 2016, Vol. 36
2016, Vol. 36文章信息
- 刘翔, 周宏飞, 刘昊, 朱海
- LIU Xiang, ZHOU Hongfei, LIU Hao, ZHU Hai.
- 不同类型生物土壤结皮覆盖下风沙土的入渗特征及模拟
- Characteristic and numerical simulation of sandy soil infiltration under the different types of biological soil crusts
- 生态学报[J]. 2016, 36(18): 5820-5826
- Acta Ecologica Sinica[J]. 2016, 36(18): 5820-5826
- http://dx.doi.org/10.5846/stxb201507211531
-
文章历史
- 收稿日期: 2015-07-21
- 修订日期: 2016-04-21





2. 中国科学院大学, 北京 100049;
3. 中国科学院阜康荒漠生态系统研究站, 阜康 831505
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Fukang Station of Desert Ecology, Chinese Academy of Sciences, Fukang 831505, China
在干旱区, 土壤水分条件是制约荒漠生态系统植被演替的关键因素[1], 水分入渗作为水文循环中的重要一环, 是降水等地表水转化为土壤水的唯一途径, 土壤入渗能力深刻影响着降水的再分配过程[2], 对降水的有效储存与转化利用至关重要。在干旱荒漠地区广泛分布的BSCs(生物土壤结皮)是由真菌、细菌、藻类、地衣、藓类等孢子植物类群及其代谢产物与土壤表层颗粒复合形成的结构十分复杂的复合体[3], 是荒漠生态系统重要的组成部分, 具有重要的生态水文学意义[4]。BSCs的出现明显的改变了沙土表层的土壤特性[5]与覆被状况, 这势必影响到土壤水分入渗过程。近年来, 国内外学者对BSCs与水分入渗的关系进行了研究[6-7], 但已有研究因试验方法、试验区域、实验对象等不同, 相关结论并不统一, 部分学者认为BSCs促进了土壤水分入渗[8], 另有一部分学者则持相反的观点[9-10]。可见, 相关实验证据还不够充分, 导致很多研究结论并不一致。同时, 已有研究中针对不同类型BSCs覆盖下的沙土水分入渗比较实验还不多见, 相关研究有待加强。
古尔班通古特沙漠作为我国最大的固定与半固定沙漠, 其间广泛发育有不同类型的BSCs, 部分学者已对其分布特征[11]、形成机理与演替过程[12]、生态功能及效应[13-14]等问题进行了一些研究, 但针对不同类型BSCs入渗特征的比较研究还未见报导。本项研究以古尔班通古特沙漠广泛发育的藻结皮、地衣结皮和藓类结皮等3类结皮为研究对象, 采用原状土柱入渗实验和模拟方法, 对不同类型BSCs的入渗特征展开研究, 可以为进一步了解BSCs对土壤水文过程的调控过程和机理提供科学依据。
1 研究区概况古尔班通古特沙漠位于北疆准噶尔盆地腹地, 面积4.88×104 km2, 地理位置为44°11′—46°20′ N, 84°31′—90°00′ E, 是中国最大的固定、半固定沙漠。该地区属温带干旱气候区, 夏季干燥炎热、冬季寒冷, 年均气温6.6℃。年均降水量90—150 mm, 沙漠腹地仅有70—100 mm, 年蒸发量(Φ20cm蒸发皿)2000 mm以上;冬季有20—30 cm的稳定积雪覆盖, 雪融水以垂直入渗为主[15], 无地表径流形成。由于降水季节分配较均匀以及春季雪融水的存在, 沙漠内植物具有多样性和稳定性的特点[16]。研究区发育有良好的BSCs, 盖度达到41.34%[17], 有藻结皮、地衣结皮和藓类结皮等3种类型的结皮存在。研究区具有以呈西北—东南走向树枝状沙垄为主的地貌格局, 土壤为风沙土, 垄顶有宽约1 m的流动沙带, 结构性差, 垄间沙土结构性较好, BSCs多分布于垄间[11]。实验样地位于古尔班通古特沙漠南缘的中国科学院阜康荒漠生态站北沙窝试验场内, 地理坐标44°23′ N, 87°55′ E。
2 材料与方法 2.1 样品采集选取人为扰动少、不同类型BSCs均发育良好的丘间低地作为样地, 共3块, 每块样地的面积为25 m2(5 m × 5 m), 在样地内按S型分别布设3个样方进行土壤容重、含水量、厚度、孔隙度等指标的调查计算, 结果如表 1。
| 结皮类型 BSCS |
土壤容重 Bulk density/ (g/cm3) |
最大持水量 Maximum water-holding capacity/% |
毛管持水量 Capillary moisture/% |
非毛管孔隙度 Non-capillary porosity/% |
毛管孔隙度 Capillary porosity/% |
土壤重量含水量 Water content/% |
结皮厚度 Thickness of crusts/mm |
| 藻结皮Algal crust | 1.52±0.03 | 28.11±0.41 | 25.06±0.82 | 2.19±0.23 | 38.17±0.19 | 1.18±0.19 | 2.04±0.38 |
| 地衣结皮Lichen crust | 1.53±0.05 | 30.42±0.37 | 26.23±0.27 | 2.12±0.25 | 40.14±0.18 | 1.55±0.40 | 8.38±0.72 |
| 藓类结皮Moss crust | 1.49±0.02 | 35.54±0.32 | 27.48±0.02 | 2.55±0.18 | 41.06±0.03 | 1.47±0.30 | 17.70±1.51 |
| 无结皮(CK)Without crust | 1.59±0.01 | 26.51±0.21 | 22.27±0.82 | 6.79±0.11 | 34.59±0.18 | 0.94±0.13 | 无 |
| 文中数据均采自于沙土表层(0—5cm) | |||||||
选择各样地内BSCs分布最具优势、覆盖最完全的位置取样, 样品为包含BSCs均匀覆盖的原状风沙土土柱, 在同一地点取裸沙(CK)样品作为对照。取样前用喷壶喷水湿润地表, 可防止结皮层在取样时破裂, 待水渗入表层土壤后, 将高(h)为25 cm, 内径(r)为6.3 cm的有机玻璃管垂向砸入土壤中, 之后剥离周边土壤, 在底部横切放置铁板后, 取出包含原状土样的样品。样品取回在室内搁置48 h, 自然干燥后的土柱样品可以供室内入渗模拟实验使用。3种类型结皮和裸沙土各重复取样4次, 共计16个样品。
2.2 实验方法采用室内土柱模拟实验模拟入渗过程。实验开始前, 用纱布将有机玻璃管及其底部铁片一同包裹, 水平放置于铁架台上, 在土柱表面放置滤纸。由于砂质土壤入渗率较大, 马氏瓶恒定供水方法供水不便, 因此采用人工供水方法供水。土壤表面刻度作为零, 土柱两侧垂向贴附纸尺, 记录湿润锋到达不同深度的时间;土柱下放置漏斗和烧杯, 收集穿透土柱的渗漏水分。实验在中国科学院阜康荒漠试验站实验室进行。
实验过程中控制水头高度在2 cm, 随时加水, 用量筒测量每一时间段内所加水量。前20 min内每隔1 min测1次所加水量, 之后每隔5 min测1次;同时, 记录柱体两侧湿润锋到达不同深度的时间。每次模拟实验进行90 min(所有处理在此时段内均达稳渗)。为消除实验时水温差异带来的影响, 用温度计测定实验时段水层温度, 将测得的入渗速率Kθ按照式(1)进行统一换算为10℃时入渗速率K:
 (1)
(1)
式中, θ为渗透测定时的水温(℃), Kθ为水温为θ时的渗透速率(mm/min)。
2.3 测定及计算内容初渗速率(mm/min)实验开始后第1分钟添加水量。
稳渗速率(mm/min)实验开始后, 前20 min每隔1 min测量1次添加水量, 后每隔5 min测量1次, 当前后供水量恒定或相差不大时视为入渗过程达到稳渗阶段, 并通过加水量与间隔时间算出稳渗速率, 达到稳渗阶段所需时间为稳渗时间。
湿润锋运移过程记录湿润锋到达不同深度的时间。
渗漏时间各处理自实验开始至底部出现渗漏水的时间。
1 h累积入渗量自实验开始1 h内的累积加水量。
2.4 入渗过程模拟国内外学者相继提出了多种描述水分入渗过程的数学模型, 按模型中参数是否具有物理意义将其区分为两类:一类是参数不具备实际物理意义的经验模型如Kostiakov模型, 另一类是物理模型如Horton入渗模型和Philip入渗模型等[18]。本研究选用以上3种入渗模型, 利用统计分析软件进行拟合, 探讨其对本实验不同类型BSCs覆盖下土壤入渗过程的实用性。
Kostiakov模型:
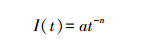 (2)
(2)
Horton模型:
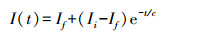 (3)
(3)
Philip模型:
 (4)
(4)
式中, I(t)为入渗率(mm/min);t为入渗时间(min);Ii为初始入渗率(mm/min); If为稳定入渗率(mm/min)a、b、c、n为模型参数。
采用SPSS 18.0进行方差分析, 采用Origin 8.5进行图形制作与模型拟合。
3 结果与分析 3.1 不同类型BSCs覆盖下风沙土壤入渗过程3种类型BSCs入渗过程见图 1(图中y轴为“入渗速率”的对数形式, 其反映的规律不变)。从图中可以看出, 各处理土壤水分入渗曲线均是双曲线形式, 并且具有相近的变化趋势。

|
| 图 1 3种类型BSCs覆盖下风沙土入渗过程对数曲线 Fig. 1 Logistic curve of sandy soil infiltration process under threedifferent biological soil crusts |
在入渗过程的初始阶段是水分渗润阶段, 入渗过程主要在分子力的作用下进行, 此时的土壤水分入渗速率称为初渗速率。由表 2可知, 不同类型BSCs覆盖均显著降低了初渗速率, 藓类结皮、地衣结皮、藻结皮覆盖下初渗速率降低幅度依次为36.10%、46.42%、50.39%。BSCs层持水性能较强[8], 导致了其表层土壤初始含水率比CK组高(表 1), 而初渗速率随土壤初始含水率的增加而降低[19], 因而各类型BSCs均降低了初渗速率。
| 结皮类型 BSCs |
初渗速率 Initial infiltration rate / (mm/min) |
稳渗速率 Stable infiltration rate/(mm/min) |
到达稳渗时间 Time to stable infiltration rate/min |
1h累积入渗量 Cumulative infiltration volume per hour/mL |
渗漏时间 Leakage time |
初渗速率 标准误 Standard error of initial infiltration rate |
稳渗速率 标准误 Standard error of stable infiltration rate |
| 藻结皮Algal crust | 7.85* | 0.65* | 30—35 | 158.67 | 25′55″ | 0.25 | 0.01 |
| 地衣结皮Lichen crust | 8.48* | 0.52* | 30—35 | 157.75 | 26′29″ | 0.52 | 0.04 |
| 藓类结皮Moss crust | 10.12 | 0.8 | 20—25 | 203.00 | 18′31″ | 0.80 | 0.06 |
| 无结皮(CK)Without crust | 15.83 | 1.01 | 25—30 | 249.50 | 8′42″ | 0.42 | 0.04 |
| *号代表与CK组差异显著 | |||||||
在入渗的第二阶段——渗漏阶段, 此时水分主要在毛细管力和重力作用下逐步填充土壤空隙。由图 1可知, 在渗漏阶段中某一时段(图中入渗曲线交叉部分)内, 各类型BSCs覆盖下沙土入渗速率均大于CK对照组, 这是因为没有BSCs覆盖的沙土的入渗性能强于BSCs覆盖下的沙土, CK对照组入渗速率衰减速度更快, 较早的降低到了比较低的水平(1.49—1.80mm/min)。
当土壤空隙被水分充满达到饱和后, 入渗过程便进入到第三阶段——渗透阶段, 到达这一阶段的时间称为稳渗时间。此阶段中, 入渗速率稳定在一个比较固定的水平上, 即稳渗速率。由表 2可见, 与CK组相比, 地衣结皮和藻结皮均显著(P<0.05)降低了稳渗速率, 降幅依次为33.98%和35.92%, 藓类结皮也明显降低稳渗速率, 降幅为16.50%。稳渗速率降低的可能原因:不同类型BSCs层在形成过程中均使沙土表层孔隙不同程度的被细粒物质填充, 从而减少了沙土中非毛管孔隙度(表 1), 相对于CK裸沙来说, 其表层土壤结皮相当于一个堵塞层, 从而降低了土壤入渗能力;而地衣结皮的壳状覆盖与藻结皮致密的结构使其具有封闭的特征[12], 使这两类结皮覆盖的沙土稳渗速率减小程度更大。
3.2 不同类型BSCs对湿润锋运移过程的影响湿润锋的位置和运移速率是反映水分垂向运动的特征之一[20]。图 2是湿润锋随时间变化过程线, 不同处理下, 湿润锋运移过程呈现出相似的趋势, 即随着入渗深度的增加, 湿润锋运移过程线逐渐平缓, 运移速率呈先快后慢的规律。在不同入渗阶段, 各处理下湿润锋运移速率均不相同, CK组湿润锋平均运移速率明显高于有BSCs覆盖的情况, 从各处理渗漏时间(表 2)可看出类似规律, 藓类结皮、地衣结皮、藻结皮的渗漏时间分别为CK组的2.13、3.04和2.98倍。以上BSCs的性质可能对本地降水的有效转化产生深刻的影响:在干旱荒漠区, 次降水中, 若水分不能及时渗入深层区域, 处在表层沙土的水分很快会被蒸发损失掉, 从而降低降水有效性。

|
| 图 2 不同类型BSCs覆盖下湿润锋运移过程 Fig. 2 The migration process of wetting front under three different biological soil crusts |
BSCs对湿润锋运移过程的影响主要是通过其较高持水性实现。在形成过程中, BSCs通过菌丝和假根网罗或粘结沙粒, 并借此形成水稳性团聚体, 从而增加了土壤中有机质的含量, 提高了土壤持水性[21], 将更多入渗水分储存于表面结皮层;部分藻类还可在细胞外分泌粘性物[22], 阻塞一些基质孔, 从而阻止入渗, 以上BSCs的水文物理特性会延缓水分下渗过程, 最终影响了湿润锋在土柱中的运移。而藓类结皮与地衣结皮、藻结皮之间渗漏时间存有区别的原因可能在于其表面结构:藓类结皮表层为单株藓类植物体密集丛生为主[12], 其株间间隙为水流进入下层土壤提供了良好通道。同时, 藓类结皮遇水会发生膨胀, 可进一步加大株间间隙, 而地衣结皮与藻结皮表层均比较致密, 阻碍了水分进入土体。
3.3 不同类型BSCs对土壤累积入渗量的影响累积入渗量随入渗过程逐渐增加。由表 2可知, 3种类型BSCs覆盖下土壤1 h累积入渗量均显著(P < 0.05)小于CK组。如图 3, 3种类型BSCs减渗的程度不一, 藓类结皮、地衣结皮、藻结皮的1 h累积入渗量分别比CK组减少16.10%、28.56%和26.56%。地衣结皮与藻结皮减渗作用更显著的可能原因:相对于藓类结皮, 地衣结皮与藻结皮的表层结构更封闭, 结皮层越封闭, 减渗作用更加明显。同时, BSCs对土壤累积入渗量的影响与其对稳渗速率影响规律一致, 这也说明累积入渗量主要受稳渗速率的影响, 受初渗速率影响较弱。

|
| 图 3 3种类型BSCs 1h累积入渗量降低率 Fig. 3 The reduction rate of one hour cumulative-infiltration underdifferent biological soil crusts |
本研究选用3种经典的入渗模型, 用Origin 8.5软件进行拟合, 探讨其对本实验中不同类型BSCs覆盖下土壤入渗过程的实用性, 结果见表 3。
| 结皮类型BSCS | Kostiakov模型Kostiakov model | Horton模型Horton model | Philip模型Philip model | |||||
| I(t)=at-b | R2 | I(t)=If+(Ii-If)e-t/c | R2 | I(t)=a+bt-0.5 | R2 | |||
| 藻结皮Algal crust | I(t)=7.04t-0.639 | 0.95 | I(t)=0.66+7.19e-t/3.734 | 0.81 | I(t)=6.717t-0.5-0.35 | 0.92 | ||
| 地衣结皮Lichen crust | I(t)=7.11t-0.681 | 0.87 | I(t)=0.68+7.80e-t/2.823 | 0.74 | I(t)=6.429t-0.5-0.33 | 0.82 | ||
| 藓类结皮Moss crust | I(t)=8.98t-0.693 | 0.90 | I(t)=0.86+9.26e-t/3.426 | 0.81 | I(t)=8.346t-0.5-0.55 | 0.85 | ||
| 无结皮(CK)Without crust | I(t)=15.16t-0.933 | 0.96 | I(t)=1.03+14.80e-t/2.849 | 0.90 | I(t)=13.231t-0.5-1.60 | 0.83 | ||
由表 3可见, 在本实验条件下, 使用Kostiakov方程对不同处理下土壤入渗过程拟合结果显示, 参数a值在7.14—15.16之间, a值越大, 入渗曲线的斜率越大, 瞬时入渗率衰减越快, 其衰减快慢顺序:CK>藓类结皮>地衣结皮>藻结皮, 这与图 1显示结果一致;而参数b没有实际物理意义, 其变化范围在0.639—0.933之间。Horton模型参数c值在2.823—3.734之间, 最大值出现在藻结皮, 最小值在地衣结皮, 反映了入渗速率递减情况。Philip模型中A为稳渗速率, 将各处理A值带入求得参数b, b值在3.95—6.81之间, 它在一定程度上反映了初始入渗率的大小, 即藓类结皮覆盖下土壤初渗速率大, 而地衣结皮及藻结皮覆盖下初渗速率小。
对入渗过程的拟合效果可以用回归方程的决定系数R2表示, 其值越大, 拟合效果越好。由表4可以看出, Kostiakov模型对本实验不同处理入渗过程的模拟效果都较好, R2都在0.85以上。对于Kostiakov模型, 当t→∞时I(t) →0仅在水平吸渗时出现, 但此实验中时间t是有限的, 因此Kostiakov模型可以用来模拟本实验入渗过程。Horton模型对4种处理的拟合效果也较好, 除地衣结皮(R2=0.74)处理外, 其余处理R2在0.81以上。Horton虽是纯经验公式, 但能描述长时间的入渗特征, 且模型参数具有物理意义, 是描述本研究入渗过程的较好模型。Philip模型是在半无限均质土壤并有积水条件下求得, 只适用于均质土壤一维入渗, 不适用于BSCs覆盖下的非均质土壤, 因此对本实验入渗过程的模拟效果较差。
由以上分析可知, 在本实验条件下, Kostiakov模型最适用于模拟不同类型BSCs覆盖下土壤水分入渗过程, Horton模型次之。此外, 结皮覆盖下的二元结构土壤, 其入渗过程与结皮发育程度、土壤理化性质等关联, 本实验所得拟合方程及其系数能否具有通用性还有待检验和讨论。
4 结论(1)不同类型BSCs均阻碍了风沙土水分入渗, 但阻碍程度差别较大。3种类型BSCs均显著降低了风沙土初渗速率, 藓类结皮、地衣结皮、藻结皮覆盖下初渗速率降低幅度依次为36.10%、46.42%、50.39%、同时, 藓类结皮、地衣结皮(P < 0.05)、藻结皮(P < 0.05)均明显降低了风沙土稳渗速率, 稳渗速率降低幅度依次为16.50%、33.98%和35.92%。
(2)本研究中, 不同类型BSCs均限制了湿润锋在风沙土的推进过程, CK组湿润锋平均运移速率明显高于有BSCs覆盖的情况, 表现为:藓类结皮、地衣结皮、藻结皮的渗漏时间分别为CK组的2.13、3.04和2.98倍。
(3)不同类型BSCs均显著减小了1 h累积入渗量。与无结皮对照相比, 藓类结皮、地衣结皮、藻结皮1 h累积入渗量分别降低16.10%、28.56%和26.56%;地衣结皮与藻结皮减渗作用相仿, 大于藓类结皮的减渗作用。
(4)本实验条件下, Kostiakov最适用于模拟不同类型BSCs覆盖下土壤水分入渗过程;Horton模型模拟效果也较好;Philip模型并不适用于模拟本实验水分入渗过程。
| [1] | 赵文智, 程国栋. 干旱区生态水文过程研究若干问题评述. 科学通报 , 2001, 46 (22) : 1851–1857. |
| [2] | 王新平, 李新荣, 康尔泗, 张景光, 周海燕, 杨诗秀, 雷志栋. 腾格里沙漠东南缘人工植被区降水入渗与再分配规律研究. 生态学报 , 2003, 23 (6) : 1234–1241. |
| [3] | 李新荣, 张元明, 赵允格. 生物土壤结皮研究:进展、前沿与展望. 地球科学进展 , 2009, 24 (1) : 11–24. |
| [4] | Bowker M A, Maestre F T, Escolar C. Biological crusts as a model system for examining the biodiversity-ecosystem function relationship in soils. Soil Biology and Biochemistry , 2010, 42 (3) : 405–417. DOI:10.1016/j.soilbio.2009.10.025 |
| [5] | Zhang Y M. The microstructure and formation of biological soil crusts in their early developmental stage. Chinese Science Bulletin , 2005, 50 (2) : 117–121. |
| [6] | Fischer T, Yair A, Veste M. Infiltration, water holding capacity and growth patterns of biological soil crusts on sand dunes under arid and temperate climates. Vienna:Egu General Assembly , 2012, 14 : 5034. |
| [7] | 李守中, 肖洪浪, 宋耀选, 李金贵, 刘立超. 腾格里沙漠人工固沙植被区生物土壤结皮对降水的拦截作用. 中国沙漠 , 2002, 22 (6) : 612–616. |
| [8] | Xiao B, Wang Q H, Zhao Y G, Shao M A. Artificial culture of biological soil crusts and its effects on overland flow and infiltration under simulated rainfall. Applied soil ecology , 2011, 48 (1) : 11–17. DOI:10.1016/j.apsoil.2011.02.006 |
| [9] | Wang X P, Li X R, Xiao H L, Berndtsson R, Pan Y X. Effects of surface characteristics on infiltration patterns in an arid shrub desert. Hydrological Processes , 2007, 21 (1) : 72–79. DOI:10.1002/(ISSN)1099-1085 |
| [10] | 徐敬华, 王国梁, 陈云明, 卢建利. 黄土丘陵区退耕地土壤水分入渗特征及影响因素. 中国水土保持科学 , 2008, 6 (2) : 19–25. |
| [11] | 张元明, 陈晋, 王雪芹, 潘惠霞, 辜智慧, 潘伯荣. 古尔班通古特沙漠生物结皮的分布特征. 地理学报 , 2005, 60 (1) : 53–60. |
| [12] | 张元明, 王雪芹. 荒漠地表生物土壤结皮形成与演替特征概述. 生态学报 , 2010, 30 (16) : 4484–4492. |
| [13] | 李卫红, 任天瑞, 周智彬, 刘加珍. 新疆古尔班通古特沙漠生物结皮的土壤理化性质分析. 冰川冻土 , 2005, 27 (4) : 619–626. |
| [14] | 王雪芹, 张元明, 张伟民, 韩致文. 古尔班通古特沙漠生物结皮对地表风蚀作用影响的风洞实验. 冰川冻土 , 2004, 26 (5) : 632–638. |
| [15] | 周宏飞, 李彦, 汤英, 周宝佳, 徐宏伟. 古尔班通古特沙漠的积雪及雪融水储存特征. 干旱区研究 , 2009, 26 (3) : 312–317. |
| [16] | 张立运, 陈昌笃. 论古尔班通古特沙漠植物多样性的一般特点. 生态学报 , 2002, 22 (11) : 1923–1932. |
| [17] | 吴林, 张元明. 以数码照相法估算生物土壤结皮盖度. 中国沙漠 , 2013, 33 (6) : 1810–1815. |
| [18] | Dashtaki S G, Homaee M, Mahdian M H, Kouchakzadeh M. Site-Dependence performance of infiltration models. Water Resources Management , 2009, 23 (13) : 2777–2790. DOI:10.1007/s11269-009-9408-3 |
| [19] | 刘目兴, 聂艳, 于婧. 不同初始含水率下粘质土壤的入渗过程. 生态学报 , 2012, 32 (3) : 871–878. |
| [20] | 陈洪松, 邵明安, 王克林. 土壤初始含水率对坡面降雨入渗及土壤水分再分布的影响. 农业工程学报 , 2006, 22 (1) : 44–47. |
| [21] | 曹成有, 蒋德明, 全贵静, 耿莉, 崔振波, 骆永明. 科尔沁沙地小叶锦鸡儿人工固沙区土壤理化性质的变化. 水土保持学报 , 2004, 18 (6) : 108–111. |
| [22] | 李守中, 肖洪浪, 李新荣, 宋耀选, 刘立超. 干旱、半干旱地区微生物结皮土壤水文学的研究进展. 中国沙漠 , 2004, 24 (4) : 500–506. |