 2016, Vol. 36
2016, Vol. 36文章信息
- 郑瞳, 牟长城, 张毅, 李娜娜
- ZHENG Tong, MU Changcheng, ZHANG Yi, LI Nana.
- 立地类型对张广才岭天然白桦林生态系统碳储量的影响
- Effects of site condition on ecosystem carbon storage in a natural Betula platyphylla forest in the Zhangguangcai Mountains, China
- 生态学报[J]. 2016, 36(19): 6284-6294
- Acta Ecologica Sinica[J]. 2016, 36(19): 6284-6294
- http://dx.doi.org/10.5846/stxb201412122473
-
文章历史
- 收稿日期: 2014-12-12
- 网络出版日期: 2016-01-15





森林生态系统在植被、凋落物及土壤中储存大量碳,并通过光合和呼吸与大气进行大量碳交换,故森林在区域和全球陆地碳循环中发挥着关键作用[1]。森林生态系统碳储量主要受到气候、森林类型、林分年龄、干扰体制、土壤条件的强烈影响[2],同时,森林生态系统碳储量各组分(植被、凋落物、土壤)对这些控制因子的反应也不同[2-5]。因此,了解森林碳储量及其在各组分中分配的决定因素对预测碳平衡对气候变化与森林管理如何响应是极其重要的[2]。
目前国际上在区域和陆地尺度上,已有大量基于森林生物量或土壤调查研究揭示了森林生物量碳密度与土壤有机碳密度的地理分布格局,即随纬度增加(气候与植被水平分异),一般森林植被碳密度会降低,而土壤碳密度会增加[1, 6-8]。也有学者研究发现温带森林随海拔升高(气候与植被垂直分异),植被碳密度显著降低,凋落物碳密度先升高后降低趋势,土壤有机碳密度呈现增加趋势[9]。一些区域尺度火烧或采伐年龄序列研究揭示了林分年龄和干扰体制对温带和北方森林碳分布的影响,得到随着林分年龄增加,植被和土壤碳密度趋于增加,而凋落物碳密度一般呈现U-型格局[3-5, 10-12]。这些过往研究增进了对森林碳储量及其组分的影响因素的认识,但有关立地类型对森林碳储量及其组分分配的影响或立地分异规律性尚缺乏系统认识。同时,在森林生态系统碳储量研究方法中仍存在一些问题[13],例如,有些研究中地上植被碳储量和地下土壤碳储量往往是独立从不同生境中收集而来,然后将两者整合代表某个特定生态系统的碳储量[1, 14-15];又如,过去使用单一立地类型上所调查的生态系统碳储量数据也缺乏广泛代表性,主要是由于某一特定生态系统碳储量受到微气候和地形因素的影响,故调查数据会随立地条件的不同而发生改变[1, 16-21]。因此,有必要从生态系统尺度上同步测定植被碳储量与土壤碳储量,并考虑立地类型的影响。
本项目研究将重点探讨立地类型对我国东北林区主要次生林类型之一的白桦林碳汇的影响规律,目前国内有关东北林区白桦林碳汇研究报道相对较少,且大多集中在立地类型对白桦生长过程的影响方面[22-23],也有学者仅就单一立地类型上白桦林的地上植被碳储量[24-26]或地下土壤碳储量[27]进行过研究,或仅关注本区一些主要树种的单木生物量模型[28]及其碳含量的测定[29],而从生态系统水平上考虑立地类型对白桦林碳汇的影响研究则尚未见有报道。故目前对东北林区白桦林碳汇作用的立地分异规律性的认识仍不十分清楚,尚有必要开展相关研究工作。
本文以我国温带张广才岭天然白桦次生林为研究对象,采用相对生长方程与碳/氮分析法,同步测定白桦林在7个立地类型(阳坡上、中、下部和阴坡上、中、下部及谷地)上的生态系统碳储量(植被和土壤)、净初级生产力与年净固碳量,揭示立地类型对白桦林生态系统碳库及固碳能力的影响规律,以便为经营与管理东北森林的长期碳汇提供科学依据。
1 研究区概况研究区位于东北林业大学帽儿山国家级野外试验站老爷岭生态站,地理位置为45°2′20″—45°18′16″N,127°18′0″—127°41′6″E。属长白山系张广才岭西坡小岭余脉,为松嫩平原向张广才岭过渡的低山丘陵地区,坡度一般6°—15°。试验站总面积2.63万hm2,主要植被为东北东部山区典型天然次生林,森林覆被率70.2%,森林总蓄积2.049×106 m3。南北长30 km,东西长20 km,境内山峦绵延,丘陵起伏,最高海拨高度为805 m,平均海拔300 m,地势由南向北逐渐升高[30]。属于温带大陆性季风气候,年平均气温为2.7℃,平均相对湿度为71%,年降雨量为728 mm。日平均气温≥10℃的天数平均为152 d/a。地带性植被为红松阔叶林,现存植被为原始植被遭反复干扰破坏后形成的阔叶混交林为主的天然次生林,主要森林类型有蒙古栎林、山杨林、白桦林、谷地硬阔叶林等,是东北东部山区较典型的次生林区。地带性土壤为暗棕壤,包括多个亚类; 隐域性土壤包括草甸土、沼泽土和白浆土等[31]。各立地类型的林分基本特征见表 1。
| 立地类型Site type | 树种Tree species | 相对盖度Relative coverage/% | 相对密度Relative density/% | 相对频度Relative frequentness/% | 重要值Importance value | 平均胸径Mean DBH/cm | 平均年龄Mean age/a | 胸径范围DBH range/cm |
| 谷地Ⅰ | 白桦Betula platyphylla | 68.92 | 41.23 | 30.60 | 46.92 | 15.4 | 40 | 5.6—30 |
| 水曲柳Fraxinus mandschurica | 21.78 | 11.36 | 14.21 | 15.78 | 14.2 | 3.9—39.2 | ||
| 茶条槭Acer ginnala | 2.89 | 17.21 | 18.58 | 12.89 | 5.0 | 3.2—9 | ||
| 榆树Ulmus pumila | 2.53 | 10.06 | 12.02 | 8.20 | 6.1 | 3.4—9.7 | ||
| 阳上Ⅱ | 白桦Betula platyphylla | 57.72 | 20.24 | 19.85 | 32.60 | 21.0 | 45 | 12.1—41.2 |
| 五角槭Acer mono | 5.90 | 23.53 | 17.65 | 15.69 | 6.0 | 3.2—13.2 | ||
| 榆树Ulmus pumila | 3.30 | 19.53 | 18.38 | 13.74 | 5.1 | 2.9—9.9 | ||
| 枫桦Betula costata | 11.87 | 4.94 | 6.25 | 7.69 | 19.0 | 7.7—30.5 | ||
| 阳中Ⅲ | 白桦Betula platyphylla | 66.07 | 24.15 | 27.62 | 39.28 | 21.9 | 46 | 5.5—39.4 |
| 榆树Ulmus pumila | 10.27 | 50.15 | 34.25 | 31.56 | 5.9 | 3.2—11.6 | ||
| 水曲柳Fraxinus mandschurica | 8.68 | 8.36 | 12.71 | 9.91 | 11.4 | 4.7—31.5 | ||
| 落叶松Larix gmelinii | 9.48 | 4.95 | 5.52 | 6.65 | 17.3 | 4.8—38.6 | ||
| 阳下Ⅳ | 白桦Betula platyphylla | 68.28 | 29.36 | 26.13 | 41.26 | 20.1 | 44 | 10.7—33.8 |
| 榆树Ulmus pumila | 8.36 | 35.47 | 28.14 | 23.99 | 6.3 | 3.2—21.1 | ||
| 五角槭Acer mono | 5.48 | 14.24 | 16.58 | 12.10 | 7.9 | 3.1—15.0 | ||
| 水曲柳Fraxinus mandschurica | 5.79 | 9.88 | 13.57 | 9.75 | 8.7 | 3.8—33.2 | ||
| 阴上Ⅴ | 白桦Betula platyphylla | 45.93 | 15.44 | 15.00 | 25.46 | 26.9 | 52 | 9.7—40.0 |
| 五角槭Acer mono | 7.50 | 24.32 | 20.56 | 17.46 | 8.2 | 3.4—20.9 | ||
| 榆树Ulmus pumila | 7.08 | 14.67 | 12.78 | 11.51 | 10.2 | 3.5—25.3 | ||
| 山杨Populus davidiana | 15.99 | 6.95 | 8.89 | 10.61 | 24.4 | 15.0—33.0 | ||
| 阴中Ⅵ | 五角槭Acer mono | 36.66 | 13.75 | 16.67 | 22.36 | 7.4 | 3.1—22.3 | |
| 白桦Betula platyphylla | 11.28 | 42.01 | 27.22 | 26.84 | 24.3 | 49 | 6.4—37.8 | |
| 山杨Populus davidiana | 25.97 | 4.46 | 10.00 | 13.48 | 27.5 | 20.2—46.2 | ||
| 紫椴Tilia amurensis | 8.97 | 11.15 | 11.67 | 10.60 | 13.3 | 4.8—23.6 | ||
| 阴下Ⅶ | 白桦Betula platyphylla | 45.69 | 17.30 | 16.76 | 26.58 | 25.6 | 50 | 14.9—38.2 |
| 五角槭Acer mono | 6.70 | 32.87 | 22.91 | 20.82 | 6.5 | 3.0—17.7 | ||
| 榆树Ulmus pumila | 7.90 | 17.65 | 18.44 | 14.66 | 10.1 | 4.7—20.6 | ||
| 山杨Populus davidiana | 25.55 | 6.23 | 8.38 | 13.39 | 30.5 | 14.0—50.3 | ||
| Ⅰ: 谷地valley floor;Ⅱ、Ⅲ、Ⅳ: 阳坡上、中、下部 the top,middle,bottom of the sunny slope; Ⅴ、Ⅵ、Ⅶ: 阴坡上、中、下部the top,middle,bottom of shady slope | ||||||||
2013年4月在张广才岭东北林业大学帽儿山国家级野外试验站老爷岭生态站开展了立地类型对天然白桦林生态系统碳库及其固碳能力影响试验,试验选择林龄较大(40—50a)、分布较集中及人为活动干扰较轻(未进行采伐作业)的天然白桦林为研究对象,依据坡向与坡位组合确定了7种立地类型(阳坡上、中、下部和阴坡上、中、下部及谷地),并在每个立地类型上各设置3块20 m×30 m的固定标准地(3次重复),共计设置21块标准地。并于2013年生长初期(5月)及生长结束期(10月)对各标准地进行了植被、土壤及凋落物等方面的调查取样。
2.2 植被碳储量测定乔木层碳储量测定方法:依据各标准地乔木树种的每木调查数据(D≥2 cm),运用研究区已经建立的白桦、山杨、榆树、蒙古栎等树种的相对生长方程[28],计算各标准地乔木层的生物量,再利用研究区已测定的各树种碳含量数据[29],计算各标准地的乔木层碳储量。乔木各部分生物量及碳含量指标见表 2[28-29]。
| 树种 Species | 组分 Component | 生物量 Biomass | 碳含量 Carbon content | ||
| 生物量方程 Biomass equation | R2 | 均方误差MSE Mean squared error | |||
| 白桦Betula platyphylla | 干 | B=102.141D2.278 | 0.988 | 0.006 | 0.459 |
| 枝 | B=100.952D2.783 | 0.956 | 0.035 | 0.498 | |
| 叶 | B=101.176D1.942 | 0.918 | 0.033 | 0.489 | |
| 根 | B=101.319D2.530 | 0.993 | 0.004 | 0.457 | |
| 紫椴Tilia amurensis | 干 | B=101.491D2.686 | 0.988 | 0.010 | 0.543 |
| 枝 | B=100.564D2.847 | 0.908 | 0.093 | 0.531 | |
| 叶 | B=100.229D2.507 | 0.897 | 0.082 | 0.557 | |
| 根 | B=101.178D2.507 | 0.966 | 0.015 | 0.527 | |
| 山杨Populus davidiana | 干 | B=101.836D2.471 | 0.993 | 0.008 | 0.434 |
| 枝 | B=100.129D3.224 | 0.971 | 0.062 | 0.465 | |
| 叶 | B=100.567D2.182 | 0.795 | 0.243 | 0.471 | |
| 根 | B=100.992D2.563 | 0.996 | 0.005 | 0.435 | |
| 五角槭Acer mono | 干 | B=101.877D2.408 | 0.971 | 0.019 | 0.466 |
| 枝 | B=100.927D2.934 | 0.977 | 0.023 | 0.476 | |
| 叶 | B=101.235D1.948 | 0.962 | 0.017 | 0.491 | |
| 根 | B=102.081D1.989 | 0.975 | 0.012 | 0.45 | |
| 水曲柳Fraxinus mandshurica | 干 | B=102.116D2.316 | 0.991 | 0.008 | 0.532 |
| 枝 | B=100.963D2.785 | 0.974 | 0.037 | 0.525 | |
| 叶 | B=100.933D2.180 | 0.981 | 0.016 | 0.506 | |
| 根 | B=101.357D2.475 | 0.982 | 0.002 | 0.517 | |
| 落叶松Larix gmelinii | 干 | B=102.311D2.154 | 0.963 | 0.005 | 0.467 |
| 枝 | B=10-1.593D4.340 | 0.953 | 0.024 | 0.504 | |
| 叶 | B=10-1.851D3.934 | 0.887 | 0.052 | 0.492 | |
| 根 | B=101.061D2.572 | 0.949 | 0.009 | 0.469 | |
| B:生物量Biomass(kg); D: 胸径Diameter at breast height(cm) | |||||
灌木层、草本层及凋落物层碳储量测定方法:在各标准地内随机设置5个2 m×2 m灌木层小样方、10个1 m×1 m草本层小样方及10个30 cm×30 cm凋落物层小样方,采用收获法测定灌木层、草本层及凋落物层的生物量鲜重,并对其地上枝叶、地下根系及凋落物分别取样,然后放入70℃恒温烘箱烘干,测量各部分含水率,计算生物量干重;然后,将各干样品再用小钢磨粉碎,用分析天平(0.1mg)取50 mg左右,使用EA4000碳氮分析仪通过1300℃干烧法测定有机碳含量,再结合各层次生物量干重计算灌木层、草本层与凋落物层的碳储量。
2.3 土壤碳储量测定在各标准地内随机设置3个土壤剖面,按照10 cm间隔划分土壤层,在每个土壤层中利用土壤环刀取样3次,直至土壤母质层(各立地类型土壤层≤70 cm)。土壤样品称鲜重后自然风干,测量其干重,计算含水率。然后,取200 g左右土壤样品初步打碎,过100目筛处理,挑出其中的石砾、根系、杂物等,记录其中石砾所占的体积百分比。再经小钢磨粉碎后,用分析天平(0.1 mg)取50 mg左右粉碎后的样品,使用EA4000碳氮分析仪通过1300℃干烧法测定有机碳含量,计算土壤碳储量。
土壤有机碳密度是指单位面积一定深度的土层中SOC的贮量,一般用t/hm2或kg/m2 表示。是评价和衡量土壤中有机碳贮量的一个极其重要的指标。某一土层i的有机碳密度 (SOCi,kg/m2) 的计算公式为[27]:
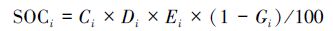
式中,Ci为土壤有机碳含量(g/kg),Di 为容重(g/cm3),Ei 为土层厚度(cm),Gi 为直径大于2 mm 的石砾所占的体积百分比(%)。
若某一土壤剖面由k层组成,那么该剖面的有机碳密度(SOCt,kg/m2) 为[27]:
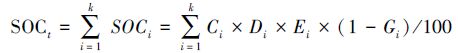
乔木层净初级生产力是依据各标准地在生长开始期(2013年5月)和生长停止期(10月)生物量的差值而得到;灌木层净初级生产力为其生物量除以平均年龄(5 a)[32-33];草本层净初级生产力为其当年的生物量;将三者加和即可得到各立地类型白桦林的植被净初级生产力。乔木层、灌木层、草本层的年净固碳量通过各自的年净初级生产力与其相应的碳含量的乘积获得,将三者加和即可得到各立地类型白桦林植被的年净固碳量。
2.5 数据处理所有所得数据使用Excel 2010 和IBM SPSS Statistics 19软件进行分析,采用单因素方差分析(ANOVA)、最小显著差异法(LSD)和Duncan 多重比较进行组间检验,显著性水平设置为α=0.05。
3 结果与分析 3.1 立地对白桦林植被碳储量的影响由表 3可知,立地类型对张广才岭天然白桦林植被碳储量具有显著影响。7种立地类型上天然白桦林植被碳储量分布在(76.28±18.11)—(115.57±5.59) tC/hm2之间,其中,阳坡上、下部和阴坡上、下部4个立地类型显著高于谷地35.1%—51.5%(P<0.05),阳坡中部与阴坡中部也高于谷地但差异性均不显著(32.5%—33.6%,P>0.05),总体上呈现出阳坡上、下部和阴坡上、下部>阳坡中部与阴坡中部>谷地的变化规律性。
| 指标 Item | 植被层 Layers | 立地类型Site type | ||||||
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ | ||
| 碳储量/(t/hm2) | 乔木层 | 72.12±17.66a | 101.62±20.81b | 93.66±7.35ab | 96.09±13.95ab | 103.32±12.56b | 94.87±10.09ab | 108.76±5.80b |
| Carbon storage | 灌木层 | 0.31±0.07ab | 0.18±0.04a | 0.25±0.04ab | 0.39±0.06b | 0.32±0.14ab | 0.34±0.13ab | 0.26±0.02ab |
| 草本层 | 0.40±0.40b | 0.06±0.01a | 0.04±0.03a | 0.06±0.04a | 0.31±0.19ab | 0.10±0.03ab | 0.07±0.00a | |
| 凋落物层 | 3.46±0.23a | 7.51±2.47b | 7.11±2.36b | 6.50±2.01ab | 6.77±2.08ab | 6.56±0.81ab | 6.48±0.46ab | |
| 植被 | 76.28±18.11a | 109.37±20.25b | 101.06±5.46ab | 103.05±15.98b | 110.72±14.61b | 101.87±10.76ab | 115.57±5.59b | |
| 分配比/% | 乔木层 | 94.42±1.00a | 92.70±2.60a | 92.60±2.76a | 93.36±0.99a | 93.41±1.25a | 93.12±0.15a | 94.10±0.59a |
| Allocation | 灌木层 | 0.42±0.18b | 0.16±0.02a | 0.25±0.05ab | 0.38±0.02b | 0.28±0.09ab | 0.34±0.17ab | 0.23±0.03ab |
| 草本层 | 0.47±0.37b | 0.06±0.00a | 0.04±0.03a | 0.06±0.05a | 0.28±0.17ab | 0.10±0.02a | 0.06±0.00a | |
| 凋落物层 | 4.69±10.30a | 7.08±2.57a | 7.11±2.68a | 6.20±1.05a | 6.03±1.30a | 6.44±0.16a | 5.62±0.57a | |
| 表中给出数据是平均值±标准差,不同小写字母表示不同立地类型之间植被碳储量的差异显著(P<0.05) | ||||||||
立地类型对白桦林植被各组成层次碳储量的影响规律也有所不同。乔木层碳储量在阳坡上部和阴坡上、下部显著高于谷地40.9%—50.8%(P<0.05),阳坡中、下部和阴坡中部高于谷地但差异性不显著(29.9%—33.2%,P>0.05),呈现阳坡上部和阴坡上、下部>阳坡中、下部与阴坡中部>谷地的变化趋势;灌木层碳储量仅在阳坡下部显著高于阳坡上部(116.7%,P<0.05),阳坡中部、阴坡各坡位及谷地也高于阳坡上部但差异性均不显著(38.9%—88.9%,P>0.05),呈阳坡上部>阳坡中部、阴坡各坡位及谷地>阳坡上部变化趋势;草本层碳储量在谷地显著高于阳坡各坡位与阴坡下部471.4%—900.0%(P<0.05),阴坡上、中部高于阳坡各坡位和阴坡下部但差异性均不显著(42.9%—675.0%,P>0.05),呈谷地>阴坡上、中部>阳坡各坡位及阴坡下部变化趋势;凋落物层碳储量在阳坡上、中部显著高于谷地105.5%—117.1%(P<0.05),阳坡下部和阴坡各坡位高于谷地但差异性均不显著(87.3%—95.7%,P>0.05),呈阳坡上、中部>阳坡下部和阴坡各坡位>谷地变化趋势。可见,仅乔木层碳储量随立地类型变化趋势与其植被相类似,而灌木层、草本层、凋落物层碳储量随立地类型变化趋势均与其植被有所不同。
3.2 立地对白桦林土壤有机碳储量的影响由表 4可知,立地类型对张广才岭天然白桦林土壤有机碳储量具有显著影响。7种立地类型上白桦林土壤有机碳储量分布在(81.53±6.15)—(181.90±21.62) tC/hm2之间,其中,阳坡各坡位(上、中、下部)显著高于阴坡中、下部与谷地24.0%—123.1%(P<0.05)(前三者也高于阴坡上部12.2%—23.1%,但仅阳坡中部提高显著),阴坡上、中部显著高于阴坡下部和谷地36.0%—81.2%,P<0.05),总体上呈现出阳坡各坡位>阴坡上、中部>阴坡下部和谷地的分布规律性。
| 土壤深度/cm Soil depth | 立地类型Site type | ||||||
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ | |
| 0—10 | 38.64±9.18 Ab | 51.14±3.61Cc | 37.41±3.12Cb | 36.73±2.72Cb | 40.96±1.89Bb | 41.36±1.92Cb | 24.86±9.55Aa |
| 10—20 | 26.67±7.12Aab | 42.38±4.08Ccd | 37.39±3.21Cbcd | 28.72±5.86BCabc | 44.67±13.84Bd | 27.28±8.96ABCab | 21.81±3.66Aa |
| 20—30 | 26.76±14.71Aab | 29.77±6.69Bab | 33.52±7.34BCb | 28.47±4.68BCab | 25.00±4.97Aab | 31.93±15.11BCab | 14.82±2.27Aa |
| 30—40 | 24.05±8.61ABa | 23.96±7.64ABa | 21.67±6.50ABa | 18.94±1.46Aa | 15.63±0.21Aa | 14.99±1.50Aa | |
| 40—50 | 19.39±0.12Aa | 19.76±2.19Aa | 18.96±5.66Aa | 18.19±5.25Aa | 17.46±0.30ABa | ||
| 50—60 | 22.46±9.39ABa | 13.89±3.50Aa | |||||
| 60—70 | 17.32±1.85Aa | ||||||
| 合计Total | 98.29±16.20a | 171.99±19.43cd | 181.90±21.62d | 165.76±16.99cd | 147.77±13.33bc | 133.67±7.55b | 81.53±6.15a |
| 不同大写字母表示不同土层深度SOC储量差异显著,不同小写字母表示不同立地类型SOC储量差异显著(P<0.05) | |||||||
立地类型对白桦林土壤有机碳储量的空间分布格局也具有影响。在水平分布上,0—10 cm土壤层的碳储量在阳坡上部显著高于其他6个立地类型23.6%—105.7%(P<0.05),阳坡中、下部和阴坡上、中部及谷地显著高于阴坡下部47.8%—66.4%(P<0.05),呈现阳坡上部>阳坡中、下部和阴坡上、中部及谷地>阴坡下部的变化趋势;10—20cm土壤层碳储量在阳坡上、中部和阴坡上部显著高于阴坡中、下部与谷地37.1%—104.8%(P<0.05),阳坡下部、阴坡中部及谷地高于阴坡下部但差异性均不显著(22.7%—31.7%,P>0.05),呈阳坡上、中部和阴坡上部>阳坡下部、阴坡中部及谷地>阴坡下部变化趋势;20—30 cm土壤层碳储量在阳坡中部显著高于阴坡下部126.2%(P<0.05),其他5个立地类型也高于阴坡下部但差异性均不显著(68.7%—115.5%,P>0.05),呈阳坡中部>谷地和阳坡上、下部及阴坡上、中部>阴坡下部变化趋势;30 cm以下各土壤层中的碳储量在不同立地类型之间差异性均不显著。可见,立地类型主要是改变了白桦林0—30 cm土壤层碳储量的水平分布格局,而对30 cm以下土壤层碳储量水平分布却无显著影响。
在垂直分布方面,谷地和阴坡下部2个立地类型的土壤层相对较薄(仅有3—4层),且各土壤层的碳储量并无显著差异性,分层不明显;阳坡上部和阴坡上部2个立地在0—20 cm土壤层的碳储量显著高于20—50 cm土壤层(42.4%—163.7%和63.8%—145.7%,P<0.05),可分为2层;阳坡中、下部和阴坡中部3个立地类型在0—30 cm土壤层的碳储量高于30 cm以下土壤层(39.9%—89.2%、31.4%—164.4%和56.2%—164.6%),也可分为2层。由此可见,张广才岭天然白桦林7种立地类型上土壤有机碳储量在水平分布、垂直分布格局方面均存在着较大的差异性。
3.3 立地对白桦林生态系统碳储量的影响由表 5可知,张广才岭天然白桦林生态系统碳储量存在着立地分异规律性。7种立地类型上白桦林生态系统碳储量分布在(174.57±20.27)—(282.96±17.92) tC/hm2之间,其中,阳坡上、中、下部显著高于阴坡中、下部与谷地14.1%—62.1%(P<0.05),阴坡上、中部显著高于阴坡下部与谷地19.5%—48.1%(P<0.05),呈现出阳坡各坡位>阴坡上、中部>阴坡下部与谷地的变化规律性。
| 指标 Item | 层次 Layers | 立地类型Site type | ||||||
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ | ||
| 碳储量/(t/hm2) | 植被 | 76.28±18.11a | 109.37±20.25b | 101.06±5.46ab | 103.05±15.98b | 110.72±14.61b | 101.87±10.76ab | 115.57±5.59b |
| Carbon storage | 土壤 | 98.29±16.20a | 171.99±19.43cd | 181.90±21.62d | 165.76±16.99cd | 147.77±13.33bc | 133.67±7.55b | 81.53±6.15a |
| 系统 | 174.57±20.27a | 281.36±8.74c | 282.96±17.92c | 268.80±14.71c | 258.49±19.44bc | 235.54±3.26b | 197.10±2.98a | |
| 分配比/% | 植被 | 43.61±7.89a | 38.85±7.05a | 35.86±3.71a | 38.33±5.54a | 42.79±4.10a | 43.21±4.00a | 58.64±2.89b |
| Allocation | 土壤 | 56.39±7.89b | 61.15±7.05b | 64.14±3.71b | 61.67±5.54b | 57.21±4.10b | 56.79±4.00b | 41.36±2.89a |
| 不同小写字母表示不同立地类型生态系统SOC储量差异显著(P<0.05) | ||||||||
立地类型对白桦林生态系统碳储量分配格局也有影响。在7种立地类型中,阳坡各坡位和阴坡上、中部及谷地的生态系统碳储量分布格局基本一致,土壤碳储量(56.4%—64.1%)与植被碳储量(35.9%—43.6%)的占比约为6∶4,呈现出土壤碳储量大于植被碳储量的分配格局;仅阴坡下部的生态系统碳储量分布格局有所不同,土壤碳储量(41.4%)与植被碳储量(58.6%)占比约为4∶6,呈现出植被碳储量大于土壤碳储量的分布格局。
3.4 立地对白桦林植被净初级生产力及年净固碳量的影响由表 6可知,立地类型对张广才岭天然白桦林植被净初级生产力和年净固碳量具有影响。7种立地类型上白桦林净初级生产力分布在(6.98±1.60)—(9.59±0.69) t hm-2 a-1之间,其中,阳坡上部与阴坡上部显著高于阴坡中部34.2%—37.4%(P<0.05),阳坡中、下部和阴坡下部及谷地也高于阴坡中部但差异性均不显著(8.5%—20.6%,P>0.05),呈现出阳坡上部与阴坡上部>阳坡中、下部和阴坡下部及谷地>阴坡中部的变化趋势;其年净固碳量分布在(3.26±0.74)—(4.56±0.36) tC hm-2 a-1之间,仅阳坡上部显著高于阴坡中部39.9%(P<0.05),其他5个立地类型也高于阴坡中部但差异性均不显著(9.2%—30.4%,P>0.05),呈现出阳坡上部>阳坡中、下部和阴坡上、下部及谷地>阴坡中部的变化趋势。因此,张广才岭天然白桦林在上坡位(阳坡上部与阴坡上部)具有相对较高的植被净初级生产力,在阳坡上部具有相对较高的植被年净固碳量。
| 指标 Item | 层次 Layer | 立地类型Site type | ||||||
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ | ||
| 净初级生产力 | 乔木层 | 6.34±0.54a | 9.28±0.66b | 7.97±1.22ab | 7.99±1.53ab | 8.01±0.50ab | 6.53±1.63a | 8.01±1.20ab |
| NPP/(t hm-2 a-1) | 灌木层 | 0.13±0.02abc | 0.08±0.01a | 0.11±0.02ab | 0.18±0.02c | 0.14±0.06abc | 0.15±0.05bc | 0.12±0.01ab |
| 草本层 | 1.09±0.84bc | 0.23±0.05ab | 0.14±0.10a | 0.24±0.19ab | 1.21±0.81c | 0.30±0.02ab | 0.23±0.03ab | |
| 植被层 | 7.57±1.34ab | 9.59±0.69b | 8.23±1.28ab | 8.42±1.36ab | 9.37±0.46b | 6.98±1.60a | 8.35±1.19ab | |
| 年净固碳量 | 乔木层 | 3.11±0.38a | 4.46±0.34b | 4.02±0.60ab | 3.99±0.77ab | 3.88±0.25ab | 3.09±0.74a | 3.82±0.49ab |
| ANCS/(t C hm-2 a-1) | 灌木层 | 0.06±0.01ab | 0.04±0.01a | 0.05±0.01ab | 0.08±0.01b | 0.06±0.03ab | 0.07±0.03ab | 0.05±0.00ab |
| 草本层 | 0.40±0.40b | 0.06±0.01a | 0.04±0.03a | 0.06±0.04a | 0.31±0.19ab | 0.10±0.02ab | 0.07±0.00a | |
| 植被层 | 3.56±0.74ab | 4.56±0.36b | 4.10±0.61ab | 4.12±0.75ab | 4.25±0.09ab | 3.26±0.74a | 3.95±0.49ab | |
| 不同小写字母表示不同立地类型植被NPP(net primary productivity)及ANCS(annual net carbon sequestration)差异显著(P<0.05) | ||||||||
立地类型对白桦林各植被层净初级生产力的影响规律有所不同。其乔木层净初级生产力仅在阳坡上部显著高于阴坡中部与谷地42.1%—46.4%(P<0.05),其他4个立地类型高于阴坡中部与谷地但差异性不显著(22.1%—26.3%,P>0.05),呈现阳坡上部>阳坡中、下部与阴坡上、下部>阴坡中部与谷地的变化趋势;灌木层净初级生产力在阳坡下部与阴坡中部显著高于阳坡上部87.5%—125.0%(P<0.05),其他4个立地类型高于阳坡上部但差异性不显著(37.5%—75.0%,P>0.05),呈阳坡下部与阴坡中部>阳坡中部与阴坡上、下部及谷地>阳坡上部变化趋势;草本层净初级生产力在阴坡上部和谷地显著高于阳坡中部678.6%—764.3%(P<0.05),其他4个立地类型高于阳坡中部但差异性不显著(64.3%—114.3%,P>0.05),呈阴坡上部和谷地>阳坡上、下部与阴坡中、下部>阳坡中部变化趋势。可见,白桦林乔木层、灌木层、草本层净初级生产力随立地类型的变化趋势完全不同,三者分别在阳坡上部、阳坡下部与阴坡中部、阴坡上部与谷地相对较高。
立地类型对白桦林各植被层年净固碳量的影响规律也有所不同。其乔木层年净固碳量在阳坡上部显著高于谷地和阴坡中部43.4%—44.3%,P<0.05),其他4个立地类型高于谷地和阴坡中部但差异性不显著(22.8%—30.1%,P>0.05),呈阳坡上部>阳坡中、下部与阴坡上、下部>阴坡中部和谷地变化趋势;灌木层年净固碳量在阳坡下部显著高于阳坡上部1.0倍(P<0.05),其他5个立地类型高于阳坡上部但差异性不显著(25.0%—75.0%,P>0.05),呈阳坡下部>阳坡中部、阴坡各坡位及谷地>阳坡上部变化趋势;草本层年净固碳量在谷地显著高于阳坡各坡位和阴坡下部4.7—9.0倍(P<0.05),阴坡上、中部也高于阳坡各坡位与阴坡下部但差异性不显著(42.9%—675.0%,P>0.05),呈谷地>阴坡上、中部>阳坡各坡位与阴坡下部变化趋势。可见,白桦林乔木层、灌木层、草本层年净固碳量随立地类型变化趋势也不同,三者分别在阳坡上部、阳坡下部、谷地相对较高。
4 结论与讨论 4.1 立地对白桦林植被碳储量的影响本研究得到张广才岭天然白桦林植被碳储量在阳坡、阴坡的上坡位与下坡位相对较高,中坡位居中等,谷地则最低。分析其原因,可能是由于植被碳储量主要取决于占其主体地位的乔木层碳储量(92.6%—94.4%)(表 3),乔木层碳储量在上坡位(阳坡、阴坡上部)和下坡位(阴坡下部)相对较高,而在谷地则相对较低,结果导致其植被碳储量呈现出与之相类似的变化趋势。具体原因则是由于上坡位的光照充足,加之白桦是阳性喜光树种,故上坡位有利于白桦林木的生长及碳素的积累过程;下坡位由于土壤湿度相对较大(水分充足),也有助于白桦林木的生长;而谷地在生长季经常处于季节性水淹状态,过多的土壤水分则限制了白桦林木的生长及其碳素积累过程,故导致谷地乔木层碳储量最低。
本研究得到张广才岭白桦林乔木层碳储量95.78 tC/hm2(72.12—108.76 tC/hm2)要高于现有研究结果(同为成熟林的小兴安岭白桦林乔木层碳储量67.37 tC/hm2及华北山区白桦林乔木层碳储量53.65 tC/hm2[34],韩国江原道白桦中龄林乔木层碳储量55.20 tC/hm2[35])42.2%—78.5%,其中,谷地白桦林乔木层碳储量(72.12 tC/hm2)略高于现有研究结果(7.1%—34.4%),而坡地白桦林乔木层碳储量(99.72 tC/hm2)较现有研究结果提高幅度较大(48.0%—85.9%),两者也高于黑龙江省森林乔木层碳储量平均值(33.44 tC/hm2[36])(115.7%—198.2%)。这说明张广才岭天然白桦林具有相对较高的乔木层碳储量。这可能与本区气候条件较为适宜、人为活动干扰强度较低(未进行采伐作业)及林分发育状况良好等有密切关系。
4.2 立地对白桦林土壤碳储量的影响本研究得到张广才岭天然白桦林土壤有机碳储量在阳坡各坡位相对较高,阴坡上、中部居于中等,阴坡下部和谷地相对较低的分布规律性。其主要原因是由于阳坡上、中、下部3个立地类型分别在0—20 cm、0—30 cm、0—10 cm土壤层中的碳储量相对较高(显著高于阴坡下部94.3%—105.7%、50.5%—126.2%和48.1%),且三者的土壤层均相对较厚(50—70 cm)(见表 3),故导致阳坡各坡位的土壤碳储量均相对较高;阴坡上、中部2个立地类型分别在0—20 cm、0—10 cm土壤层中的碳储量相对较高(显著高于阴坡下部64.8%—104.8%和66.8%),且两者的土壤层厚度属于中等水平(50 cm),故导致阴坡上、中部土壤有机碳储量居于中等;而阴坡下部和谷地2个立地类型不仅土壤层相对较薄(30—40 cm),且各土壤层碳储量也低,故导致两者土壤碳储量也相对较低。
此外,张广才岭7种立地类型上天然白桦林的土壤碳储量平均值140.13 tC/hm2(81.53—181.90 tC/hm2)高于我国天然林土壤碳储量109 t/hm2 [37]和北方森林土壤碳储量85 t/hm2 [32](28.6%—64.9%),低于吉林长白山森林土壤碳储量164.66 tC/hm2[38]及日本松本山森林土壤碳储量333.60 tC/hm2[13](14.9%—58.0%),故其土壤碳储量总体上应属于中等偏上水平;但从各立地类型来看,张广才岭白桦林土壤碳储量在阳坡立地上(173.22 tC/hm2)略高于长白山(5.2%),而在阴坡(120.99 tC/hm2)与谷地(98.29 tC/hm2)立地上其土壤碳储量则远低于长白山(26.5%—40.3%),这也说明白桦林土壤碳储量存在着明显的立地分异规律性。
4.3 立地对白桦林生态系统碳储量的影响本研究得到张广才岭天然白桦林生态系统碳储量在阳坡各坡位均相对较高,阴坡上、中部次之,阴坡下部与谷地最低。这与现有研究结论张广才岭天然白桦林生态系统碳储量在阳坡中坡位最高及谷地最低[22, 39]并不完全一致,即其生态系统碳储量在整个阳坡(上、中、下坡位)均相对较高,阴坡上、中部居中,而阴坡下部与谷地最低。分析其原因,主要是由于占白桦林生态系统碳储量主体地位的土壤碳储量随立地类型变化呈现出阳坡各坡位>阴坡上、中部>阴坡下部和谷地的分布规律性,导致其生态系统碳储量呈现出与之相类似的变化规律性,而植被碳储量在阳坡、阴坡各坡位上均高于谷地(32.5%—51.5%)且在各坡位之间差异性并不显著(表 5),故其对生态系统碳储量随立地类型变化趋势的影响不及前者。
同时,张广才岭7种立地类型天然白桦林生态系统碳储量平均值242.69 tC/hm2(174.57—282.96 tC/hm2)与长白山桦树林生态系统碳储量估计值(180.65—254.63 tC/hm2)[38] 的上限值接近(-4.7%)。说明张广才岭白桦林生态系统碳库略高于长白山的白桦林。
4.4 立地对白桦林植被净初级生产力和年净固碳量的影响本研究得到张广才岭天然白桦林的植被净初级生产力与年净固碳量分别在阳坡与阴坡的上坡位或阳坡上坡位相对较高。其原因主要在于阳坡上部白桦林乔木层净初级生产力和年净固碳量均最高(显著高于阴坡中部与谷地42.1%—46.4%和43.4%—44.3%),阴坡上部白桦林乔木层净初级生产力虽居中等,但其草本层的净初级生产力却最高(显著高于除谷地以外的其他5种立地类型3.0—7.6倍)(表 5),结果导致白桦林植被净初级生产力在阳坡与阴坡的上部相对较高,植被年净固碳量在阳坡上部相对较高。
此外,张广才岭天然白桦林的植被净初级生产力8.4 t hm-2 a-1(7.0—9.6 t hm-2 a-1)与现有中国东北森林植被净初级生产力(6.0—14.3 t hm-2 a-1)[40-41]的研究结论基本一致,高于其下限值(40.0%),但低于其上限值(-41.3%),说明其植被净初级生产力属于中等偏下水平。其植被年净固碳量4.0 t hm-2 a-1(3.3—4.6 t hm-2 a-1)略低于中国陆地植被固碳量(4.9 t hm-2 a-1)[42](-18.4%),但与全球平均植被固碳量(4.1 t hm-2 a-1)[43]相近(-2.4%)。因此,张广才岭天然白桦林的植被固碳能力接近全球植被平均固碳水平。
| [1] | Dixon R K, Solomon A M, Brown S, Houghton R A, Trexler M C, Wisniewski J. Carbon pools and flux of global forest ecosystems. Science , 1994, 263 (5144) : 185–190. DOI:10.1126/science.263.5144.185 |
| [2] | Pregitzer K S, Euskirchen E S. Carbon cycling and storage in world forests:biome patterns related to forest age. Global Change Biology , 2004, 10 (12) : 2052–2077. DOI:10.1111/gcb.2004.10.issue-12 |
| [3] | Gower S T, Vogel J G, Norman J M, Kucharik C J, Steele S J, Stow T K. Carbon distribution and aboveground net primary production in aspen, jack pine, and black spruce stands in Saskatchewan and Manitoba, Canada. Journal of Geophysical Research , 1997, 102 (D24) : 29029–29041. DOI:10.1029/97JD02317 |
| [4] | Law B E, Sun O J, Campbell J, Van Tuyl S, Thornton P E. Changes in carbon storage and fluxes in a chronosequence of Ponderosa pine. Global Change Biology , 2003, 9 (4) : 510–524. DOI:10.1046/j.1365-2486.2003.00624.x |
| [5] | Martin J L, Gower S T, Plaut J, Holmes B. Carbon pools in a boreal mixed wood logging chronosequence. Global Change Biology , 2005, 11 (11) : 1883–1894. |
| [6] | Jobbágy E G, Jackson R B. The vertical distribution of soil organic Carbon and its relation to climate and vegetation. Ecology Applications , 2000, 10 (2) : 423–436. DOI:10.1890/1051-0761(2000)010[0423:TVDOSO]2.0.CO;2 |
| [7] | Piao S L, Fang J Y, Zhu B, Tan K. Forest biomass carbon stocks in China over the past 2 decades, estimation based on integrated inventory and satellite data. Journal of Geophysical Research , 2005, 110 (G1) : G01006. |
| [8] | Yang Y H, Mohammat A, Feng J M, Zhou R, Fang J Y. Storage, patterns and environmental controls of soil organic carbon in China. Biogeochemistry , 2007, 84 (2) : 131–141. DOI:10.1007/s10533-007-9109-z |
| [9] | Zhu B, Wang X P, Fang J Y, Piao S L, Shen H H, Zhao S Q, Peng C H. Altitudinal changes in carbon storage of temperate forests on Mt Changbai, Northeast China. Journal of Plant Research , 2010, 123 (4) : 439–452. DOI:10.1007/s10265-009-0301-1 |
| [10] | Smithwick E A H, Harmon M E, Remillard S M, Acker S A, Franklin J F. Potential upper bounds of Carbon stores in forests of the Pacific Northwest. Ecology Applied , 2002, 12 (5) : 1303–1317. DOI:10.1890/1051-0761(2002)012[1303:PUBOCS]2.0.CO;2 |
| [11] | Wang C K, Gower S T, Wang Y H, Zhao H X, Yan P, Bond-Lamberty B P. The influence of fire on Carbon distribution and net primary production of boreal Larix gmelinii forests in north-eastern China. Global Change Biology , 2001, 7 (6) : 719–730. DOI:10.1046/j.1354-1013.2001.00441.x |
| [12] | Wang C K, Bond-Lamberty B, Gower S T. Carbon distribution of a well-and poorly-drained black spruce fire chronosequence. Global Change Biology , 2003, 9 (7) : 1066–1079. DOI:10.1046/j.1365-2486.2003.00645.x |
| [13] | Jia S G, Tsuyoshi A. A precise, unified method for estimating carbon storage in cool-temperate deciduous forest ecosystems. Agricultural and Forest Meteorology , 2005, 134 (1/4) : 70–80. |
| [14] | Adams J M, Faure H, Faure-Denard L, McGlade J M, Woodward F I. Increases in terrestrial carbon storage from the last glacial maximum to the present. Nature , 1990, 348 (6303) : 711–714. DOI:10.1038/348711a0 |
| [15] | Schlesinger W H. Soil organic matter:a source of atmospheric CO2//Woodwell G M, ed. The Role of Terrestrial Vegetation in the Global Carbon Cycle. New York:John Wiley & Sons Ltd , 1985 : 111–150. |
| [16] | Bohn H L. Estimate of organic carbon in world soils:Ⅱ. Soil Science Society of America Journal , 1982, 46 (5) : 1118–1119. DOI:10.2136/sssaj1982.03615995004600050050x |
| [17] | Cannell M G R. World Forest Biomass and Primary Production Data. London:Academic Press , 1982 . |
| [18] | Eswaran H, Van Den Berg E, Reich P. Organic carbon in soils of the world. Soil Science Society of America Journal , 1993, 57 (1) : 192–194. DOI:10.2136/sssaj1993.03615995005700010034x |
| [19] | Houghton R A, Hackler J L. Changes in terrestrial Carbon storage in the United States. Part 1:the roles of agriculture and forestry. Global Ecology and Biogeography , 2000, 9 (2) : 125–144. DOI:10.1046/j.1365-2699.2000.00166.x |
| [20] | Fang J Y, Chen A P, Peng C H, Zhao S Q, Ci L J. Changes in forest biomass carbon storage in China between 1949 and 1998. Science , 2001, 292 (5525) : 2320–2322. DOI:10.1126/science.1058629 |
| [21] | Garnett M H, Ineson P, Stevenson A C, Howard D C. Terrestrial organic carbon storage in a British moorland. Global Change Biology , 2001, 7 (4) : 375–388. DOI:10.1046/j.1365-2486.2001.00382.x |
| [22] | 王鹏, 王石磊, 王庆成, 李开隆. 帽儿山实验林场白桦适生立地条件. 东北林业大学学报 , 2010, 38 (10) : 9–11. |
| [23] | 柴一新. 天然白桦林的特点与经营. 东北林业大学学报 , 2000, 28 (5) : 31–34. |
| [24] | 宋熙龙, 毕君, 刘峰, 王超. 木兰林管局白桦次生林生物量与碳储量研究. 北京林业大学学报 , 2010, 32 (6) : 33–36. |
| [25] | 闫平. 帽儿山林场4类天然次生林碳储量研究. 林业资源管理 , 2006 (4) : 61–65. |
| [26] | 姜慧泉, 张会儒, 亢新刚. 长白山阔叶次生林主要乔木地上碳储量分布. 林业资源管理 , 2009 (5) : 58–63. |
| [27] | 杨金艳, 王传宽. 东北东部森林生态系统土壤碳贮量和碳通量. 生态学报 , 2005, 25 (11) : 2875–2882. |
| [28] | Wang C K. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests. Forest Ecology and Management , 2006, 222 (1/3) : 9–16. |
| [29] | Zhang Q Z, Wang C K, Wang X C, Quan X K. Carbon concentration variability of 10 Chinese temperate tree species. Forest Ecology and Management , 2009, 258 (5) : 722–727. DOI:10.1016/j.foreco.2009.05.009 |
| [30] | 东北林学院. 帽儿山实验林场基础资料. 哈尔滨:东北林学院 , 1984 : 1. |
| [31] | 潘建平, 赵克尊. 东北林业大学帽儿山实验林场林业资源增长与生态环境建设的研究. 哈尔滨: 东北林业大学出版社, 20071–2. |
| [32] | Giese L A B, Aust W M, Kolka R K, Trettin C C. Biomass and carbon pools of disturbed riparian forests. Forest Ecology and Management , 2003, 180 (1/3) : 493–508. |
| [33] | Mu C C, Lu H C, Wang B, Bao X, Cui W. Short-term effects of harvesting on carbon storage of boreal Larix gmelinii-Carex schmidtii forested wetlands in Daxing'anling, northeast China. Forest Ecology and Management , 2013, 293 : 140–148. DOI:10.1016/j.foreco.2012.12.031 |
| [34] | 马林. 小兴安岭白桦天然林生态系统碳储量的研究[D]. 哈尔滨:东北林业大学, 2012. http://cdmd.cnki.com.cn/article/cdmd-10225-1012445442.htm |
| [35] | Kim S W, Kang J T, Hwang J S, Lee S J, Park H, Son Y M. Stand growth analysis and carbon stocks/removals assessment on forest growth monitoring Plots in Korea. Journal of Agriculture & Life Science , 2014, 48 (6) : 11–19. |
| [36] | 焦燕, 胡海清. 黑龙江省森林植被碳储量及其动态变化. 应用生态学报 , 2005, 16 (12) : 2248–2252. |
| [37] | 刘世荣, 王晖, 栾军伟. 中国森林土壤碳储量与土壤碳过程研究进展. 生态学报 , 2011, 31 (19) : 5437–5448. |
| [38] | 王新闯, 齐光, 于大炮, 周莉, 代力民. 吉林省森林生态系统的碳储量、碳密度及其分布. 应用生态学报 , 2011, 22 (8) : 2013–2020. |
| [39] | 王鹏. 白桦适生立地条件研究[D]. 哈尔滨:东北林业大学, 2009. http://cdmd.cnki.com.cn/article/cdmd-10225-2009134517.htm |
| [40] | 周广胜, 张新时. 全球气候变化的中国自然植被的净第一性生产力研究. 植物生态学报 , 1996, 20 (1) : 11–19. |
| [41] | 毛德华, 王宗明, 罗玲, 韩佶兴. 1982-2009年东北多年冻土区植被净初级生产力动态及其对全球变化的响应. 应用生态学报 , 2012, 23 (6) : 1511–1519. |
| [42] | 何浩, 潘耀忠, 朱文泉, 刘旭拢, 张晴, 朱秀芳. 中国陆地生态系统服务价值测量. 应用生态学报 , 2005, 16 (6) : 1122–1127. |
| [43] | 李银鹏, 季劲钧. 全球陆地生态系统与大气之间碳交换的模拟研究. 地理学报 , 2001, 56 (4) : 379–389. |