。实验前将卤虫置于消毒海水中饥饿处理24h,用消毒海水清洗3次以清除体外附着的微藻。
实验组设置 每孔分别加入10 mL以不同细胞数比例混合的金藻(I)和颗石藻P.carterae(P)藻液, 颗石藻P.carterae的比例逐渐增加(I/P比例分别为10∶0,8∶2,5∶5,2∶8,0∶10),混合总藻细胞密度约1×105 个/mL。每孔加入5只大小相同的活跃成虫。以消毒海水作为饥饿对照。各3个平行。所有培养板均置于温度22 ℃,光照2200—3000 lx(D∶L=12 h∶12 h)的培养条件下。
取样时间组设置和检测方法 按照上述实验组,相同设置5、24 h取样时间组。实验开始后,对各时间点各实验组,观察卤虫的活动敏捷度,记录卤虫的存活数,取出全部卤虫,于消毒海水中清洗3次以去除体表藻液。迅速冷冻备测肠道中的叶绿素。
卤虫存活率
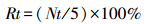
式中,Rt为t时间点的卤虫存活率,Nt为t时间点的存活卤虫数。
卤虫捕食率测定 通过测定叶绿素a的含量来表示卤虫肠道中藻细胞色素含量,表征其捕食能力的大小。叶绿素测定基于Arnon的方法[23],将卤虫样品避光解冻,置于玻璃离心管中,定量添加90% 丙酮,并以移液器头捣碎虫体,移液器头保留在玻璃管中,4 ℃避光密闭萃取24h。离心取上清液,以90% 丙酮为空白,于酶标仪上分别检测630 nm和663 nm处的吸光值(OD630和OD663)。以下列公式计算叶绿素含量,n各实验组表示卤虫的总只数。
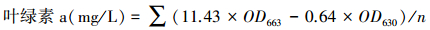
采用单因素方差分析方法进行差异显著性分析。
1.3 捕食压力对颗石藻P.carterae生理生化的影响 1.3.1 捕食压力下,颗石藻P.carterae的叶绿素荧光参数ETR和Fv/Fm的变化实验在250 mL锥形瓶中进行。藻液体积200 mL,密度约1×105 个/mL的颗石藻P.carterae或饵料等鞭金藻指数期藻液。藻液中投入10只卤虫成虫,另以不加卤虫的颗石藻P.carterae和饵料等鞭金藻为非捕食对照组。各组3平行。所有锥形瓶均置于22 ℃、2200 lx、全光照培养条件下。分别在实验起始和24 h后,用叶绿素荧光仪(Water-PAM)检测各组叶绿素荧光参数ETR和Fv/Fm。各组测样4次,取平均值。
1.3.2 捕食压力下,颗石藻P.carterae的脂组变化实验组所用藻液体积为4000 mL,指数后期颗石藻P.carterea和饵料金藻,密度分别为3×105 个/mL和1.8×106 个/mL。所有锥形瓶均置于22 ℃、2200 lx、全光照培养条件下。分别设置加卤虫捕食组(卤虫3日龄,终密度1只/mL)以及不加卤虫的非捕食对照组。各3平行。分别在实验开始前和加入卤虫捕食24 h后收集各实验组藻液1000 mL,冰浴以停止其代谢过程,冷冻离心(5000 r/min,10 min,4 ℃)收集藻细胞制冻干藻粉备测。
总脂提取:分别称取各实验组藻粉50 mg,每组3个平行,参考改进后的Bligh-Dyer法[24],用CHCl3∶CH3OH∶H2O (1∶2∶0.8)溶剂提取3次,旋转蒸发仪真空干燥,氮气吹干,甲醇复溶,进行LC-MS分析。LC-MS条件以及数据处理分析参考文献[25]。
1.4 磷对颗石藻P.carterae细胞中DMSP含量的影响及其与卤虫致死率的关系提供不同的磷源以分析细胞DMSP含量差异,同时分析DMSP含量的差异是否造成P.carterae致害卤虫作用差异。
1.4.1 磷源差异对颗石藻P.carterae细胞内DMSP含量的影响将经过预培养至指数生长期的颗石藻P.carterae,接种到缺磷的f/2加富的人工海水[26]培养液中进行1周的磷饥饿培养。盐度28‰,pH 8.0±0.2。初始接种密度为4×105 个/mL。磷饥饿培养后,取藻液离心(6000 r/min,10 min,4 ℃)收集藻细胞制冻干藻粉备测初始参数。将处于磷饥饿状态的藻液分3组,其中一组以1∶1接种到缺磷的f/2培养液中,一组以1∶1接种到完整的以KH2PO4 为磷源的f/2培养液中,一组以1∶1接种到完整的以ATP为磷源(相同P摩尔数)的f/2培养液中。藻液总体积1000 mL,每组各3平行。培养条件为22℃,4000—5000 lx(D∶L=12 h∶12 h)。培养3d后,从每个实验组中取藻液800 mL,相同离心除上清液,收集藻细胞制冻干藻粉备测。
取等量冻干藻粉制样,于核磁共振仪(Bruker AVⅢ 400MHz谱仪,德国Bruker公司)检测样品DMSP含量。NMR定性定量检测及数据分析方法参考叶央芳(2011)[27]。DMSP的一维1HNMR谱图分别在化学位移2.74、2.76、3.44、3.48以及4.77处可见氢原子的吸收峰,通过颗石藻P.carterae提取物的一维1H NMR谱对DMSP定性定量分析。
1.4.2 磷源差异下培养的颗石藻P.carterae对卤虫致死率的影响分别取上述处于不同磷态的实验组藻液,置于24孔组织培养板,必要时,以对应实验组去藻滤液相应稀释,使各孔的藻细胞终密度为1×105 个/mL,藻液总体积2 mL。各孔分别加入10尾3日龄卤虫幼虫(以10 μL移液器每次吸取1尾),以相同密度的饵料金藻为喂养对照,以消毒海水为饥饿对照。每组3个平行。分别于0、24、48、72 h在倒置显微镜下检测卤虫死亡数。
2 结果 2.1 颗石藻P.carterae对卤虫摄食率的影响通过提取卤虫体内叶绿素a含量表征其捕食情况,实验结果如图 1所示,颗石藻P.carterae占比显著影响卤虫摄食:虽然缺乏实验前卤虫体内叶绿素状态的数据,但是,除I/P 8∶2组外,5 h叶绿素a含量差异不显著,均在0.35 mg/L以上。I/P 8∶2组最大,为0.46 mg/L。24 h,I/P 8∶2组叶绿素含量与5 h没有显著差异,可能也是因为该组初始状态体内食物含量即较高缘故。但其他各组叶绿素含量均比5 h降低,且随颗石藻P.carterae的占比增加而显著下降。此时,卤虫存活率还维持相对稳定的状态,如表 1的结果显示,5 h,实验组没有出现卤虫死亡,卤虫活跃。24 h,有卤虫死亡现象,但卤虫存活率与颗石藻P.carterae占比增加尚未见显著的梯度相关。因此,卤虫体内叶绿素含量的降低是卤虫摄食能力降低的体现。说明颗石藻对卤虫的捕食能力产生了影响。

|
| 图 1 球等鞭金藻(I)和颗石藻P.carterae(P)不同混合比例(I/P)混合喂养的卤虫体内叶绿素a含量 Fig.1 Chlorophyll a content in Artemia salina feeding on co-cultures of Isochrysis galbana (I)and Pleurochrysis carterea(P)mixed in different proportions (I/P) |
| 处理 Treatment | 存活率Survival rates/% | |||||
| I/P 10∶0 | I/P 8∶2 | I/P 5∶5 | I/P 2∶8 | I/P 0∶10 | 消毒海水Sterilized sea water | |
| 5 h | 100 | 100 | 100 | 100 | 100 | 100 |
| 24 h | 100 | 93.3 | 100 | 100 | 93.3 | 100 |
相同捕食压力下,颗石藻P.carterae和饵料金藻的叶绿素荧光参数呈现不同变化特征(图 2)。捕食压力作用24 h后,饵料金藻的最大量子产率Fv/Fm和电子传递速率ETR均显著下降(P < 0.01)。但颗石藻P.carterae的Fv/Fm以及ETR均没有显著差异(P > 0.05)。

|
| 图 2 捕食或无捕食压力下球等鞭金藻和颗石藻P. carterea的Fv/Fm和ETR变化 Fig.2 Changes of Fv/Fm and ETR in Isochrysis galbana and Pleurochrysis carterae when cultured with or without predators |
叶绿素荧光参数Fv/Fm和ETR反应了两种微藻受到捕食作用的差异,参数变化的特征显示,饵料金藻受到了来自卤虫的显著捕食胁迫,而卤虫对颗石藻P.carterae的捕食胁迫则显著小。因此,叶绿素参数特征也显示颗石藻P.carterae具有抗捕食能力。
2.2.2 捕食压力下,颗石藻P.carterae脂组代谢物变化经MarkerLynx处理,对微藻样品在正、负离子模式下信号峰的相对信号总强度经归一化进行PCA主成分分析。结果显示,饵料金藻在捕食压力存在与否的情况下,不同样品组的分布趋势没有差异,说明饵料金藻在捕食压力存在与否的情况下,其脂类组成没有变化。而颗石藻P.carterae的捕食压力样品组与没有捕食压力的对照组的分布趋势差异显著(图 3),说明颗石藻P.carterae在有捕食压力时,其脂类代谢发生显著变化(正负离子模式下R2分别为0.868和0.84)。

|
| 图 3 正负离子扫描模式,球等鞭金藻和颗石藻P.carterae在捕食压力存在与否情况下脂组PCA得分图 Fig.3 PCA scores plot in positive and negative ion scan mode for Isochrysis galbana and Pleurochrysis carterae with or without predators A: 球等鞭金藻的正离子模式,B: 球等鞭金藻的负离子模式,C: 颗石藻P.carterae正离子模式,D: 颗石藻P.carterae的负离子模式,空心点表示捕食压力组,实心点表示没有捕食压力的对照组 |
数据分析结果显示,相对于没有捕食压力对照组,捕食压力下,颗石藻P.carterae发生显著变化的脂类物质主要为:单半乳糖甘油二酯(MGDG)、双半乳糖甘油二酯(DGDG)、二酰甘油(DAG)、三酰甘油(TAG)、磷脂酰甘油(PG)、甜菜碱酯(DGCC,DGTS),二酰甘油硫代糖脂(SQDG)、磷脂酰胆碱(PC)以及磷脂酰肌醇(PI)。表 2显示了捕食压力下的颗石藻P.carterae细胞脂类相对于没有捕食压力的变化。结果显示,细胞的叶绿体脂类发生变化,MGDG,DGDG,PG都呈现上升,而SQDG下降;与促细胞分裂密切相关的DAG和PI也上升。因捕食压力造成的这些脂类含量差异,可能在颗石藻P.carterae抵抗捕食的生理代谢过程中有重要的意义。
| 脂类名称 Lipid species | PG | PI | PC | TAG | DAG | DGDG | MGDG | PA | DGCC | DGTS | SQDG |
| PG:磷脂酰甘油Phosphatidylglycerol,PI:磷脂酰肌醇Phosphatidylinositols,PC:磷脂酰胆碱Phosphatidylcholines,TAG:三酰甘油Triglycerides,DAG:二酰甘油Diglyceride,MGDG:单半乳糖基二酰甘油Monogalactosyldiacylglycerol,DGDG:双半乳糖基二酰甘油Digalactosyldiacylglycerol,PA:二酰甘油磷脂酸Phosphatidicacid,DGCC:二酰基甘油羧基羟甲基胆碱Diacylglyceryl-3-O-carboxyl-(hydroxymethyl)-choline,DGTS:二酰基甘油三甲基高丝氨酸Diacylglyceryl-N,N,N-trimethylhomoserine,SQDG: 硫代异鼠李糖甘油二脂Sulfoquinovosyldiacylglycerol;▲:相对于对照组升;▽:相对于对照组降 | |||||||||||
| 含量变化 Content variation | ▲ | ▲ | ▲ | ▲ | ▲ | ▲ | ▲ | ▽ | ▽ | ▽ | ▽ |
从表 3可以看出,相对于初始状态,缺磷条件下生长使细胞DMSP的含量增加。完整的f/2培养液中,细胞的DMSP含量下降。但增加或下降后的两组DMSP含量没有显著统计学差异。以ATP为唯一磷源时,细胞DMSP下降显著(P < 0.05)。结果显示,细胞的DMSP代谢与培养介质中磷态相关,能源物质ATP可能在DMSP代谢中发挥作用。
| 处理Treatment | f/2 (-P) | f/2 | f/2 (ATP) |
| DMSP:Dimethyl sulfonio propionate | |||
| 初始状态 Initial state | 2.35±0.27 | 2.35±0.37 | 2.35±0.27 |
| 培养3 d后After 3 days | 2.42±0.46 | 2.26±0.71 | 1.90±0.52 |
不同磷源差异培养液中的颗石藻P.carterae对卤虫的致死结果显示:各组藻液均对卤虫产生致死效应(图 4)。颗石藻P.carterae对卤虫无急性毒性,24 h内的致死效果不明显。但是,缺磷组DMSP含量相对高,在24 h首先显示卤虫致害。以ATP为磷源的实验组DMSP含量最低,其卤虫致死率在48 h和72 h的检查中始终为所有实验组中最低。由于缺磷组DMSP与完整f/2培养液中细胞DMSP含量差异不显著,可能导致这两组的卤虫致死率差异不显著(P > 0.05)。

|
| 图 4 磷差异状态造成颗石藻对卤虫的致死效应变化 Fig.4 Mortalities of Artemia salina in cultures of Pleurochrysis carterae cultured in mediums with different phosphorus states |
作为食物链中的初级生产者,高防御捕食能力的微藻具有更强的种群增殖能力。目前研究显示,微藻抗捕食的机制主要表现为形态学防御、化学防御和行为防御[28]。例如,硅藻的硅壳可能是长期进化过程中有利于抵抗捕食的一种结构[29]。Pondaven等的研究表明,威氏海链藻Thalassiosira weissflogii受到捕食压力时,细胞硅化程度会提高[30]。但是,通常认为,颗石藻P.carterae的钙壳并不具有抵抗捕食的作用,并且也不是颗石藻P.carterae致害卤虫的原因。Houdan等的也研究显示,Pleurochrysis属的几种颗石藻对卤虫的作用是不同的,颗石藻P.carterae具有显著的致死作用,而同属的P. dentata以及同样具有钙壳的Hymenomonadaceae科的几种颗石藻则没有致害作用[4]。
具有化学防御能力的微藻,通过向水中分泌捕食排斥物[31]或藻毒素[32, 33],使捕食者回避捕食或捕食后致害而实现抗捕食防御功能。例如,剧毒卡罗藻Karlodinium veneficum分泌的系列卡罗藻毒素(KmTx),可以使原生动物捕食者改变方向或致害捕食者[32]。Strom等研究发现,在颗石藻Emiliania huxleyi与隐藻Rhodomonas salina不同体积混合的藻液中,异养甲藻Amphidinium longum会优先摄食隐藻,从而使颗石藻E. huxleyi可以在竞争中胜出[34]。
DMSP是一种被广泛研究的水生生物化学防御物质。现代海洋中,生物量巨大且易形成赤潮的甲藻和定鞭藻都是DMSP高产类群。对不同级的捕食者而言,水生环境中的DMSP的作用有两种:诱导捕食和抵抗捕食。植食性原生动物可能回避DMSP及其裂解酶含量高的微藻,实现这类微藻的抗捕食化学防御目标[35]。DMSP通过机械或生化作用裂解产生挥发性DMS,又是诱导更高一级捕食者的信号物质[36]。
本文研究显示,随着颗石藻P.carterae在饵料金藻中的比例增加,卤虫的摄食率明显下降;有卤虫的捕食实验组中,颗石藻P.carterae的叶绿素荧光参数没有显著变化,证明其受到的捕食压力小,均说明颗石藻P.carterae有抵抗卤虫摄食能力。目前尚未在颗石藻中发现任何毒素,但普遍认为,细胞DMSP及其裂解产物DMS和丙烯酸在颗石藻与生物及非生物环境关系中发挥着重要作用[36]。同时,DMSP裂解酶及其活性在其间扮演重要角色。本研究结果显示,不同磷态培养液会使颗石藻P.carterae细胞内的DMSP含量发生变化,各培养条件下的颗石藻P.carterae细胞对卤虫的致害也有差异。缺磷组首先导致卤虫死亡;外源ATP培养条件下,对卤虫的致死率则降低。DMSP在藻细胞被消化时才产生DMS,同时产生的丙烯酸可能是致害原因[36]。实验镜检发现,卤虫肠道中有明显颗石藻P. carterae细胞。卤虫属滤食性,在饵料金藻与颗石藻P.carterae混合液中,可能无法选择或排斥颗石藻P.carterae细胞,从而导致受害。并且,不同DMSP裂解酶活性差异可能是产生卤虫致害的关键因素。有研究显示,氮限制对颗石藻E.huxleyi的细胞内DMSP含量没有影响[37, 38],但是,氮限制可提高DMSP裂解酶的活性,从而使氮限制条件下DMS的产量增加[38]。磷营养水平及化合物状态是否影响颗石藻P.carterae的DMSP裂解酶活性有待研究。DMSP裂解酶活性高的种类可能产生更高的致害产物。这样或许可以解释Houdan实验里不同种类的Pleurochrysis属的颗石藻不同的致害强度,以及在不同的生长周期,同种颗石藻对卤虫的致害差异。颗石藻是低营养环境中具有高竞争能力的微藻[39],在营养盐限制条件下,其DMSP裂解酶及其活性提高的特性,可能使其抵抗捕食能力提高,从而增加其种群竞争能力,实现增殖优势。
捕食胁迫下,浮游植物启动的抗捕食防御机制都是在复杂的代谢调节下实现的[28]。例如,硅藻除了以硅壳结构的物理性质抵抗捕食之外,还具有受捕食压力诱导的细胞硅化调节响应过程[30]。Zwirglmaier等[40]发现,聚球藻Synechococcus不同株细胞表面脂多糖的差异,形成了各株对捕食者的抗捕食能力差异。Parris等认为,微藻糖脂很可能是甲藻生态毒性的一个重要原因[20]。Cutignano等发现,海链藻Thalassiosira rotula的叶绿体膜上的糖脂是在浮游动物摄食过程中产生化学防御物质不饱和脂肪醛的来源[21]。本研究发现,颗石藻P. carterae脂类结构组显示其具有典型定鞭藻门(Haptophyta)的特点,即具有DGCC,这个类别的脂类在微藻中不常见,一般认为是进化上后期出现的脂类[41]。受到捕食压力和未受到捕食压力的颗石藻P. carterae比较,总脂组成有显著差异。其中细胞的叶绿体脂类MGDG,DGDG,PG都呈现上升,而只有SQDG下降,但一般认为MGDG和DGDG往往在叶绿体中更加重要[42, 43]。PI往往是细胞储能水平的一个前提物质,通过PI继续磷酸化过程,细胞内可以储存更多的能量,肌醇磷酸的能量储存比ATP更高[44]。DAG则反过来,往往是PI被裂解后产生的二次信使物质,往往是促进细胞分裂的信号[45]。颗石藻在受到捕食压力时,其脂组发生显著变化的这些特征,可能在其抵抗捕食以及致害捕食者的过程中发挥作用。颗石藻种群在经过捕食压力后,光合作用能力可能会上升,种群增殖能力可能会提高。但需要进一步的实验研究。
总之,本研究结果显示,利用卤虫与颗石藻P.carterae构成的捕食者与被捕食者体系中,颗石藻P. carterae使卤虫的摄食率显著降低、微藻叶绿素荧光参数无显著变化,均显示颗石藻P. carterae具有抵抗卤虫捕食特征。捕食压力下,颗石藻P.carterae的脂组代谢发生显著变化,可能在其抵抗捕食的生化调节过程中发挥重要作用。磷元素的生化代谢过程可能影响DMSP及其裂解酶的代谢调节,从而造成颗石藻P.carterae对卤虫致害的差异。具有高生物量广泛分布的颗石藻P.carterae,可能通过高效且多样的生理生化调节能力,达到抵御捕食压力的目标,实现种群增殖优势。
| [1] | Moore T S, Dowell M D, Franz B A. Detection of coccolithophore blooms in ocean color satellite imagery:A generalized approach for use with multiple sensors. Remote Sensing of Environment, 2012, 117:249-263. |
| [2] | 侯奎, 陈镇东, 陈延成. 颗石藻与海洋、环境关系之探讨. 化工矿产地质, 1999, 21(1):31-36, 56-56. |
| [3] | Li Y, Gao Y H, Huang D Q. Advances in study of marine Coccolithophorids. Marine Sciences, 2002, 26(3):13-16. |
| [4] | Houdan A, Bonnard A, Fresnel J, Fouchard S, Billard C, Probert I. Toxicity of coastal coccolithophores(Prymnesiophyceae, Haptophyta). Journal of Plankton Research, 2004, 26(8):875-883. |
| [5] | 蒋莹, 周成旭, 骆其君, 马斌. 颗石藻 Pleurochrysis carterae不同细胞状态对卤虫的致死效应研究. 生态毒理学报, 2009, 4(4):561-568. |
| [6] | 周成旭, 严小军, 孙雪, 徐继林, 傅永静. 一种颗石藻水华种的特征界定. 水生生物学报, 2008, 32(6):947-951. |
| [7] | Pohnert G, Steinke M, Tollrian R. Chemical cues, defence metabolites and the shaping of pelagic interspecific interactions. Trends in Ecology & Evolution, 2007, 22(4):198-204. |
| [8] | Tang K W. Grazing and colony size development in Phaeocystis globosa(Prymnesiophyceae):the role of a chemical signal. Journal of Plankton Research, 2003, 25(7):831-842. |
| [9] | Ianora A, Poulet S A, Miralto A, Grottoli R. The diatom Thalassiosira rotula affects reproductive success in the copepod Acartia clausi. Marine Biology, 1996, 125(2):279-286. |
| [10] | Teegarden G J, Campbell R G, Durbin E G. Zooplankton feeding behavior and particle selection in natural plankton assemblages containing toxic Alexandrium spp. Marine Ecology Progress Series, 2001, 218(1):213-226. |
| [11] | Ribalet F, Berges J A, Ianora A, Casotti R. Growth inhibition of cultured marine phytoplankton by toxic algal-derived polyunsaturated aldehydes. Aquatic Toxicology, 2007, 85(3):219-227. |
| [12] | 王岩, 张鸿雁. 海水实验围隔中桡足类对海洋原甲藻摄食的研究. 应用生态学报, 1990, 10(4):189-191. |
| [13] | Moheimani N R, Borowitzka M A. Limits to productivity of the alga Pleurochrysis carterae(Haptophyta) grown in outdoor raceway ponds. Biotechnology and Bioengineering, 2007, 96(1):27-36. |
| [14] | Wolfe G V, Levasseur M, Gantin G, Michaud S. DMSP and DMS dynamics and microzooplankton grazing in the Labrador Sea:application of the dilution technique. Deep Sea Research Part I:Oceanographic Research Papers, 2000, 47(12):2243-2264. |
| [15] | Steinke M, Codling E A, Breckels M N, Bode N W F. Effect of grazing-mediated dimethyl sulfide(DMS) Production on the swimming behavior of the copepod Calanus helgolandicus. Marine Drugs, 2013, 11(7):2486-2500. |
| [16] | 王秀娟, 周成旭, 严小军, 蒋莹. 颗石藻藻源性二甲基硫丙酸(DMSP)在致死卤虫时的作用检测. 生态毒理学报, 2010, 5(6):1-7. |
| [17] | 杨桂朋, 景伟文, 陆小兰. 海洋中DMSP的研究进展. 中国海洋大学学报, 2004, 34(5):854-860. |
| [18] | Sorgeloos P, Remiche-Van Der Wielen C, Persoone G. The use of Artemia nauplii for toxicity tests-a critical analysis. Ecotoxicology and Environmental Safety, 1978, 2(3/4):249-255. |
| [19] | 杨州, 孔繁翔, 史小丽, 张民, 曹焕生. 棕鞭毛虫牧食作用对铜绿微囊藻形态和生理特性的影响. 湖泊科学, 2008, 20(4):403-408. |
| [20] | Parrish C C, Bodennec G, Gentien P. Haemolytic glycoglycerolipids from Gymnodinium species. Phytochemistry, 1998, 47(5):783-787. |
| [21] | Cutignano A, d'Ippolito G, Romano G, Lamari N, Cimino G, Febbraio F, Nucci R, Fontana A. Chloroplastic glycolipids fuel aldehyde biosynthesis in the marine diatom Thalassiosira rotula. ChemBioChem, 2006, 7(3):450-456. |
| [22] | Satoh M, Iwamoto K, Suzuki I, Yoshihiro S. Cold Stress stimulates intracellular calcification by the Coccolithophore, Emiliania huxleyi(Haptophyceae) under phosphate-deficient conditions. Marine Biotechnology, 2009, 11(3):327-333. |
| [23] | Arnon D I. Copper enzymes in isolated chloroplasts, polyphenoloxidase in Beta vulgaris. Plant Physiology, 1949, 24(1):1-15. |
| [24] | Bligh E G, Dyer W J. A rapid method of total lipid extraction and purification. Canadian Journal of Biochemistry and Physiology, 1959, 37(8):911-917. |
| [25] | Su X L, Xu J L, Yan X J, Zhao P, Chen J J, Zhou C X, Zhao F, Li S. Lipidomic changes during different growth stages of Nitzschia closterium f. minutissima. Metabolomics, 2013, 9(2):300-310. |
| [26] | Harrison P J, Waters R, Taylor F J R. A broad spectrum artificial sea water medium for coastal and open ocean phytoplankton. Journal of Phycology, 1980, 16(1):28-35. |
| [27] | 叶央芳, 张利民, 安艳捧, 豪富华, 唐惠儒. 大肠杆菌代谢物组成的核磁共振分析. 分析化学研究报告, 2011, 39(8):1186-1194. |
| [28] | Roberts E C, Legrand C, Steinke M, Wootton E C. Mechanisms underlying chemical interactions between predatory planktonic protists and their prey. Journal of Plankton Research, 2011, 33(6):833-841. |
| [29] | Hamm C E, Merkel R, Springer O, Jurkojc P, Maier C, Prechtel K, Smetacek V. Architecture and material properties of diatom shells provide effective mechanical protection. Nature, 2003, 421(6925):841-843. |
| [30] | Pondaven P, Gallinari M, Chollet S, Bucciarelli E, Sarthou G, Schultes S, Jean F. Grazing-induced changes in cell wall silicification in a marine diatom. Protist, 2007, 158(1):21-28. |
| [31] | Robinette E D, Gulley K T, Cassity K J, King E E, Nielsen A J, Rozelle C L, Warren T J, Morrow J M, Kuruvilla H G. A comparison of the polycation receptors of Paramecium tetraurelia and Tetrahymena thermophila. Journal of Eukaryotic Microbiology, 2008, 55(2):86-90. |
| [32] | Waggett R J, Tester P A, Place A R. Anti-grazing properties of the toxic dinoflagellate Karlodinium veneficum during predator-prey interactions with the copepod Acartia tonsa. Marine Ecology Progress Series, 2008, 366:31-42. |
| [33] | Wolfe G V. The chemical defense ecology of marine unicellular plankton:constraints, mechanisms, and impacts. Biological Bulletin, 2000, 198(2):225-244. |
| [34] | Suzanne S, Wolfe G V, Holmes J, Stecher H, Shimeneck C, Lambert S, Moreno E. Chemical defense in the microplankton I:feeding and growth rates of heterotrophic protists on the DMS-producing phytoplankter Emiliania huxleyi. Limnology and Oceanography, 2003, 48(1):217-229. |
| [35] | Wolfe G V, Steinke M, Kirst G O. Grazing-activated chemical defence in a unicellular marine alga. Nature, 1997, 387(6636):894-897. |
| [36] | Wolfe G V, Steinke M. Grazing-activated production of dimethyl sulfide(DMS) by two clones of Emiliania huxleyi. Limnology and Oceanography, 1996, 41(6):1151-1160. |
| [37] | Keller M D, Kiene R P, Matrai P A, Bellows W K. Production of glycine betaine and dimethylsulfoniopropionate in marine phytoplankton. II. N-limited chemostat cultures. Marine Biology, 1999, 135(2):249-257. |
| [38] | Sunda W G, Hardison R, Kiene R P, Bucciarelli E, Harada H. The effect of nitrogen limitation on cellular DMSP and DMS release in marine phytoplankton:climate feedback implications. Aquatic Sciences, 2007, 69(3):341-351. |
| [39] | Dyhrman S T, Haley S T, Birkeland S R, Wurch L L, Cipriano M J, McArthur A G. Long serial analysis of gene expression for gene discovery and transcriptome profiling in the widespread marine Coccolithophore Emiliania huxleyi. Applied and Environmental Microbiology, 2006, 72(1):252-260. |
| [40] | Zwirglmaier K, Spence E, Zubkov M V, Scanlan D J, Mann N H. Differential grazing of two heterotrophic nanoflagellates on marine Synechococcus strains. Environmental Microbiology, 2009, 11(7):1767-1776. |
| [41] | Eichenberger W, Gribi C. Lipids of Pavlova lutheri:Cellular site and metabolic role of DGCC. Phytochemistry, 1997, 45(8):1561-1567. |
| [42] | Nakamura Y, Shimojima M, Ohta H, Kobayashi K. Biosynthesis and Function of Monogalactosyldiacylglycerol(MGDG), the Signature Lipid of Chloroplasts. In:Rebeiz C A, Benning C, Bohnert H J, Daniell H, Kenneth Hoober J, Lichtenthaler H K, Portis A R, Tripathy B C. The Chloroplast:Basics and Applications, Constantin. Advances in Photosynthesis and Respiration, 2010, 31:185-202. |
| [43] | Jouhet J, Maréchal E, Baldan B, Bligny R, Joyard J, Block M A. Phosphate deprivation induces transfer of DGDG galactolipid from chloroplast to mitochondria. The Journal of Cell Biology, 2004, 167(5):863-874. |
| [44] | Heydarizadeh P, Poirier I, Loizeau D, Ulmann L, Mimouni V, Schoefs B, Bertrand M. Plastids of marine phytoplankton produce bioactive pigments and lipids. Marine Drugs, 2013, 11(9):3425-3471. |
| [45] | Joyard J, Douce R. Site of synthesis of phosphatidic acid and diacyglycerol in spinach chloroplasts. Biochimicaet Biophysica Acta, 1997, 486(2):273-285. |