 2016, Vol. 36
2016, Vol. 36文章信息
- 王红妮, 王学春, 陶诗顺, 李军
- WANG Hongni, WANG Xuechun, TAO Shishun, LI Jun.
- 低温潜沼性逆境下不同耐受能力水稻根系动态变化
- The law of root changing of different rice varieties with different tolerance capacity under low temperature and gleization soil environment
- 生态学报[J]. 2016, 36(19): 6235-6245
- Acta Ecologica Sinica[J]. 2016, 36(19): 6235-6245
- http://dx.doi.org/10.5846/stxb201501130098
-
文章历史
- 收稿日期: 2015-01-13
- 网络出版日期: 2016-01-15





2. 西南科技大学生命科学与工程学院, 绵阳 621010;
3. Irrigated Agriculture Research & Extension Center, Washington State University, Prosser, Washington State, USA 99350
2. School of life science and technology, Southwest University of Science and Technology, Mianyang 621010, China;
3. Irrigated Agriculture Research & Extension Center, Washington State University, Prosser, Washington State, 99350 USA
低温潜沼性逆境是指分布在丘陵沟槽地带或山荫树林间,以“毒、闭、冷、烂、瘠”为主要特征,以土温和水温偏低(较正常稻田低2—5℃)、还原性物质和有机酸含量高、氧化还原电位低、矿质元素供应不足为主要障碍因子,直接或间接阻碍水稻正常生长发育,导致稻谷产量低而不稳的水稻生长环境[1-2]。四川盆地约有47万hm2 低温潜沼性稻田,占四川水稻总播种面积的22%—25%,由于春旱频发,当地农民常在水稻收获后开始蓄水以确保来年及时插秧。一般情况下,水稻拔节孕穗期要进行晒田,通过改善水稻生长环境,提高土壤养分有效性。然而,丘陵山区水利设施不足,伏旱严重,晒田后的灌溉用水无法保证,依靠工程蓄水和提灌来确保适时插秧和晒田,短期内难以实现,亟需探索新的途径与方法,克服低温潜沼性逆境,提高稻谷产量。
作物对生态逆境的适应能力可以在一定程度上缓解生态逆境对作物生长的不利影响,提高作物产量。李达模等[2]、李绍清等[3]和李阳生等[4-5]均认为,不同水稻品种对低温潜沼性逆境的耐受能力差异较大,依靠品种的耐性提高低温潜沼性稻田生产力,是比较经济且有效的途径[2-5]。王国莉等的研究结果表明,低温条件下,冷敏感品种的水稻产量比耐冷品种降低幅度更大[6]。赵锋等认为适当增加氧气供给,可以缓解淹水条件下缺氧对水稻的胁迫[7]。王学春等综合考虑穗粒数、有效分蘖等农艺指标,以水稻耐潜指数为主要依据对四川常用杂交水稻品种进行了分类[8]。李达模等[2]和李阳生等[3]认为水稻根系生长量与水稻耐低温潜沼性逆境能力具有很强的相关性。
为有效控制水分、养分等条件,本研究采用团队独立设计的水稻栽培环境模拟系统,进行盆栽试验,通过比较不同耐潜指数水稻品种在低温潜沼性逆境下的根系形态特征及不同土层分布情况,研究低温潜沼性逆境下水稻根系的动态变化规律,为低温潜沼性稻田水肥管理及耐潜品种筛选提供必要的理论依据。
1 材料与方法 1.1 盆栽环境1) 培养桶:装土,进行秸秆翻埋和水稻栽培试验; 2)控制桶:装水,控制培养桶水位,可连接多个培养桶试验于2012—2014年在西南科技大学青义试验基地展开。盆栽试验在水稻栽培环境模拟系统(图 1)中进行,试验地南北为丘陵山地,是典型的丘陵沟槽地带。上游(约100 m)为大型堰塘,通过水泵将堰塘底部冷水引入水稻栽培环境模拟系统,模拟低温潜沼性逆境的低温;将一定量的秸秆截短加入培养桶,翻埋入土,冬季淹水(3 cm)浸泡,提高培养桶土壤还原性物质总量,降低土壤氧化还原电位[9]。

|
| 图 1 水稻栽培环境模拟系统简图 Fig. 1 Schematic sketch of the rice cultivation simulating system |
连续两年测定结果表明,培养桶0—20cm土层地温较非低温潜沼性稻田低2.0—4.0℃;0—50 cm土层土壤氧化还原电位为-230—-400 mv,显著低于非低温潜沼性稻田,土壤还原性物质总量为45—60 cmol/kg,Fe2+含量为70—130 mg/kg,均显著高于非低温潜沼性稻田,符合低温潜沼性稻田特征。
1.2 试验设计试验在前期研究[8-9]基础上选取8个水稻品种作为实验材料(表 1),其中,协优027和C优130的耐潜指数超过12,耐潜等级为A;川香优907和Q优6号的耐潜指数大于6小于12,耐潜等级为B;Ⅱ优11和川丰2号的耐潜指数大于3小于6,耐潜等级为C;川谷优204和泸优5号的耐潜指数小于3,耐潜等级为D。
| 编号 Number | 品种 Variety | 耐潜指数 Index of gleization tolerance | 耐潜等级 Grade of gleization tolerance | 来源 Source |
| 1 | 协优027 | 16.50 | A | 协青早A×华恢027 |
| 2 | C优130 | 16.30 | CA×130 | |
| 3 | 川香优907 | 10.00 | B | 川香29A×华恢007 |
| 4 | Q优6号 | 8.40 | Q2A×R1005 | |
| 5 | II优11 | 6.00 | C | Ⅱ-32A×龙恢11 |
| 6 | 川丰2号 | 6.80 | 冈46A×江恢364 | |
| 7 | 川谷优204 | 3.00 | D | 川谷A×蜀恢204 |
| 8 | 泸优5号 | 1.04 | 泸077A×金恢5号 |
试验期间,每个水稻品种栽22桶,每桶单株栽插秧苗3株,培养桶半径0.35 m,栽培面积0.38 m2。其中11桶在低温潜沼性逆境下栽培(处理组);另外11桶在非低温潜沼性逆境下栽培(对照组)。处理组和对照组中各有5桶用来观测根系长度和数量,5桶用来观测根系分布特征,1桶用做定点观测分蘖动态,并测产(图 2)。对照组和处理组的灌溉、排水措施均与大田生产相同,即对照组冬季排水晒田,秧苗移栽后按正常管理排水晒田(除孕穗期和抽穗扬花期建立水层外,其余时期干湿交替、间歇灌溉),创造非低温潜沼性逆境;处理组冬季灌水泡田,水稻移栽后尽量减少排水,维持低温潜沼性逆境。

|
| 图 2 单个品种实验处理示意图 Fig. 2 Schematic sketch for the treatment of each rice variety |
水稻3月下旬开始育秧,秧龄45 d,5月中旬单株移栽(0.25 m×0.30 m),9月下旬收获。期间,施纯氮180 kg/hm2、五氧化二磷120 kg/hm2、氯化钾225 kg/hm2。肥料运筹方式为:氮肥以5∶3∶2的比例分别作基肥、分蘖肥、穗肥施入,磷肥在秧苗移栽前以基肥施入,钾肥以1∶1的比例分别作基肥和穗肥施入。其它管理与当地大田相同。
1.3 取样与统计水稻返青-分蘖盛期,每15 d取6株(A1、C1和A2、C2各1桶)。其中,A1、A2各取1桶将栽培套筒取下,用水流冲洗水稻根部,观测根系长度和数量;C1、C2各取1桶将栽培套筒取下,用电锯将培养柱自上而下,分三段(0—10 cm,10—30 cm,30—50 cm)切割,分段观测根系分布特征。统计根系数量时,每个分枝计数一次,根长小于2 cm不计入统计数;根系长度为所有根系的总长度;白根只统计整条或一半以上颜色为白色的根。
移栽后至7月底,每个水稻品种选取6株(B1和B2各1桶)定点观测株高、分蘖数等;9月下旬,将稻穗带回实验室风干考种。考种指标包括:千粒重、穗长、实粒数、着粒数等。
水稻根系特征受品种特性影响较大,品种间的根系长度和数量等直接进行比较,难以分析出不同水稻品种在低温潜沼性逆境下的根系特征差异及逆境对水稻根系生长的影响。本研究引入绝对变化率(M)和相对变化率(P),分别反映不同耐潜指数水稻品种在低温潜沼性逆境下的自身生长规律以及低温潜沼性逆境对不同品种生长规律的影响程度。绝对变化率(M)指水稻根系某一考察指标比上次测量增加的百分比(公式1),M值越大,根系生长越快;相对变化率(P)指以非低温潜沼性逆境为参照,根系某一考察指标比上次测量结果增加的百分比(公式2),P值越大,低温潜沼性逆境对水稻根系的影响越小:
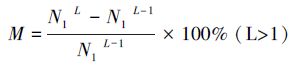 (1)
(1)
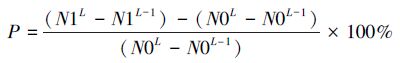 (2)
(2)
式中,M为根系某一指标的绝对变化率,P为根系某一指标的相对变化率。N1L和N1L-1分别为第L次和第L-1次观测时,水稻根系某指标在低温潜沼性逆境下的观测值; N0L和N0L-1分别为第L次和第L-1次观测时,水稻根系某指标在非低温潜沼性逆境中的观测值。
采用SAS8.0对数据进行差异显著性和相关性分析,采用3年数据平均值对相关指标进行比较和分析,以降低气象因素对数据分析和试验结果的影响。
2 结果与分析 2.1 低温潜沼性逆境下水稻根系变化规律水稻移栽后至7月底(表 2),8个品种的根系长度M值均大于0,而根系长度P值均小于0,表明低温潜沼性逆境下水稻根系长度虽有一定量的增加,但与非低温潜沼性稻田相比,根系生长较为缓慢;1、2号品种的根系长度M值与P值显著高于7、8号,表明耐潜指数高的品种在低温潜沼性逆境下根系长度的平均生长量显著高于耐潜指数低的品种,且根系长度的降低程度显著低于耐潜指数低的品种;8个品种的根系长度M值在6月30日均达到最大,随后开始减少,根系长度P值在6月15日均降到最小,随后开始升高,表明随着时间的推移,低温潜沼性逆境下水稻根系生长速率表现为先升高后降低的趋势,且逆境对根系生长的影响程度逐渐降低。
| 品种 Variety | 5月31日May 31 | 6月15日June 15 | 6月30日June 30 | 7月15日July 15 | 7月30日July 30 | |||||
| M/% | P/% | M/% | P/% | M/% | P/% | M/% | P/% | M/% | P/% | |
| 1 | 91.80b | -22.92d | 154.80a | -36.59a | 201.25a | -29.98a | 66.33a | -12.65a | 22.6a | 0.00a |
| 2 | 97.26a | -24.18c | 152.49a | -39.10a | 198.24a | -32.44ab | 67.57a | -14.98a | 21.5ab | 0.00a |
| 3 | 74.05d | -25.45bc | 135.38b | -41.60ab | 176.00b | -34.89b | 62.60b | -17.32b | 19.4bc | -0.09a |
| 4 | 81.22c | -24.99c | 132.20b | -40.69ab | 171.86b | -34.00b | 61.36b | -16.47b | 19.7b | -0.06a |
| 5 | 74.05d | -26.71b | 125.53c | -44.11b | 159.42c | -37.34c | 58.87c | -19.65c | 18.5c | -1.03b |
| 6 | 72.00e | -27.06b | 122.63c | -44.80b | 163.19c | -38.01c | 61.25b | -20.28c | 16.7d | -2.09b |
| 7 | 68.59g | -35.57a | 91.90d | -61.67c | 119.47d | -54.51d | 59.55c | -35.97d | 15.8e | -2.18b |
| 8 | 69.28f | -36.95a | 94.51d | -64.40c | 122.86d | -57.19d | 58.87c | -38.52d | 13.6f | -4.19b |
M为绝对变化率absolute changing rate,P 为相对变化率relative changing rate,后表及后图的M和P含义与此相同; 数字后的小写字母表示品种间的差异显著性
水稻移栽后至7月底(表 3),8个品种的根系数量M值均大于0,而P值均小于0,表明低温潜沼性逆境下水稻根系数量仍有一定量的增加,但与非低温潜沼性稻田相比,新根生成速率较低;1、2号品种的根系数量M值和P值显著高于7、8号,表明耐潜指数高的品种在低温潜沼性逆境下的新根平均生成量显著高于耐潜指数低的品种,且根系数量的降低程度显著低于耐潜指数低的品种。8个品种的根系数量M值在6月15日均达到最大,随后又降低,而根系数量P值基本保持逐渐增加的趋势,表明随着时间的推移,低温潜沼性逆境下水稻新根生成速率表现为先升高后降低的趋势,且逆境对水稻新根生成的影响程度逐渐降低。
| 品种 Variety | 5月31日May 31 | 6月15日June 15 | 6月30日June 30 | 7月15日July 15 | 7月30日July 30 | |||||
| M/% | P/% | M/% | P/% | M/% | P/% | M/% | P/% | M/% | P/% | |
| 1 | 81.24a | -10.32a | 119.08a | -10.14a | 86.18a | -7.07a | 55.18a | 0.00a | 22.91a | 3.20a |
| 2 | 86.07a | -10.58a | 117.30a | -10.15a | 88.62a | -7.07a | 56.21a | 0.00a | 19.24b | 2.10b |
| 3 | 65.53c | -12.69b | 104.14b | -11.31b | 79.19b | -7.65b | 52.08b | -0.08a | 17.36c | 0.00c |
| 4 | 71.88b | -12.65b | 101.69b | -12.29c | 79.34b | -8.14b | 51.04b | -0.02a | 17.92c | 0.00c |
| 5 | 65.53c | -11.80c | 96.56c | -12.35c | 76.15c | -8.17b | 50.27c | -0.80b | 15.61d | -0.02d |
| 6 | 63.72c | -11.83c | 94.33c | -12.37c | 76.30c | -8.18b | 50.95c | -0.60b | 14.95d | -0.05d |
| 7 | 60.70d | -13.57d | 70.69d | -13.74d | 72.35d | -8.87c | 49.54d | -1.10c | 11.33e | -0.13e |
| 8 | 61.31d | -15.69d | 72.70d | -15.74d | 65.66e | -9.87c | 48.97cd | -1.20c | 10.11e | -0.14e |
水稻移栽后至7月底(表 4),8个品种的白根长度M值在6月30日前均为正值,随后成负值,P值在6月15日前相对较小,表明低温潜沼性逆境下水稻根系在6月30日前具有较强的活力,随后根系活力开始下降,且移栽后至分蘖始期,逆境对新根生长影响较大;1、2号品种白根长度M值和P值的5次观测平均值均显著高于7、8号,表明在低温潜沼性逆境下,耐潜指数高的水稻品种根系活力较强,且新根生成受逆境影响的程度相对较小;8个品种的白根长度M值均在6月30日达到最大,随后开始降低,而白根长度P值基本保持逐渐增加趋势,表明随着时间的推移,低温潜沼性逆境下水稻白根长度表现为先升高后降低的趋势,且逆境对白根长度的影响程度逐渐降低。
| 品种 Variety | 5月31日May 31 | 6月15日June 15 | 6月30日June 30 | 7月15日July 15 | 7月30日July 30 | |||||
| M/% | P/% | M/% | P/% | M/% | P/% | M/% | P/% | M/% | P/% | |
| 1 | 52.62a | -7.11a | 264.14a | -7.01a | 307.70a | -3.24a | -49.94a | -0.73a | -17.11a | 0.00a |
| 2 | 52.34a | -7.16a | 265.10a | -7.02a | 305.85a | -5.23b | -50.16a | -0.98a | -17.18a | 0.00a |
| 3 | 46.32b | -8.52b | 234.64b | -8.48b | 264.74b | -8.35c | -70.29b | -7.84b | -23.22b | -2.63b |
| 4 | 45.92b | -8.50b | 233.68b | -8.46b | 264.74b | -8.25c | -71.06b | -7.81b | -23.45b | -3.14b |
| 5 | 37.87c | -10.07c | 182.34c | -10.02c | 211.37c | -9.9d | -86.35c | -9.7c | -28.04c | -9.30c |
| 6 | 37.99c | -10.09c | 183.63c | -10.04c | 211.37 c | -9.85d | -86.79c | -9.6c | -28.17c | -9.30c |
| 7 | 27.46d | -12.36d | 137.97d | -12.31d | 143.48 d | -12.2e | -99.11d | -11.75d | -31.86d | -10.24d |
| 8 | 27.40d | -12.42d | 137.02d | -12.31d | 149.05 d | -12.2e | -99.44d | -11.8d | -31.96d | -10.25d |
水稻移栽后至7月底(表 5),8个品种的白根数量M值在6月30前为正值,随后成为负值,P值在6月15日前相对较小,表明低温潜沼性逆境下水稻根系在6月30日前具有较强的新根生成能力,随后开始下降,且移栽后至分蘖始期,逆境对水稻新根生长的影响较大;1、2号品种白根数量M值和P值的5次观测平均值均显著高于7、8号,表明在低温潜沼性逆境下,耐潜指数高的水稻品种新根生成能力较强,且新根生成受逆境影响的程度相对较小;8个品种的白根数量M值均在6月30日达到最大,随后开始减少,而白根数量P值基本保持逐渐增加趋势,表明随着时间的推移,低温潜沼性逆境下水稻白根数量表现为先升高后降低的趋势,且逆境对白根数量的影响程度逐渐降低。
| 品种 Variety | May 31 | June 15 | June 30 | July 15 | July 30 | |||||
| M/% | P/% | M/% | P/% | M/% | P/% | M/% | P/% | M/% | P/% | |
| 1 | 39.88a | -12.33a | 133.05a | -12.29a | 139.57a | -11.52a | -45.01a | -9.75a | -28.93a | -8.95a |
| 2 | 40.10a | -12.40a | 132.85a | -12.29a | 140.00a | -11.52a | -45.06a | -9.75a | -29.00a | -9.23a |
| 3 | 33.17b | -14.92b | 110.45b | -14.58b | 117.36b | -13.66b | -51.71b | -11.77b | -33.21b | -11.75b |
| 4 | 33.33b | -14.91b | 110.66b | -14.82b | 117.36b | -13.79b | -51.91b | -11.76b | -33.27b | -11.75b |
| 5 | 29.99c | -15.70c | 101.09c | -15.84c | 106.30c | -14.79c | -57.11c | -12.95c | -36.64c | -12.76c |
| 6 | 29.96c | -16.21c | 100.88c | -15.84c | 106.30c | -14.80c | -57.22c | -12.90c | -36.71c | -12.76c |
| 7 | 25.76d | -17.44d | 84.80d | -17.49d | 91.95d | -16.27d | -67.83d | -14.33d | -43.22d | -14.08d |
| 8 | 25.79d | -17.97d | 85.00d | -17.99d | 90.99d | -16.52d | -67.98d | -14.35d | -43.32d | -14.09d |
低温潜沼性逆境下,不同土层水稻根系长度M值变化规律不同(图 3)。其中,0—10 cm土层M值在6月30日前逐渐增加,之后逐渐降低;10—30 cm土层M值先缓慢增加,后缓慢降低;30—50 cm土层M值在7月15日前逐渐增大,之后逐渐降低。

|
| 图 3 低温潜沼性逆境下不同耐潜等级水稻品种在不同土层根系长度动态变化比较 Fig. 3 Comparison of the rood length of different rice varieties in different soil layers under the low temperature and gleization soil environment |
不同耐潜等级水稻品种间根系长度M值差异因土层的不同而不同(图 3)。其中,0—10 cm土层M值差异在6月30日前逐渐扩大,之后逐渐缩小;10—30 cm土层M值差异变化不大;30—50 cm土层M值差异随着时间的推移逐渐扩大。
低温潜沼性逆境下,不同耐潜等级水稻品种的根系长度P值在不同土层变化规律不同(图 3)。其中,0—10 cm土层P值在6月15日前后最小,A、B、C、D 4个耐潜等级的P值分别为-42.1%、-53.7%、-62.8%和-66.4%;10—30 cm土层P值也在6月15日前后最小,A、B、C、D 4个耐潜等级的P值分别为-32.8%、-43.5%、-47.4%和-54.8%;30—50 cm土层P值在6月30日前后最小,A、B、C、D 4个耐潜等级的P值分别为-35.6%、-45.2%、-53.2%和-63.8%。
不同耐潜等级水稻品种间根系长度P值差异因土层的不同而不同(图 3)。其中,0—10 cm土层P值差异在6月15日前最大,之后逐渐缩小;10—30 cm土层P值差异在6月15日前后最大,5月下旬和7月底最小;30—50 cm土层P值差异在6月30日前后最大,5月底至6月初最小。
低温潜沼性逆境下,不同土层水稻根系数量M值均表现为先增加后降低的趋势(图 4)。其中,0—10 cm土层M值在6月15日至6月30日最大;10—30 cm土层M值在6月30日前后最大;30—50 cm土层M值在7月15日前逐渐增大,之后逐渐降低。

|
| 图 4 低温潜沼性逆境下不同耐潜等级水稻品种在不同土层根系数量动态变化比较 Fig. 4 Comparison of the Rood number of different rice variety in different soil layers under the low temperature and gleization soil environment |
不同耐潜等级水稻品种间根系数量M值差异因土层的不同而不同(图 4)。其中,0—10 cm土层M值差异在6月30日前逐渐扩大,之后逐渐缩小;10—30 cm土层M值差异在6月30日前逐渐扩大,之后逐渐缩小;30—50 cm土层M值差异随时间的推移逐渐扩大。
低温潜沼性逆境下,不同耐潜等级水稻品种的根系数量P值在不同土层变化规律不同(图 4)。其中,0—10 cm土层P值在5月31日前后最小,A、B、C、D 4个耐潜等级的P值分别为-48.48%、-64.35%、-75.50%和-89.37%;10—30 cm土层P值在6月15日前后最小,A、B、C、D 4个耐潜等级的P值分别为-34.13%、-44.94%、-48.87%和-56.35%;30—50 cm土层P值在6月30日前后最小,A、B、C、D 4个耐潜等级的P值分别为-41.23%、-52.08%、-61.12%和-73.09%。
不同耐潜等级水稻品种间根系数量P值差异因土层的不同而不同(图 4)。其中,0—10 cm土层P值差异在5月31日最大,之后逐渐缩小;10—30 cm土层P值差异在6月15日前后最大,其次为5月下旬,7月底最小;30—50 cm土层P值差异在6月30日前后最大,5月底至6月初最小。
2.4 低温潜沼性逆境下水稻产量构成因素低温潜沼性逆境下,不同耐潜指数的水稻品种千粒重间存在一定差异,但与非低温潜沼性逆境相比,其变化不显著(表 6)。其中,8个参试品种中,只有3号和7号品种的千粒重与非低温潜沼性逆境下的千粒重存在显著差异,其它品种间的差异均不显著,表明低温潜沼性逆境下,品种间的千粒重差异主要由水稻品种本身特性引起。
| 品种 Variety | 有效穗数 Available panicle | 千粒重/g Weight of 1000 grains | 实粒数/粒 Full grains per panicle | 结实率/% Rate of seed setting | 产量/(t/hm2) Yield | ||||||||||
| LTGS | CK | P>t | LTGS | CK | P>t | LTGS | CK | P>t | LTGS | CK | P>t | LTGS | CK | P>t | |
| 1 | 14.3 | 15.3 | 0.012 | 27.1 | 27.2 | 0.163 | 101.8 | 145.8 | 0.021 | 60.2 | 77.5 | 0.011 | 6.2 | 9.6 | 0.009 |
| 2 | 13.8 | 15.2 | 0.004 | 29.8 | 30.2 | 0.181 | 100.0 | 148.0 | 0.018 | 57.3 | 72.4 | 0.029 | 6.5 | 10.7 | 0.001 |
| 3 | 11.7 | 13.5 | 0.003 | 27.1 | 29.1 | 0.040 | 98.0 | 136.0 | 0.009 | 65.8 | 75.6 | 0.013 | 4.9 | 8.4 | 0.002 |
| 4 | 11.5 | 13.8 | 0.003 | 28.0 | 28.0 | 0.250 | 92.3 | 132.3 | 0.007 | 70.5 | 75.8 | 0.026 | 4.7 | 8.1 | 0.002 |
| 5 | 10.0 | 12.0 | 0.004 | 28.3 | 28.5 | 0.101 | 87.9 | 120.9 | <0.001 | 61.5 | 76.5 | 0.021 | 3.9 | 6.5 | 0.006 |
| 6 | 9.2 | 12.2 | 0.002 | 26.4 | 26.3 | 0.215 | 85.2 | 121.2 | <0.001 | 62.8 | 70.9 | 0.017 | 3.3 | 6.1 | 0.004 |
| 7 | 8.5 | 11.5 | 0.001 | 27.1 | 27.3 | 0.042 | 76.0 | 112.0 | <0.001 | 53.1 | 70.2 | 0.009 | 2.8 | 5.6 | 0.002 |
| 8 | 8.5 | 11.5 | 0.001 | 28.1 | 30.2 | 0.217 | 62.9 | 113.9 | <0.001 | 58.9 | 78.9 | 0.007 | 2.4 | 6.3 | 0.001 |
| LGTS:低温潜沼性逆境下栽培 rice was planted at the low temperature and gleization soil environment;CK:非低温潜沼性逆境下栽培 no LGTS stress on rice | |||||||||||||||
与非低温潜沼性逆境下的结实率相比,低温潜沼性逆境下,所有参试品种的实粒数和结实率均有不同程度的降低,相对而言,耐潜指数越高,结实率越高。表明,低温潜沼性逆境对水稻结实率的影响较大(表 6)。
2.5 低温潜沼性逆境下水稻根系特征与产量构成间相关性分析低温潜沼性逆境下,不同时期水稻根系特征与产量构成因素相关性不同。水稻返青至分蘖初期(6月15日前)根系主要形态特性与产量构成因素间有一定相关性,但有相关性的根系特征不多。其中,根系长度和数量均与结实率呈显著正相关,相关系数分别为0.64和0.68;白根长度和数量均与着粒数呈显著正相关,相关系数分别为0.65和0.62。
孕穗-抽穗期(6月中下旬至7月下旬)水稻根系主要形态特性与产量构成因素呈显著相关性(表 7)。其中根系长度和数量与着粒数、实粒数、结实率、产量之间呈显著正相关,与单株有效穗数和千粒重呈显著负相关;白根长度和数量与实粒数和结实率呈显著负相关,与着粒数的相关性在孕穗期未达到显著水平,在抽穗期则呈俩俩显著负相关,与单株有效穗数的相关性在孕穗期也未达到显著水平,在抽穗期则呈俩俩显著正相关,与千粒重呈俩俩显著正相关。
| 农艺性状 Agronomic traits | 根系长度 Total length of root | 根系数量 Total number of root | 白根长度 Length of white root | 白根数量 Number of white root |
| 有效穗数Available panicle | -0.873* | -0.782* | 0.785* | 0.722* |
| 着粒数Grains per panicle | 0.698* | 0.658* | -0.842* | -0.898* |
| 实粒数Full grains per panicle | 0.885* | 0.849* | -0.951* | -0.954* |
| 结实率Seed setting rate | 0.903* | 0.932* | -0.982* | -0.752* |
| 千粒重Weight of 1000 grains | -0.642* | -0.618* | 0.740* | 0.888* |
| 产量 Yield | 0.523* | 0.157* | 0.342* | -0.014 |
低温潜沼性逆境下,不同土层水稻根系形态特征与产量构成因素间的相关性不同。其中,0—10 cm土层根系长度和数量与有效分蘖数呈显著负相关,相关系数为-0.684和-0.742;10—30 cm 土层白根长度和数量与结实率呈显著正相关,相关系数分别为0.852和0.637;30—50 cm土层根系长度和数量与着粒数呈显著正相关,相关系数分别为0.826和0.761。
3 讨论与结论 3.1 讨论氧气对根尖细胞的伸长和延续是必须的,适当晒田给根系提供了一定的氧气,间歇灌溉条件下水稻根系的生物量显著高于淹水灌溉条件[10];在低温潜沼性逆境下,水稻根区氧气供应相对不足,根系生长受到抑制,水稻根系伸长速率及生成速率都显著低于非低温潜沼性稻田。在低温潜沼性逆境下,耐潜指数高的水稻品种的根系长度M值和P值与根系数量M值和P值显著高于耐潜指数低的品种(表 2,表 3)。李阳生等的研究结果表明在淹水条件下,耐淹能力强的水稻品种,白根数量显著高于耐淹能力弱的品种[11]。与非低温潜沼性稻田相比,在低温潜沼性逆境下,白根长度P值和白根数量P值长期小于0,白根生长速率和生成速率显著降低。白根数量的减少,表明水稻根系在低温潜沼性逆境下活力较低。耐潜指数高的水稻品种的白根长度M值和P值与白根数量M值和P值显著高于耐潜指数低的品种(表 4,表 5)。耐潜指数高的水稻品种白根生成速率和生长速率显著高于耐潜指数低的品种。在苗期水稻品种筛选过程中,可考虑将水稻根系白根数量和长度作为耐潜能力强弱的指标之一。
水稻根系由不定根和定根构成,不同类型的水稻根系在其发生位置、数量、直径、功能以及分布特征上均存在差异,这些差异是根系对土壤环境适应的重要表现[12-13]。侧根和根毛数量的增多,可以看作水稻对不良氧营养状况在形态学上的自身调整[7],淹水条件下水稻根系主要分布于上层土壤,适度控水能促进根系生长,尤其是不定根的发生,并促进根系向中下层土壤扩展,但重度干旱会影响根系生长[14]。低温潜沼性逆境下,0—10 cm土层根系在整个6月生长较快(根系长度和根系数量的M值均达到最大);10—30 cm土层根系在6月下旬至7月上旬生长较快(根系长度M值在7月15日达到最大、根系数量M值在6月30日达到最大);30—50 cm土层根系在7月15日前后生长最快,之后生长速率开始降低(图 3,图 4)。
李达模等的研究结果表明,在低温潜沼性稻田,随着水稻生育进程的发展,根系生长量的受害程度逐渐加重[2]。在低温潜沼性逆境下,不同土层土壤温度、氧气含量、还原性物质总量和氧化还原电位等差异较大[15]。虽然水稻根系对逆境具有一定的适应能力,但根系的生长对根区氧气含量和根区温度敏感[16],低温环境不利于根系生长,低于22℃次生根不能生长[17]。因此不同土层对土壤根系生长的影响不同。本研究表明,低温潜沼性逆境对水稻根系生长的影响因土层的不同而不同(图 3,图 4)。低温潜沼性逆境下,0—10 cm土层根系生长在5月底6月初受到的影响最大;10—30 cm土层根系生长在6月底受到的影响最大; 30—50 cm土层根系生长在6月30日前后受到的影响最大。根系形态及生理特性与地上部生长发育及产量形成有密切联系,良好的根系构型是维持作物产量的基本保证[18-19]。本研究表明,低温潜沼性逆境下,孕穗至抽穗期的水稻根系长度和数量与水稻着粒数、实粒数、结实率呈显著正相关,而白根长度和数量则与水稻着粒数、实粒数、结实率呈显著负相关(表 7),这与赵启辉[1]和Eriko等[19]的研究结果相似。
李阳生等对29个杂交稻及其亲本苗期耐淹涝特性的研究证实,杂交稻的耐淹涝能力具有显著的杂交优势,恢复系和不育系耐性强,配出的杂交稻耐性更强[4]。近年,国内外对水稻耐逆境的育种取得了较大进步,Sangin等通过对多个水稻亲本的比较研究表明,M202,IR50 可作为选育耐低温水稻品种的亲本材料[20];Surekha 等对水稻耐Fe2+特性进行了基因定位研究,认为对水稻根系耐Fe2+有重要作用的基因为IRT1,IRT2,YSL2 和 ZIP8[21];Eduardo 等在对多个水稻品种筛选的基础上,研究了Fe2+对水稻光合作用的影响,认为BRA 041171 和 BRA 041152 在高Fe2+浓度下的光合作用相对较好[22],这些研究为选育耐低温潜沼性逆境水稻品种奠定了良好的前期基础,但低温潜沼性逆境是多种逆境(淹水期较长、温度偏低、还原性物质含量高、氧化还原电位低等)的综合体,仅考虑一种因素进行品种选育难以适应。本研究通过水稻栽培环境模拟系统研究了低温潜沼性逆境下不同耐潜指数水稻品种的根系动态变化规律,对今后耐潜品种的选育具有一定的指导意义。
3.2 结论(1) 低温潜沼性逆境下,水稻根系生长缓慢,长度和数量均显著降低,降低程度因品种耐潜指数的高低而不同;耐潜指数高的品种根系长度和数量降低程度显著低于耐潜指数低的品种。在水稻耐潜品种选育过程中,根系长度和数量的M值和P值,尤其是白根长度和数量的M值和P值可作为一个耐潜性评价指标进行考虑。
(2) 低温潜沼性逆境对根系生长的影响,因土层和水稻生育期的不同而不同。低温潜沼性逆境对0—10 cm土层根系的影响在5月底6月初最大;对10—30 cm土层根系的影响在6月底最大;对30—50 cm土层根系的影响在6月30日前后最大。
(3)低温潜沼性逆境下,孕穗-抽穗期水稻根系特征与产量间存在显著相关性;不同土层水稻根系与产量构成因素间的相关性不同。0—10 cm 土层根系长度和数量与有效分蘖数呈显著负相关;10—30 cm 土层白根长度和数量与结实率呈显著正相关;30—50 cm土层根系长度和数量与着粒数呈显著正相关。
| [1] | 赵启辉. 分蘖期淹涝胁迫对水稻农艺和品质性状及生理特性的影响[D]. 南昌:江西农业大学, 2013. |
| [2] | 李达模, 唐建军, 李阳生. 水稻耐(抗)潜育性土壤逆境的生理生态机制及抗逆品种选育进展. 水稻文摘 , 1991, 10 (2) : 1–4. |
| [3] | 李绍清, 李阳生, 李达模, 廖江林. 乳熟期淹水对两系杂交水稻源库特性的影响. 杂交水稻 , 2000, 15 (2) : 38–40. |
| [4] | 李阳生, 李绍清, 李达模, 李振声. 杂交稻与常规稻对涝渍环境适应能力的比较研究. 中国水稻科学 , 2002, 16 (1) : 45–51. |
| [5] | 李阳生, 李达模, 周建林, 李绍清. 水稻耐不良土壤因子胁迫品种改良的现状和展望. 中国水稻科学 , 2000, 14 (2) : 107–111. |
| [6] | 王国莉, 郭振飞. 低温下水稻不同品种的光合作用及磷的影响//中国植物生理与分子生物学学会. 中国植物生理学会第九次全国会议论文摘要汇编. 上海:中国植物生理与分子生物学学会 , 2004 : 99. |
| [7] | 赵锋, 王丹英, 徐春梅, 张卫建, 章秀福. 水稻氧营养的生理、生态机制及环境效应研究进展. 中国水稻科学 , 2009, 23 (4) : 335–341. |
| [8] | 王学春, 王红妮, 杨国涛. 不同基因型水稻对低温潜沼性逆境的耐受能力及其农艺表现. 云南大学学报:自然科学版 , 2014, 36 (5) : 765–773. |
| [9] | 王红妮, 王学春, 陶诗顺, 李军. 秸秆残茬对低温潜沼性稻田土壤还原性物质含量及稻谷产量的影响. 干旱地区农业研究 , 2014, 32 (3) : 179–183. |
| [10] | 徐芬芬, 曾晓春, 石庆华, 叶利民. 不同灌溉方式对水稻根系生长的影响. 干旱地区农业研究 , 2007, 25 (1) : 102–104. |
| [11] | 李阳生, 彭凤英, 李达模, 李振声. 杂交水稻苗期耐淹涝特性及其与亲本的关系. 杂交水稻 , 2001, 16 (2) : 50–53. |
| [12] | Khan M R, Haque Z, Kausar N. Management of the root-knot nematode Meloidogyne graminicola infesting rice in the nursery and crop field by integrating seed priming and soil application treatments of pesticides. Crop Protection , 2014, 63 : 15–25. DOI:10.1016/j.cropro.2014.04.024 |
| [13] | Kijoji A A, Nchimbi-Msolla S, Kanyeka Z L, Serraj R, Henry A. Linking root traits and grain yield for rainfed rice in sub-Saharan Africa:Response of Oryza sativa×Oryza glaberrima introgression lines under drought. Field Crops Research , 2014, 165 : 25–35. DOI:10.1016/j.fcr.2014.03.019 |
| [14] | Gowda V R P, Henry A, Yamauchi A, Shashidhar H E, Serraj R. Root biology and genetic improvement for drought avoidance in rice. Field Crops Research , 2011, 122 (1) : 1–13. DOI:10.1016/j.fcr.2011.03.001 |
| [15] | Henry A, Gowda V R P, Torres R O, Mcnally K L, Serraj R. Variation in root system architecture and drought response in rice (Oryza sativa):Phenotyping of the OryzaSNP panel in rainfed lowland fields. Field Crops Research , 2011, 120 (2) : 205–214. DOI:10.1016/j.fcr.2010.10.003 |
| [16] | Passaia G, Fonini L S, Caverzan A, Jardim-Messeder D, Christoff A P, Gaeta M L, De Araujo Mariath J E, Margis R, Margis-Pinheiro M. The mitochondrial glutathione peroxidase GPX3 is essential for H2O2 homeostasis and root and shoot development in rice. Plant Science , 2013, 208 : 93–101. DOI:10.1016/j.plantsci.2013.03.017 |
| [17] | Xu C M, Wang D Y, Chen S, Chen L P, Zhang X F. Effects of aeration on root physiology and nitrogen metabolism in rice. Rice Science , 2013, 20 (2) : 148–153. DOI:10.1016/S1672-6308(13)60118-3 |
| [18] | Singh I, Shah K. Exogenous application of methyl jasmonate lowers the effect of cadmium-induced oxidative injury in rice seedlings. Phytochemistry , 2014, 108 : 57–66. DOI:10.1016/j.phytochem.2014.09.007 |
| [19] | Maejima E, Watanabe T, Osaki M, Wagatsuma T. Phosphorus deficiency enhances aluminum tolerance of rice (Oryza sativa) by changing the physicochemical characteristics of root plasma membranes and cell walls. Journal of Plant Physiology , 2014, 171 (2) : 9–15. DOI:10.1016/j.jplph.2013.09.012 |
| [20] | Kim S I, Kim D M, Tai T H. Evaluation of rice seedling tolerance to constant and intermittent low temperature stress. Rice Science , 2012, 19 (4) : 295–308. DOI:10.1016/S1672-6308(12)60054-7 |
| [21] | Agarwal S, Vgn T V, Kotla A, Mangrauthia S K, Neelamraju S. Expression patterns of QTL based and other candidate genes in Madhukar×Swarna RILs with contrasting levels of iron and zinc in unpolished rice grains. Gene , 2014, 546 (2) : 430–436. DOI:10.1016/j.gene.2014.05.069 |
| [22] | Pereira E G, Oliva M A, Rosado-Souza L, Mendes G C, Colares D S, Stopato C H, Almeida A M. Iron excess affects rice photosynthesis through stomatal and non-stomatal limitations. Plant Science , 2013, 201-202 : 81–92. DOI:10.1016/j.plantsci.2012.12.003 |