 2016, Vol. 36
2016, Vol. 36文章信息
- 郑惠茹, 罗红霞, 邹扬庆, 程玉丝, 张锐
- ZHENG Huiru, LUO Hongxia, ZOU Yangqing, CHENG Yusi, ZHANG Rui.
- 基于地学信息图谱的重庆岩溶石漠化植被恢复演替研究
- Quantifying vegetation restoration in a karst rocky desertification area in Chongqing based on Geo-informatic Tupu
- 生态学报[J]. 2016, 36(19): 6295-6307
- Acta Ecologica Sinica[J]. 2016, 36(19): 6295-6307
- http://dx.doi.org/10.5846/stxb201411122233
-
文章历史
- 收稿日期: 2014-11-12
- 网络出版日期: 2016-01-15





岩溶石漠化现象是我国南方热带岩溶地区严峻的生态问题,据资料统计,截止2005年底,南方石漠化土地的总面积为12.96万km2[1],占该地区岩溶面积的28.7%。地学信息图谱是在继承中国传统图谱研究成果的基础上运用先进技术获取及其丰富的信息源,并以数据库为依托,通过交互式的操作系统,显示事物或现象的三维图形或空间动态变化,并建立相关模型的地理时空分析方法论[2-4]。继1998年陈述彭院士提出地学信息图谱的思想后,地学信息图谱无论是从理论、方法还是应用中都逐渐深入和成熟,其主要研究主要集中在对土地利用的时空动态格局演变[5-7]、城市空间的扩展研究[8-9]、生态景观的格局演变[10-11]和自然灾害的宏观分析和预测[12-13]等方面。国内外学者对植被群落演替的研究大都集中在使用单一技术手段来对演替的现象、规律和生理生态特性的研究,使用空间信息科学技术与生态学研究方法结合的较少;对中梁山的植被恢复研究也主要集中在使用生态学的方法对其生态恢复重建模式的研究。本研究在岩溶石漠化植被恢复区植被演替研究中引入地学信息图谱的相关理论方法,在获取1996、2001、2007和2013年4期遥感影像的基础上,结合野外实地所获取的高光谱数据,采用加入高程数据的神经网络分类方法对四个时相中不同的植被群落类型进行分类,并结合地学图谱的相关分析方法[14-16],探讨了中梁山植被演替时空变化规律,实现了空间-属性-过程的一体化,能够定量和直观地反映植被恢复过程中的演替时空变化规律。为生态工作者在植被恢复研究方面提供新的研究手段和技术方法,为研究区退化生态系统的恢复、重建与生态工程实施给予理论指导。
1 研究区域与数据来源 1.1 研究区概况为了探讨重庆市岩溶植被恢复区植被的演替规律与特征,本研究选取了位于重庆市市区北部的中梁山(106°18′14″—106°56′53″E,29°39′10″—10°3′53″N)约7354.8 hm2范围作为研究区域,该区域植被群落类型比较丰富,是石漠化植被恢复具有代表性的区域(图 1)。中梁山是川东平行岭谷主体山脉华蓥山的三条支脉之一,年均温18℃,年均降水量1000—1300 mm,属于亚热带湿润季风气候,相对湿度大;海拔400—700 m,坡度0—40°,属于中低山丘陵地区。由于中梁山缺土、缺水和偏碱性的岩溶环境导致该地区植物种多具有耐旱、石生和喜钙的特性。其中,中梁山轴部以旱生、喜钙性植物种类为主,如五节芒、黄花蒿等以及一些散生的落叶乔木树种,主要植被类型包括草丛、灌草丛、藤刺灌丛和乔幼灌丛。两翼以马尾松林和杉木次生林为主,形成较大面积的常绿针叶林和针阔混交林。竹林主要分布于居民居住区附近和山麓地区。

|
| 图 1 研究区地理位置 Fig. 1 Location of the research area |
考虑到8月份是该地区植被生长比较茂盛的时期,因此本文选取了1996年8月20日、2001年8月2日、2007年8月19日的Landsat TM影像和2013年8月19日的Landsat OLI影像共4期遥感影像作为基本数据,所有影像的轨道号均为128/39。影像数据来源于中国科学院计算机网络信息中心国际科学数据镜像网站(http://www.gscloud.cn)。其他数据包括:美国Analytical Spectral Devices公司生产的FieldSpec HandHeld 便携式光谱仪结合手持GPS机于2013年10月1—10日野外实测的主要植被群落的地物波谱数据,主要用于支持遥感图像的解译;吴征镒《中国植被》分类系统、1∶400万中国植被图分类系统、中国1∶100万陆地地表覆被分类系统和林业相关部门的森林资源调查专题矢量图和野外实测数据主要用于植被类型的确定和遥感影像的解译;重庆地理信息公共服务平台提供的大比例尺高清航拍影像图以及Quickbird高空间分辨率卫星影像图等主要用于后期的辅助解译和精度校准。
2 研究方法 2.1 植被分类信息提取(1) 数据的预处理
首先进行传感器端的辐射定标(获得地物的辅亮度图)、大气校正(获得地物实际反射率图)、图像增强处理和研究区裁剪等,并选用基于辐射传输理论的MODTRAN模型进行大气校正;然后用ViewSpecPro软件对野外采集的各个群落类型样本的地物波谱数据进行预处理并重采样到与遥感影像可见光-近红外-短波红外(450—900 nm)一致的波段;结合重采样后的高光谱曲线特征和野外实测的影像像元光谱的特点,最后选择OLI563(RGB)、TM452(RGB)波段进行假彩色影像合成,能明显地突出具有岩溶地质背景的地貌景观和不同用地类型的色调差异和纹理信息。
(2) 分类方案的获取
参考吴征镒《中国植被》分类系统、1∶400万中国植被图分类系统、中国1∶100万陆地地表覆被分类系统和本次研究重点即重庆中梁山岩溶石漠化区的植被恢复演替研究,并结合对野外实测光谱数据的光谱特征分析,最终确定研究区的遥感分类为:Ⅰ暖性针叶林:马尾松林、柏木林、杉木林;Ⅱ竹林:慈竹林;Ⅲ亚热带常绿落叶阔叶林(栓皮栎林、板栗林等);Ⅳ针阔混交林;Ⅴ灌丛(酸性土和碱性土上的常绿灌丛、矮林,常绿阔叶落叶乔幼灌丛,以及灌草丛);Ⅵ草丛共5大类8小类。其余的非植被包括耕地、人工建筑、采石场、水体4类。
(3) 遥感图像的解译分类
人工神经网络是大量简单神经元联接而成的非线性复杂网络系统并大量应用于遥感影像的分类研究[17]。本研究选择具有三层网络结构的BP神经网络模型,包括输入层、隐含层和输出层三层拓扑结构。BP算法通过网络对样本数据的自学习和利用学习结果对整个影像数据进行分类两个阶段来实现。首先,在训练样本选取的基础上确定网络系统,对相关控制参数和样本进行输入。根据2013年OLI多光谱数据的波普特性,分别选取OLI3、5、6波段为输入层节点,输出层节点数目设置为分类方案中的地物类别数即12,隐含层的节点数目选择从6开始并根据网络训练来不断调整。传递函数采用logistic非线性函数,训练学习的样本数据采用传统监督分类时的样本数据,控制参数主要包括学习率(0,1)、动量因子(0,1),权值和阈值的初始化根据经验值随机设定。经过大量的学习和相关参数的多次调整,最终确定在学习率η=0.2、动量因子α=0.9、训练1500次和隐含层为1时,达到多次调整后的较佳精度。由于精度的局限性,在之后的试验中,尝试使用OLI2、3、4、5、6、7波段为输入层节点,并分别加入可能影响分类精度的阴影影响参数RA5/4(OLI5/OLI4)和RA7/3(OLI7/OLI3)、纹理特征值(均值、方差、均匀值、对比度、相异性和熵)、高程、坡度、坡向等因子分别进行试验;然后,在保证小类别有足够的分析点的前提下,采用简单随机采样法对随机样本进行采样。
(4) 精度评价
验证样本主要来源于研究区野外实地调查所收集到的实测资料,植被群落调查样方如图 2,林业部门所提供的研究区森林资源调查专题矢量图和Quickbird高空间分辨率卫星影像图以及2012年中梁山土地利用现状图,通过经纬度定位和目视判读选定2106个检验样本并一一确定像元点的实际类别;最后,采用混淆矩阵对分类结果进行定量评价。结果表明,运用加入高程数据的神经网络分类方法对4个时相中不同的植被群落类型进行分类所取得效果最好,分类总精度达到87.42%,总体精度比传统监督分类的结果提高了5.57%,分类混淆矩阵如表 1。使用同样的分类方法对前3期影像进行解译分类,最终分类结果如图 3。

|
| 图 2 研究区植被群落调查样方分布图 Fig. 2 The distribution of plots in research area |
| 分类类型Category type | 马尾松 Pi | 杉木 Ch | 柏木 Ka | 竹林 Ba | 阔叶林 Br | 针阔混交林 Co | 灌丛 Sh | 草丛 Gr | 人工建筑 Ac | 采石场 Qu | 耕地 Al | 水体 Wa | 合计 Total | 错分误差/% Misclassification deviation | 用户精度/% User accuracy |
| 马尾松Pi | 873 | 61 | 0 | 30 | 0 | 38 | 0 | 0 | 0 | 0 | 0 | 0 | 1016 | 14.07 | 85.93 |
| 杉木Ch | 1 | 45 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 46 | 2.17 | 97.83 |
| 柏木Ka | 0 | 0 | 134 | 0 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 140 | 4.29 | 95.71 |
| 竹林Ba | 0 | 0 | 0 | 1055 | 0 | 0 | 16 | 0 | 0 | 0 | 1 | 0 | 1072 | 1.59 | 98.41 |
| 阔叶林Br | 85 | 8 | 92 | 17 | 227 | 19 | 26 | 0 | 0 | 0 | 0 | 0 | 474 | 52.11 | 47.89 |
| 针阔混交林Co | 0 | 0 | 0 | 0 | 0 | 37 | 0 | 0 | 0 | 0 | 2 | 0 | 39 | 5.13 | 94.87 |
| 灌丛Sh | 0 | 0 | 24 | 16 | 15 | 0 | 289 | 1 | 0 | 0 | 1 | 0 | 346 | 16.47 | 83.53 |
| 草丛Gr | 0 | 0 | 5 | 0 | 1 | 0 | 10 | 126 | 3 | 1 | 8 | 0 | 154 | 18.18 | 81.82 |
| 人工建筑Ac | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 199 | 0 | 0 | 0 | 202 | 1.49 | 98.51 |
| 采石场Qu | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 8 | 266 | 0 | 1 | 275 | 3.27 | 96.73 |
| 耕地Al | 0 | 0 | 0 | 0 | 6 | 8 | 18 | 0 | 42 | 24 | 897 | 0 | 998 | 10.12 | 89.88 |
| 水体Wa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 141 | 143 | 1.40 | 98.60 |
| 合计Total | 959 | 114 | 258 | 1122 | 291 | 102 | 359 | 127 | 254 | 291 | 909 | 142 | 4906 | ||
| 漏分误差/% Leakage deviation | 8.97 | 60.53 | 48.06 | 5.97 | 15.61 | 63.73 | 19.50 | 0.79 | 21.65 | 8.59 | 1.32 | 0.70 | |||
| 制图精度/% Cartographic accuracy | 91.03 | 39.47 | 51.94 | 94.03 | 84.39 | 36.27 | 80.50 | 99.21 | 78.35 | 91.41 | 98.68 | 99.30 | |||
| 总体精度=87.4236% Overall accuracy=87.4236%; Kappa 系数=0.85Kappa coefficient=0.85;Pi:马尾松 Pinus massoninana;Ch:杉木 Chinese fir;Ka:柏木 Kashiwagi;Ba:竹林 Bamboo forest;Br:阔叶林 Broad forest;Sh:灌丛 Shrub;Co:针阔混交林 Coniferous and broad-leaved mixed forest;Gr:草丛 Grass;Ac:人工建筑 Artificial construction;Qu:采石场 Quarry;Wa:水体 Water area;Al:耕地 Arable land | |||||||||||||||

|
| 图 3 不同时相遥感影像植被分类图 Fig. 3 The classification for remote sensing images in four period |
(1) 地学信息图谱分析法
地学信息图谱以图像、图表的形式来反映事物和现象的形态结构、成因机制、动态变化等综合性、复杂性的规律[2-4, 18-19]。图谱单元是将不同时刻所采集到的空间数据进行融合或代数运算从而得到的空间、时间和属性等一体化的数据[20]。
(2) 转移矩阵法
转移矩阵可以定量化地识别不同地类在不同时间上的空间格局变化。在图谱的基础上通过ArcGIS软件进行数据的融合、叠置分析、面积的计算和转移矩阵的制作等,得到定量化的图谱变化数据。转移矩阵的数学表达式如下:
 (1)
(1)
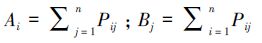 (2)
(2)
式中,Pij表示地类i转变为地类j的转移量,Ai表示t1时期第i种地物类型的面积;Bj表示t2时期第j种地物类型的面积。
(3) 质心迁移法
地类分布质心的变化情况可以反映在一定历史发展时期中地类在空间上的聚散与迁移[12]。质心坐标的计算公式如下:
 (3)
(3)
式中,Xt、Yt分别表示在t年某地类分布区域的质心坐标,Cti表示在t年某地类分布区域第i斑块的面积,Xi、Yi分别表示在t年该地类第i斑块的质心坐标。分布质心迁移距离采用欧氏距离来量算。
3 结果与分析 3.1 植被群落面积变化分析经过对多源数据的处理分析,得到植被群落的面积变化(图 4)和变化幅度情况(图 5)。从图中可知:(1)在1996年,耕地和马尾松在研究区内所占的比重相对较大(分别占研究区总面积的29.50%和24.52%),而2013年,马尾松、耕地、灌丛和阔叶林所占比重较大(分别占研究区总面积的28.16%、19.98%、20.55%和15.01%);(2)1996—2001年间,马尾松、阔叶林、竹林、灌丛4种主要植被的面积均呈减少趋势,发生了逆行演替的现象。其中竹林减少幅度最大(较1996年减少了57.32%),马尾松、阔叶林和灌丛次之,这主要与人们对竹林和松林的砍伐有关,从这个时期的草本群落的明显增加也可以看出。自2002年“退耕还林”工程全面启动后,植被恢复工作成果显著,在2001—2007年之间,马尾松面积增加幅度最大(较2001年增加了15.49%),2007—2013年灌丛增加幅度最大(较2007年增加了111.00%),而同时这两个阶段里耕地的减少幅度最大。(3)退耕还林后即2001—2013年,植被群落中马尾松、灌丛和阔叶林呈增加的趋势且增加幅度大小依次是灌丛>马尾松>阔叶林;而柏木、针阔混交林、竹林和草地呈减少的趋势且减少幅度大小依次是草丛>竹林>柏木>针阔混交林。

|
| 图 4 研究区主要植被群落及非林地面积变化图 Fig. 4 The variation map of the main vegetation communities and nonforest in the study area |

|
| 图 5 不同覆被类型面积在3个阶段中的变化幅度 Fig. 5 The change of different cover types are in three stages |
在遥感影像地物信息提取的基础上,构建了研究区退耕还林后(2001—2013)植被恢复演替变化图谱(图 6),并根据地表覆被类型转移矩阵制作了研究区主要植被群落变化图谱单元排序表(表 2)。结合图表的信息可知:在该时期内,研究区地表覆被发生变化的总面积为3862.62 hm2,变化区域和未变化区域呈斑块状交错分布,总变化强度为52.52%(总变化面积/研究区总面积)。具体来讲:植被演替在该时期内存在两个方向即进展演替和逆行演替。进展演替的比例为81.37%,主要表现为耕地经过自然恢复演替、人工辅助演替等转变为了草本群落、灌丛群落、阔叶林群落和柏木群落,占进展演替的50.31%,演替面积分别为233.46、730.80、249.93、28.53 hm2;草本群落演替发展为灌丛群落、阔叶林群落和柏木群落,占进展演替的16.22%,演替面积分别为257.94、111.60、30.96 hm2;灌丛群落发展为阔叶林群落、马尾松群落、竹林群落,占进展演替的10.44%,演替面积分别为138.69、68.49、50.58 hm2;马尾松群落则来源于阔叶林、竹林、针阔混交林、灌丛和草本群落的进展演替,共计550.26 hm2。由于岩溶区特殊的地质环境和外界的人为干扰,使得部分植被发生逆向演替在所难免,虽然所占比例很小,但却使植被演替研究变得复杂。

|
| 图 6 植被群落演替图谱(2001—2013年) Fig. 6 The succession map of main vegetation communities (2001—2013) |
| 面积排序 Area rank | 图谱单元面积 Change area/hm2 | 占总变化比例 Proportion in total change/% | 累计变化 Grand proportion in total change/% | 植被群落的主要变化类型 Main change type |
| 1 | 730.80 | 24.08 | 24.08 | 耕地→灌丛群落 |
| 2 | 361.26 | 11.90 | 35.98 | 常绿落叶阔叶林群落→马尾松群落 |
| 3 | 257.94 | 8.50 | 44.48 | 草本群落→灌丛群落 |
| 4 | 249.93 | 8.23 | 52.71 | 耕地→常绿落叶阔叶林群落 |
| 5 | 233.46 | 7.69 | 60.40 | 耕地→草本群落 |
| 8 | 138.69 | 4.57 | 64.97 | 灌丛→常绿落叶阔叶林群落 |
| 9 | 111.60 | 3.68 | 68.65 | 草本群落→常绿落叶阔叶林群落 |
| 10 | 74.52 | 2.46 | 71.10 | 竹林群落→马尾松群落 |
| 11 | 68.49 | 2.26 | 73.36 | 灌丛群落→马尾松群落 |
| 13 | 50.58 | 1.67 | 75.03 | 灌丛群落→竹林群落 |
| 14 | 41.58 | 1.38 | 76.40 | 人工建筑→灌丛群落 |
| 16 | 30.96 | 1.02 | 77.42 | 草本群落→柏木群落 |
| 18 | 28.53 | 0.94 | 78.36 | 耕地→柏木群落 |
| 19 | 26.01 | 0.86 | 79.22 | 针阔混交林群落→马尾松群落 |
| 20 | 22.77 | 0.75 | 79.97 | 人工建筑→常绿落叶阔叶林群落 |
| 21 | 22.5 | 0.74 | 80.71 | 人工建筑→草本群落 |
| 22 | 19.98 | 0.66 | 81.37 | 人工建筑→灌丛群落 |
| 6 | 164.34 | 5.41 | 86.78 | 常绿落叶阔叶林群落→灌丛群落* |
| 7 | 143.64 | 4.73 | 91.52 | 马尾松群落→常绿落叶阔叶林群落* |
| 12 | 61.29 | 2.02 | 93.54 | 常绿落叶阔叶林群落→竹林群落* |
| 15 | 40.77 | 1.34 | 94.88 | 柏木群落→常绿落叶阔叶林群落* |
| 17 | 30.69 | 1.01 | 95.89 | 竹林群落→灌丛群落* |
| *植被逆行演替变化部分 | ||||
由表 2可知,从2001—2013年研究区内植被恢复演替过程中,由最初的耕地转变为草本群落所占的变化比例最大,但在研究区内的部分区域已经构成了一个完整的植被恢复演替序列(图 7)。由图 7可知,在2001年为耕地的区域经过6a的自然恢复过程先演变为灌草丛(2007年),并在此基础上进一步演变为阔叶林、马尾松等乔木种群(2013年),其中2001—2007年间、2007—2013年间植被的变化强度分别为14.99%和28.24%。

|
| 图 7 研究区内典型植被恢复样区图 Fig. 7 sample-lands of typical vegetation in study area |
将4个时期不同演替阶段的植被群落质心坐标进行叠合,形成了植被群落质心分布图(图 8)和质心迁移图(图 9),以及基于欧式距离的植被群落质心迁移距离和迁移方向表(表 3)。

|
| 图 8 不同演替阶段植被群落的质心分布 Fig. 8 Centroid distribution of different succession stages of vegetation community |

|
| 图 9 不同演替阶段植被群落的质心迁移方向 Fig. 9 Centroid migration direction of different succession stages of vegetation community |
| 主要植被群落类型 Vegetation types | 1996—2001 | 2001—2007 | 2007—2013 | 2001—2013 | ||||
| 迁移距离 Migration distance/m | 迁移方向 Migration direction | 迁移距离 Migration distance/m | 迁移方向 Migration direction | 迁移距离 Migration distance/m | 迁移方向 Migration direction | 迁移距离 Migration distance/m | 迁移方向 Migration direction | |
| 马尾松Pi | 185.0 | 北偏东 | 196.4 | 南偏西 | 192.7 | 正北 | 28.3 | 正南 |
| 柏木Ka | 917.2 | 南偏西 | 619.1 | 北偏东 | 1972.2 | 南偏西 | 1353.3 | 南偏西 |
| 阔叶林Br | 727.2 | 南偏东 | 435.3 | 北偏西 | 367.7 | 南偏东 | 425.4 | 南偏西 |
| 针阔混交林Co | 3735.2 | 南偏西 | 2680.0 | 北偏东 | 1083.4 | 正南 | ||
| 竹林Ba | 389.9 | 东南 | 1804.2 | 东北 | 207.7 | 西南 | 1599.8 | 北偏东 |
| 灌丛Sh | 511.0 | 东南 | 1252.9 | 西南 | 1206.7 | 正北 | 905.8 | 西北 |
| 草丛Gr | 895.1 | 正北 | 664.7 | 东北 | 3347.3 | 南偏西 | 2791.7 | 南偏西 |
由图表信息可知,1996年和2001年,除柏木林外的其他群落类型质心基本集聚在了研究区的几何中心位置;2007年作为过度阶段,除柏木群落、竹林质心偏北而针阔混交林、灌丛群落质心偏南外,阔叶林、马尾松、草本群落质心居中;到2013年,阔叶林、灌丛群落、马尾松群落质心又集聚在了研究区中部,说明在1996—2013年间阔叶林和马尾松林在该区域内分布均匀且生长均衡。植被群落质心多在南北方向上迁移,这与该区南北走向的地势有关,且在2001—2013年间,质心迁移距离依次由大到小为草本群落>竹林群落>柏木群落>针阔混交林>灌丛群落>常绿落叶阔叶林群落>马尾松群落。具体表现为:马尾松群落和阔叶林群落的质心仅仅是在南北方向略有变动,并没有一个明显的迁移变化方向;柏木群落在2001—2007年期间质心向北迁移,在2007—2013年又向南移动,这表明在该时期里原质心所在地南部出现了柏木群落的分布,这与中梁山中部海石公园封山育林下的柏木自然恢复和退耕坡地上人工种植柏木的实际情况相符;常绿针阔混交林作为阔叶林到针叶林的过渡阶段,其分布质心在2001—2007年期间明显南移,在2007—2013年又向北移,但总体上针阔混交林还是向南迁移,说明这个时期研究区南部有更多的阔叶林群落在向针叶林群落演进;竹林的分布质心从2001—2007年向东北方向迁移,之后变化一直较小,这是由于中梁山的竹林以速生慈竹林群落为主;灌丛群落的质心迁移较明显,2001—2013年整体向西北方向移动,且移动距离较大,说明在研究区的西北方向上有较多的非灌丛群落转变为了灌丛群落,且增加面积较大;草本群落的质心迁移也很显著,1996—2001年向北移动,之后又向东北移动,2007—2013年长距离的向南偏西方向迁移,说明草本群落在这个时期里变化最大。总的来说,植被群落质心多在南北方向上迁移,这与该区南北走向的地势有关。
4 结论与讨论 4.1 主要研究结论本研究是在区域尺度的基础上,结合3S技术、野外实测高光谱技术及生态学方法实现区域尺度的植被类型划分。分类试验结果和分类精度表明:①植被演替识别分类中涉及到多个时期植被种群的识别,中分辨率多光谱影像混合像元问题又不可避免,BP神经网络较强的自学习能力和高度的容错能力,以及BP神经网络的并行式分布结构可以很方便地引入非光谱信息参与分类,使得遥感影像分类精度达到87.42%,比传统监督分类精度提高了5.57%,但在区分杉木林、柏木林和针阔混交林精度依然很低,存在错分和误分的现象;②在研究时段内,研究区域植被群落在不同的时间段内面积变化特征明显。1996—2001年,以竹林的减少为主,2001—2007年,以马尾松林的增加为主,2007—2013年,以灌丛的增加为主,分别占各变化时期植被变化面积的36.36%、38.47%和55.75%。从演替序列来看,中梁山岩溶石漠化地区的植被恢复整体上遵循了旱生进展演替的一般规律,即草本群落到灌木群落再到乔木群落,但是由于中梁山属于中低山地区,受人为影响因素大,研究区内海拔高差相对较小加之土壤稀薄不均的生境条件的限制,使得该地区的顶级群落更多的表现为土壤顶级[21-22]。人工林的种植加速了该区域的进展演替,但由于其物种构成和群落结构比较简单,不如自然恢复植被稳定,更容易被破坏和入侵。因此,在进行该地区的生态功能的恢复时,选择和引入自然恢复演替过程中后期的植物种如柏木、马尾松等对中梁山石漠化地区进行植被重建应该更有利于该地区稳定植被生态系统的形成。
4.2 讨论①仅有光谱数据的网络分类完全不能识别杉木、柏木和针阔混交林,而随着其他类型波段(比值波段、纹理、坡度、坡向、高程)的加入整个分类精度才有所提高,但并不是影响因子的全部加入就会起到提高分类精度的作用,反而可能造成一定的干扰。②由于数据源、野外采集高光谱仪的局限性和研究区地形等方面的影响,使得在对研究区不同演替阶段具有代表性群落的划分比较粗略,例如常绿落叶阔叶林并未给出具体的建群种或共优种类型。此外,本研究只是重点探讨了小区域尺度上人工神经网络方法在植被群落划分和演替中的应用,在实际工作中,还应根据研究对象、研究目的和数据源的不同,尝试将多源数据多方法相结合的分类方法以提高分类精度。③从所得到的结果看,自国家在西南喀斯特地区实施退耕还林还草等一系列生态建设工程特别是2008年国务院批复了《岩溶地区石漠化综合治理大纲(2006—2015)》以来,喀斯特地区植被恢复变化明显。中梁山属中低山地区,受人类活动影响大,加之特殊的地质环境和薄厚不均的土壤分布,使得该区域的植被恢复情况有特定的变化特征。变化较显著的是中梁山两侧外围的耕地和中轴山丘的耕地,而未变化植被群落类型主要是分布在中梁山“两翼”酸性土壤上的顶级群落——马尾松(少量杉木)群落和分布在钙质土壤上的顶级群落——柏木群落,它们具有较好的能量物质转换能力和稳定的生态系统功能,因此在自然演替过程中变化小。④植被恢复演替图谱很好地诠释了地学信息图谱的特点和内涵,能够定量和直观地反映各植被类型在不同演替阶段的空间格局、分布规律和变化过程,图谱单元可以研究各影像单元在各采样时刻的利用状态,实现空间-属性-过程的一体化,可见地学信息图谱在分析植被恢复演替的时空变化方面具有非常好的图形表现形式和分析手段,从而为岩溶石漠化地区的植被恢复提供强有力的科学依据和技术手段。
| [1] | 国家林业局. 岩溶地区石漠化状况公报. 中国绿色时报 , 2006 : 6–23. |
| [2] | 陈述彭, 岳天祥, 励惠国. 地学信息图谱研究及其应用. 地理研究 , 2000, 19 (4) : 337–343. |
| [3] | 廖克. 地学信息图谱的探讨与展望. 地球信息科学 , 2002 (1) : 14–20. |
| [4] | 齐清文. 地学信息图谱的最新进展. 测绘科学 , 2004, 29 (6) : 15–23. |
| [5] | 张国坤, 邓伟, 张洪岩, 宋开山, 李恒达. 新开河流域土地利用格局变化图谱分析. 地理学报 , 2010, 65 (9) : 1111–1120. |
| [6] | 孙晓芳, 岳天祥, 范泽孟. 中国土地利用空间格局动态变化模拟——以规划情景为例. 生态学报 , 2012, 32 (20) : 6440–6451. |
| [7] | 孙倩, 塔西甫拉提·特依拜, 张飞, 丁建丽, 买买提·沙吾提, 韩桂红, 桂东伟. 渭干河-库车河三角洲绿洲土地利用/覆被时空变化遥感研究. 生态学报 , 2012, 32 (10) : 3252–3265. |
| [8] | 韩晨. 基于地学信息图谱的西安城市空间扩展研究[D]. 西安:陕西师范大学, 2007. http://cdmd.cnki.com.cn/article/cdmd-10718-2007199643.htm |
| [9] | 李锋, 叶亚平, 宋博文, 王如松. 城市生态用地的空间结构及其生态系统服务动态演变——以常州市为例. 生态学报 , 2011, 31 (19) : 5623–5631. |
| [10] | 雷璇, 杨波, 蒋卫国, 杨一鹏, ClaudiaK, 陈强. 东洞庭湿地植被格局变化及其影响因素. 地理研究 , 2012, 31 (3) : 461–470. |
| [11] | 叶庆华, 田国良, 刘高焕, 叶景敏, 娄维国. 黄河三角洲新生湿地土地覆被演替图谱. 地理研究 , 2004, 23 (2) : 257–264. |
| [12] | 陈菁, 廖克. 基于地学信息图谱的福建省生态环境脆弱性分析. 世界地理研究 , 2009, 18 (2) : 169–176. |
| [13] | 安静. 区域滑坡灾害危险性评价信息图谱研究[D]. 长沙:中南大学, 2009. http://cdmd.cnki.com.cn/article/cdmd-10533-2009237605.htm |
| [14] | Xiao J Y, Ge J F, Shen Y J, Liang Y Q. Research on land use/cover change in Shijazhuang using landsat TM and ETM+ data. Geographical Science , 2005, 25 (4) : 495–500. |
| [15] | Mulders M A. Advances in the application of remote sensing and GIS for surveying mountainous land. International Journal of Applied Earth Observation and Geoinformation , 2001, 3 (1) : 3–10. DOI:10.1016/S0303-2434(01)85015-7 |
| [16] | Soares-Filho B S. Dinamica-a stochastic cellular automata model designed to simulate the landscape dynamics in an Amazonian colonization frontier. Ecological Modelling , 2002, 154 (3) : 217–235. DOI:10.1016/S0304-3800(02)00059-5 |
| [17] | 李双成, 郑度. 人工神经网络模型在地学研究中的应用进展. 地球科学进展 , 2003, 18 (1) : 68–76. |
| [18] | 韦玉春, 汤国安, 杨昕. 遥感数字图像处理教程. 北京: 科学出版社, 2011235. |
| [19] | 田永中, 岳天祥. 地学信息图谱的研究及其模型应用探讨. 地球信息科学 , 2003 (3) : 103–106. |
| [20] | Ye Q H, Tian G L, Liu G H, Ye J M, Yao X, Liu Q S, Lou W G, Wu S G. Tupu methods of spatial-temporal pattern on land use changes:A case study in the Yellow River Delta. Journal of Geographical Sciences , 2004, 14 (2) : 131–142. DOI:10.1007/BF02837528 |
| [21] | 范文武, 陈晓德, 李加海, 张淑琴, 李艳霞. 重庆中梁山海石公园石灰岩山地植物多样性研究. 西南大学学报:自然科学版 , 2009, 31 (5) : 106–110. |
| [22] | 贾红杰, 傅瓦利, 赵俊丽, 张文晖, 甄晓君, 杜富芝, 马志敏, 段正峰. 重庆中梁山岩溶区耕作侵蚀影响因素研究. 水土保持学报 , 2008, 22 (1) : 28–32. |