 2016, Vol. 36
2016, Vol. 36文章信息
- 周延山, 花立民, 楚彬, 刘丽, 姬程鹏, 田永亮
- ZHOU Yanshan, HUA Limin, CHU Bin, LIU Li, JI Chengpeng, TIAN Yongliang.
- 祁连山东段高原鼢鼠对高寒草甸危害评价
- Assessment of damage caused by plateau zokor to an alpine meadow in eastern Qilian Mountain
- 生态学报[J]. 2016, 36(18): 5922-5930
- Acta Ecologica Sinica[J]. 2016, 36(18): 5922-5930
- http://dx.doi.org/10.5846/stxb201507211529
-
文章历史
- 收稿日期: 2015-07-21





高原鼢鼠(Myospalax baileyi)是青藏高原优势鼠种之一, 对高寒草地生态系统有着重要影响[1]。近年来, 由于气候变化和人为因素, 部分区域高原鼢鼠种群密度激增导致鼠害发生, 严重影响了当地的农牧业生产和生态安全[2]。余晓华和刘荣堂认为高原鼢鼠挖洞掘土行为破坏土壤和地表结构并引发水土流失, 是草地质量降低的主因之一[3]。高原鼢鼠采食也会影响草地植被群落结构, 对植被群落稳定性和物种多样性产生很大影响[4-5]。但是, 高原鼢鼠也是草地生物多样性的重要组成部分, 在草地生态系统食物网结构及其相应的能量流通和物质循环中有其独特的地位[6-7]。Grinnell等指出, 地下鼠类的灭绝将导致生活在该地区植物生命价值的贬值[8]。研究也表明, 鼢鼠造丘行为不仅会促进草地演替, 其挖掘和采食活动还会提高土壤通透性, 通过大量采食双子叶植物为家畜可食牧草生长提供有利条件[7, 9-11]。目前, 虽然有多项研究表明高原鼢鼠会造成草地生产力降低[12-13], 但这些研究大多是在有鼢鼠的草地和无鼢鼠的草地之间进行比较, 且指标较单一, 一般以草地产草量为主[14]。然而, 鼢鼠在草地生态系统中独特的生态位, 对草地植被乃至土壤的影响比较复杂[15]。无鼢鼠栖息的草地未必意味着草地质量较高, 不能简单的以单一标准去评价鼢鼠对草地的危害。鉴于对高原鼢鼠危害等级评价的分歧, 本研究设置不同高原鼢鼠种群密度梯度, 探讨其对草地产草量、植物多样性以及土壤的影响, 并探索适合于高寒草甸地区高原鼢鼠的取样面积, 为合理控制高原鼢鼠危害和保护草地生物多样性提供技术支撑。
1 研究地概括本试验在甘肃省天祝县抓喜秀龙乡的高寒草甸区进行, 该地区海拔2700—3100m, 地形受马牙雪山和雷公山的影响, 形成东向的峡谷地带, 西高东低, 地理坐标为N 37°11′, W 102°29′, 昼夜温差较大, 空气稀薄, 太阳辐射强, 气候寒冷潮湿, 年均降水量416 mm, 主要集中在7—9月, 年均蒸发量1592 mm, 无绝对无霜期。仅分冷、热两季。年均气温0.1 ℃, 7月和1月气温分别为12.7 ℃和-18.3 ℃, ≥0℃的年积温1380 ℃。植物生长期120—140 d。主要植物有鹅绒委陵菜(Potentilla anserina)、垂穗披碱草(Elymus nutans)、洽草(Koeleria cristata)、蒲公英(Taraxacum maurocarpam)、苔草(Carexscabri rostris)、平车前(Plantago depressa)、条叶龙胆(Gentiana manshurica)、矮嵩草(Kobresia humilis)、甘肃马先蒿(Pedicularis kansuensis)等,总计约34种[16]。
2 研究方法 2.1 实验设计实验区位于天祝县马营滩地区冬季草场(约7.5 hm2), 高原鼢鼠是该地区唯一造丘鼠类[17]。通过目测法选择3块有不同鼠丘密度的放牧草地作为样地。2014年4月开始, 在3个不同鼠丘密度区每月10—20日用地弓集中捕获鼢鼠, 连续捕获3个月捕尽为止。采用空间序列代替时间序列的方法, 对每个样地的鼠丘划分为3个类型:新鼠丘(当年形成)、半新鼠丘(1—2a内形成)和旧鼠丘(形成在3a以上)。在判断鼠丘类型时, 主要以鼠丘的土壤状况、植被盖度和植物种类组成为依据, 其中新鼠丘土壤疏松, 有龟裂, 基本无植被;半新鼠丘土壤较疏松, 植被盖度大于20%, 一年生植物约占90%以上;旧鼠丘土壤紧实, 植被盖度大于60%, 一年生植物比例小于10%。然后利用RTK(实时动态差分法)对3种类型的鼠丘进行标记, 以确定各类型鼠丘具体数量[18]。以鼠丘数和绝对鼠个体数代表高原鼢鼠种群密度[19], 将整个研究区划分为3个种群密度区(高密度区、中密度区和低密度区)(表 1)。在每个种群密度区根据样地面积设置3个重复, 单个重复面积为高密度区1 hm2, 中密度区为0.5 hm2, 低密度区为1 hm2。
| 样地类型 Sample type | 样地面积 Area of sample/hm2 | 捕获数量 Number of captured/ (hm2/只) | 旧鼠丘数 Number of old mounds/ (hm2/个) | 新鼠丘数 Number of new mounds/ (hm2/个) | 半新鼠丘数 Number of secondary new mounds/ (hm2/个) | 鼠丘总数 Number of total mounds/ (hm2/个) |
| 高密度区High density area | 3 | 28.33 | 1696.67 | 55.67 | 74.33 | 1826.67 |
| 中密度区Medium density area | 1.5 | 20.00 | 1041.33 | 33.33 | 117.33 | 1191.99 |
| 低密度区Low density area | 3 | 14.00 | 447.33 | 25.33 | 37.00 | 509.66 |
在所设置的重复样地内, 随机选择3种不同类型的土丘各5个, 并逐一编号, 将1/4 m2样方框置于鼠丘顶部, 将样方框内植物齐地刈割带回实验室烘干称其干重, 记录生物量。
2.2.2 鼠丘表面积计算为了监测各类型鼠丘面积占调查区面积比例和单位面积鼠丘产草量, 在计算鼠丘表面积时, 将鼠丘近似看作球缺。鼠丘高度(H)为土丘顶部到地面的垂直距离;鼠丘直径(R)为鼠丘的最长直径和最短直径的平均值。根据球缺面积公式可计算出鼠丘表面积公式:
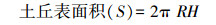
每个重复样地内非土丘地带, 按照“V”字型设置15个点, 每个样地共45个点。DWR是估测草地植物地上生物量和记录植物学组成的一种草地快速监测方法, 用0.1 m2正方形样方框估测设置的每个点产草量和草地植物学组成, 在估测完成后, 在同一样地中剪下一定数量0.1 m2样方的地上生物量, 经85 ℃、24 h烘干后做为实际产草量值, 用估测值与实际值做回归分析, 并用此回归方程作为后期估测的校准方程[20-21]。
统计植物学组成:在进行产草量估测的同时, 在同一样方内通过眼观样方中植物种类, 根据各植物种在样方内干物质重量的多少进行排名, 记录下前3种植物。在植物种记录过程中, 需注意以下情况, 若样方内只出现1种植物则将Rank1、Rank2、Rank3都记为该植物;若样方内出现2种植物A与B, 则根据干物质重比例分配两种植物;若样方内出现3种以上植物且有两种相等则在此后记录中一次记为A一次记为B。并且将同一样方中所出现前3种植物干物质百分含量赋值:Rank1为70.2%、Rank2为21.1%、Rank3为8.7%[22]。
2.2.4 土壤紧实度和水分测量在所设置的每个重复样地内按照“V”字型设置9个点, 每个样地共27个点。逐一编号以利于数据记录。在每个点上测量0—40 cm土壤紧实度, 并在此点周围50 cm内重复4次, 然后求平均值。用环刀法测量0—10、10—20、20—30 cm的土壤水分。
2.2.5 鼠丘取样面积调查在3个不同种群密度区, 每个区设置5个不同样方(100 m×100 m, 75 m×75 m, 50 m×50 m, 25 m×25 m和10 m×10 m), 调查每个样方内新鼠丘和半新鼠丘面积, 计算每个样方内新鼠丘和半新鼠丘面积占调查样方面积的比例。
2.3 统计分析所有数据采用Excel 2010、SPSS 19.0 for Windows软件进行One-Way ANOVA检验和相关性分析;BOTANAL进行植物学组成分析[23]。
3 结果与分析 3.1 鼠丘数量与绝对鼠密度的关系将3个调查区鼠丘数量和绝对捕尽数量进行相关性分析(图 1), 结果表明:各类型鼠丘数量和总鼠丘数量与绝对鼠密度呈显著正相关, 其中, 旧鼠丘数量与绝对鼠密度呈显著正相关(r=0.999, P < 0.05), 总鼠丘数量(r=0.995, P>0.05)和新鼠丘数量(r=0.983, P>0.05)与绝对鼠密度呈正相关, 半新鼠丘数量与绝对鼠密度相关性不显著(r=0.390, P>0.05)。说明3个调查区总鼠丘数量、旧鼠丘数量和新鼠丘数量可反映其种群密度, 可以用作表示高原鼢鼠种群密度的指标。

|
| 图 1 鼠丘数量与绝对鼠密度的关系 Fig. 1 Relationship between number of mounds and density of absolute zokors |
通过对不同密度区各类型鼠丘面积占样地面积百分比以及鼠丘产草量进行进行监测, 结果显示, 3个密度区中旧鼠丘(df=2, F=14.674, P < 0.05)和半新鼠丘(df=2, F=22.025, P < 0.05)面积占整个样地面积比例存在显著性差异, 而3个区新鼠丘面积占样地面积百分比无显著性差异(df=2, F=12.139, P>0.05)(图 2), 说明由高原鼢鼠不同种群密度当年内造成的次生裸地面积(新土丘面积)无差异。不同密度区中旧鼠丘和半新鼠丘产草量无显著性差异(df=2, F=2.649, P>0.05)(图 3), 说明种群密度的大小不会影响鼠丘产草量的变化。

|
| 图 2 不同类型鼠丘面积占样地面积百分比变化 Fig. 2 The percentage change of different types of mounds area of sample area |

|
| 图 3 不同种群密度鼠丘产草量的变化 Fig. 3 The changes of forage yield in sample area with different population density |
在3个高原鼢鼠种群密度区内, 对草地产草量和可食牧草产量进行One-Way ANOVA检验(图 4), 结果显示3个不同密度区非鼠丘草地产草量(df=2, F=2.909, P>0.05)和可食牧草产量(df=2, F=2.994, P>0.05)无显著性差异, 说明随着种群密度的升高, 地上生物量和可食牧草产量并没有显著降低, 高原鼢鼠不同种群密度对草地产草量的影响不大。

|
| 图 4 不同密度区地上生物量和可食牧草产量变化 Fig. 4 The changes of aboveground biomass and edible herbage yield in different density |
在植被盛长期(8月初), 3个不同密度区植物组成和所占比例的结果显示植物优势种相同(表 2), 依次为艾蒿(Artemisia argyi)、线叶嵩草(Kobresia capillifolia)和草地早熟禾(Poa pratensis);3个种群密度区可食牧草植物学组成和所占比例(高密度区为51%, 中密度区为51.7%, 低密度区为50%)基本相似;3个种群密度区单子叶植物学组成和所占比例基本相似, 双子叶植物分布有所不同(高密度区为13种, 中密度区为13种, 低密度区为15种), 但其所占比例都较小。总体来说, 3个不同种群密度区的植物组成和所占比例无较大差异。
| 物种名称 Plant species | 高密度区 High density | 中密度区 Medium density | 低密度区 Low density |
| 艾蒿Artemisia argyi Levl. et Vent. | 15.8 | 29.6 | 20.2 |
| 鹅绒委陵菜Potentilla anserine L. | 9.8* | 6.7* | 6.2* |
| 赖草 Aneurolepidium dasytachys (Trin) Nevski | 2.9 | 3.3 | 3.0 |
| 草地早熟禾Poa pratensis L. | 13.5 | 16.1 | 14.6 |
| 珠芽蓼Polygonum viviparum L. | 5.0* | 0.5* | 1.6* |
| 毛茛Ranunculus japonicus Thunb. | 1.0* | 0.2* | 1.6* |
| 线叶嵩草 Kobresia capillifolia(Decne.)C B.Clarke | 15.7 | 16.8 | 14.7 |
| 迷果芹Sphallerocarpus gracilis (Bess.) K. | 0.5* | 0.0* | 0.0* |
| 问荆Equisetum arvense L. | 4.7* | 0.9* | 0.0* |
| 扁蓿豆 Pocockia ruthrnia (L.) Boiss. | 3.5* | 0.0* | 0.7* |
| 阿尔泰狗哇花 Heteropappus altaicus (Willd.) Novopokr. | 0.2* | 0.0* | 0.0* |
| 小球花蒿Artemisia moorcroftiana Wall. | 6.5* | 2.0* | 6.8* |
| 矮嵩草Kobresia humilis (C.A.Mey.) Serg. | 3.7 | 3.8 | 2.4 |
| 垂穗披碱草Elymus nutans Griseb. | 12.4 | 11.7 | 13.6 |
| 乌头Aconitum carmichaeli Debx. | 1.6* | 0.0* | 3.8* |
| 二裂委陵菜Potentilla bifurca L. | 0.7* | 0.0* | 0.0* |
| 西伯利亚蓼Polygonum sibiricum Laxm. | 0.5* | 0.2* | 0.0* |
| 长叶火绒草Leontopodium longifolium Ling | 1.6* | 0.0* | 0.0* |
| 克氏针茅Stipa krylovii Roshev. | 0.0 | 0.0 | 0.5 |
| 高山唐松草 Thalictrum alpinum L. | 0.2* | 0.4* | 1.5* |
| 秦艽Gentiana macrophylla Pall. | 0.0* | 1.6* | 4.0* |
| 黄花棘豆Oxytropis ochrocephala Bunge | 0.0* | 2.6* | 2.4* |
| 葛缕子Carum carvi L. | 0.0* | 1.6* | 1.0* |
| 银莲花Anemone cathayensis Kitagawa | 0.0* | 0.0* | 0.4* |
| 甘肃马先蒿Pedicularis kansuensis Maxim. | 0.2* | 0.0* | 0.0* |
| 泛白委陵菜Potentilla discolor Bunge | 0.0* | 0.0* | 0.2* |
| 小米草Euphrasia regelii Wettst. | 0.0* | 1.6* | 0.5* |
| 兰石草Herb of Tibet Lancea, Tibet Lancea | 0.0* | 0.2* | 0.0* |
| 甘青老鹳草Geranium wilfordii Maxim | 0.0* | 0.2* | 0.0* |
| 多枝黄芪Astragalus polycladus Bur.et Franch. | 0.0* | 0.0* | 0.5* |
| *在植物学组成监测中出现的双子叶植物 | |||
3个不同高原鼢鼠种群密度区在土壤深度0—30 cm之间, 对土壤紧实度和水分统计分析表明(图 5, 图 6):在0—5 cm之间3个种群密度区土壤紧实度存在显著性差异(df=2, F=11.754, P < 0.05), 在5—10 cm之间高密度区土壤紧实度值显著小于中密度和低密度区(df=2, F=18.098, P < 0.05), 10—15 cm(df=2, F=8.822, P < 0.05)和15—20 cm(df=2, F=5.975, P < 0.05)之间高密度区和中密度区土壤紧实度显著小于低密度区, 20—25 cm(df=2, F=1.253, P>0.05)和25—30 cm(df=2, F=0.229, P>0.05)之间3个密度区土壤紧实度无显著性差异。说明高原鼢鼠采食深度范围(0—20 cm)内土壤紧实度随着种群密度的不同存在差异, 表现为种群密度越高土壤紧实度越小;其他土层紧实度随种群密度的增大无明显变化, 3个区0—10 cm(df=2, F=16.045, P < 0.05)和10—20 cm(df=2, F=20.948, P < 0.05)之间的水分存在显著性差异, 在20—30 cm之间高密度区土壤水分显著小于中密度区和低密度区(df=2, F=9.904, P < 0.05), 中密度区和低密度区土壤水分没有显著性差异(df=2, F=9.904, P>0.05)。说明高原鼢鼠种群密度与其栖息地土壤含水量有一定关系。

|
| 图 5 种群密度与土壤紧实度的关系 Fig. 5 Relationship between population density and soil compaction |

|
| 图 6 种群密度与土壤水分的关系 Fig. 6 Relationship between population density and soil moisture |
高原鼢鼠所推出的新鼠丘和半新鼠丘所占调查面积比例, 从产草量角度能够反映其对草地危害的大小。其中, 新鼠丘作为次生裸地, 是造成草地危害的主要指标。在不同调查面积下测量其面积所占调查区面积的比例(图 7), 结果显示, 随着调查面积的增加, 不同密度区新鼠丘和半新鼠丘面积占调查区面积比例随之变化, 当调查面积大于50 m×50 m时, 新鼠丘和半新鼠丘面积占调查区面积比例逐渐减小, 至75 m×75 m之后比例值趋于平缓。说明调查样地大小与危害评价有着密切关系, 调查高原鼢鼠危害的样方面积大小应该大于75 m×75 m(0.5 hm2)。

|
| 图 7 不同密度区新鼠丘面积占调查面积比例的变化 Fig. 7 The percentage changes of new moles area in survey area in different density |
地形、地貌特征、土壤特性、光照及降水量的变化是影响植被群落结构的非生物因素[24]。研究表明, 地形不仅影响着土壤的形成、土壤肥力的高低, 同时也在很大程度上决定着生物群落的构成[25];地貌特征也会对植被群落结构, 特别是对植被的恢复演替产生影响, 而土壤表面微地貌的变化对种子萌发也具有重要的影响[26]。这些不同特性的、自然发生的非生物因素对草地有着直接或间接影响, 从而改变植物群落的结构及草地的草产量[27]。鼠类在一定的种群数量之下, 并不像以前认为的对草地完全起到破坏作用, 反而适度的鼠密度有利于土壤水肥交替, 促进草地植物的生长, 扩大草场种子库等, 利于草地植被的恢复与优化[28]。本研究发现, 随着鼠丘数量(种群密度)的升高, 其草地产草量和植物学组成并没有出现差异, 这就印证了上述结果, 即在一定数量范围内的高原鼢鼠, 可能对草地质量不会造成影响。鼢鼠在采食过程中回避须根较多的单子叶植物草地, 选择2年生或者多年生双子叶生长地, 这就会为家畜可食的单子叶牧草生长创造有力条件[29]。此外, 草地产草量随高原鼢鼠种群密度增高没有出现差异的原因也可能跟鼢鼠造丘有关[9]。周建伟等研究发现形成在3a及以上的鼠丘, 其鼠丘上面的植被组成及产草量与草地原生植被无明显差异[17]。本研究结果也证实了随着鼠丘数量的增多, 由旧鼠丘隆起而造成整个生草地表面积就会增大, 造成草地产草量也随之增加。因此, 在适宜的种群密度下, 高种群密度区产草量与较低种群密度区产草量无显著差异。高原鼢鼠每年可推出大量新鼠丘, 即次生裸地, 这对于改善草地的通透性无疑具有不可低估的作用。张卫国等研究发现, 高原鼢鼠的造丘活动对草地的演替也有着积极作用[11], 不仅增大土壤的通透性, 也为草地植被生长创造了有力条件。由于温度和供氧等条件的变化有利于土壤微生物的活动, 致使大量富含有机质的土壤由地下转移至地表, 从而为以禾草为代表的喜肥植物的定植和扩张提供了极好的条件[30]。
一般来说, 高原鼢鼠对草地生态系统的扰动作用主要体现在2个方面, 一是由其采食降低产草量[31];二是由其挖掘活动导致草地产生大量异质性斑块, 引起植被群落结构的变化[32-33]。当草地中鼠丘数量增加时, 尤其是新鼠丘数量的增加, 尽管减少了生草地面积, 从而降低了产草量[9]。但是, 地下鼠挖掘形成的鼠丘能够增加栖息地的异质性, 允许具有不同竞争和入侵能力的物种共存, 从而影响了群落结构的格局和土壤理化性质[34-35]。本研究从植物学组成、优势种、土壤状况以证明群落稳定性一致, 生态功能正常发挥。根据高原鼢鼠危害防治标准可知, 当每公顷数量达到8只时, 即要进行防治[36], 在本文研究中, 低密度区高原鼢鼠种群密度已超过农业部规定的防治标准, 但是, 其草地产草量及可食牧草产量和优势种与中密度区和高密度区均无差异。在适度干扰的情况下, 高原鼢鼠通过其挖掘活动形成的大量鼠丘而成为高寒草甸异质性微生境的创造者, 增强了草地生态系统的稳定性, 是推动高寒草甸植被演替、维持其动态平衡的重要因素[30, 37]。当然, 因鼢鼠对退化草地的介入而引起草地植被变化的机制是一个极为复杂的生态过程, 其他因素的干扰也会使草地生态系统发生变化, 因此对于高原鼢鼠对草地造成的危害程度有必要进一步去界定。
由于各种啮齿动物的生态习性和栖息地环境不同, 其数量调查的方法和调查尺度亦各不相同[38]。在不同调查尺度下, 高原鼢鼠食草和挖掘活动对草地生态系统的影响有全然不同的结果[11]。目前农业部草原植保手册规定的样方为面积0.25 hm2[39]。但在本研究中, 分析3个不同种群密度区新鼠丘和半新鼠丘面积占草地面积比例, 发现调查样地大小与危害评价有着密切关系。当样方面积过小时, 新鼠丘和半新鼠丘面积会对调查结果造成很大影响。本研究发现在大于75 m×75 m的取样面积下, 新鼠丘和半新鼠丘面积趋于稳定, 就不会对草地产草量、可食牧草产量等调查造成误差。此外, 前人关于鼢鼠对草地危害评价的研究都是以产草量为指标, 有鼠丘与无鼠丘草地作为对照[7, 12]。但是, 动物选择栖息地是由多种因素决定的, 外部因子如气候、植被、土壤、地形、种间竞争、天敌等, 内部因子主要是种群密度所导致的种内竞争[40]。不能简单的以无鼠丘草地作为对照。因为, 无鼠丘草地有可能是鼢鼠的非栖息地而导致无对比性。在大尺度下调查高原鼢鼠对草地的危害有待深入研究。
| [1] | 周建伟, 花立民, 左松涛, 苏军虎, 王巧玲. 高原鼢鼠栖息地的选择. 草业科学 , 2013, 30 (4) : 647–653. |
| [2] | 段军红. 草地鼢鼠怀孕期的解剖研究. 青海畜牧兽医杂志 , 2011, 41 (2) : 31–32. |
| [3] | 余晓华, 刘荣堂. 高原鼢鼠的经济损害和经济阈值研究. 草原与草坪 , 2002, 98 (3) : 36–37. |
| [4] | 周俗. 草原无鼠害区建设技术设计. 草业科学 , 2006, 23 (5) : 82–86. |
| [5] | 张同作, 苏建平, 冯俊义, 雷晓水. 退耕还林还草地鼠类调查及控制对策研究. 草业科学 , 2005, 22 (4) : 83–87. |
| [6] | 钟文勤, 樊乃昌. 我国草地鼠害的发生原因及其生态治理对策. 生物学通报 , 2002, 37 (7) : 1–4. |
| [7] | 江小蕾, 张卫国, 杨振宇. 试论草原鼠类之定位. 草业科学 , 1997, 14 (5) : 34–35. |
| [8] | Grinnell J. The burrowing rodents of California as agents in soil formation. Journal of Mammalogy , 1923, 4 (3) : 137–149. DOI:10.2307/1373562 |
| [9] | 辛小娟, 杨莹博, 王刚, 任正炜, 储诚进, 张仁懿. 鼢鼠土丘植物群落演替生态位动态及草地质量指数. 生态学杂志 , 2011, 30 (4) : 700–706. |
| [10] | 杨莹博, 辛小娟, 艾得协措, 杨雪, 王绍美, 储诚进, 任正炜, 王刚. 鼢鼠土丘植被恢复演替过程中的物种多样性变化. 草业学报 , 2010, 19 (1) : 14–20. |
| [11] | 张卫国, 江小蕾, 王树茂, 杨振宇. 鼢鼠的造丘活动及不同休牧方式对草地植被生产力的影. 西北植物学报 , 2004, 24 (10) : 1882–1887. |
| [12] | 刘锦上, 张卫国, 江小雷, 卫万荣, 葛庆征. 高原鼢鼠洞道空间对高寒草甸植被性状的影响. 草地学报 , 2011, 19 (6) : 927–932. |
| [13] | 魏万红, 王权业, 周文扬, 樊乃昌. 灭鼠干扰后高原鼢鼠的种群动态与扩散. 兽类学报 , 1997, 17 (1) : 53–61. |
| [14] | 韩天虎, 张卫国, 丁连生. 高原鼢鼠栖息地的植被特征. 草业学报 , 1999, 8 (2) : 43–49. |
| [15] | 张灵菲, 张新中, 张燕堃, 冶娜娜, 傅华, 江小雷. 鼢鼠土丘演替对植物功能群与生产力的影响. 草业学报 , 2014, 23 (2) : 305–312. |
| [16] | 李凤霞, 张德罡. 草地退化指标及恢复措施. 草原与草坪 , 2005 (1) : 24–28. |
| [17] | 周建伟, 花立民, 王巧玲, 刘丽, 王贵珍. 天祝高寒草甸高原鼢鼠鼠丘植被演替调查. 草原与草坪 , 2014, 34 (3) : 8–13. |
| [18] | 蔡建忠, 张志勇. 实时动态定位技术在土地勘测工作中的应用. 测绘通报 , 2003 (5) : 32–34. |
| [19] | 宗浩, 樊乃昌, 于福溪, 朱嘉城. 高寒草甸生态系统优势鼠种高原鼢鼠(Myospalax baileyi)和高原鼠兔(Ochotona curzoniae)种群空间格局的研究. 生态学报 , 1991, 11 (2) : 125–129. |
| [20] | Forbis T A, Larmore J, Addis E. Temporal patterns in seedling establishment on pocket gopher disturbances. Oecologia , 2004, 38 (1) : 112–121. |
| [21] | Rodgers W E, Hartnett D C, Elder B. Effects of plants pocket gopher (Geomys bursarins) disturbances on tallgrass prairie plant community structure. The American Midland Naturalist , 2001, 145 : 344–357. DOI:10.1674/0003-0031(2001)145[0344:EOPPGG]2.0.CO;2 |
| [22] | Rezsutek M, Cameron G N. Vegetative edge effects and pocket gopher tunnels. Journal of Mammalogy , 2000, 81 (4) : 1062–1070. DOI:10.1644/1545-1542(2000)081<1062:VEEAPG>2.0.CO;2 |
| [23] | Mannetje L T, Haydock K P. The dry-weight-rank method for the botanical analysis of pasture. Grass and Forage Science , 1963, 18 (4) : 268–275. DOI:10.1111/gfs.1963.18.issue-4 |
| [24] | Pihlgren A. Vascular Plants and Small-Scale Spatial Heterogenity in Semi-Natural Grasslands. Uppsala:Swedish University of Agricultural Sciences, 2005. |
| [25] | Smith M, Capelle J. Effects of soil surface microtopography and litter cover on germination, growth and biomass production of chicory (Cichorium intybus L.). The American Midland Naturalist , 1992, 128 (2) : 246–253. DOI:10.2307/2426458 |
| [26] | Hobbs R J, Mooney H A. Effects of rainfall variability and gopher disturbance on serpentine annual grassland dynamics. Ecology , 1991, 72 (1) : 59–68. DOI:10.2307/1938902 |
| [27] | 张堰铭, 刘季科. 高原鼢鼠挖掘对植物生物量的效应及其反应格局. 兽类学报 , 2002, 22 (4) : 292–298. |
| [28] | Huntly N, Inouye R. Pocket gophers in ecosystems:patterns and mechanisms. BioScience , 1988, 38 (11) : 786–793. DOI:10.2307/1310788 |
| [29] | 刘仁华, 刘炳友, 赵秀成, 柳劲松, 王俊森. 林区鼢鼠鼠害的主要特征及其生态控制对策. 兽类学报 , 1997, 17 (4) : 272–278. |
| [30] | Zhang Y M, Zhang Z B, Liu J K. Burrowing rodents as ecosystem engineers:the ecology and management of plateau zokors Myospalax fontanierii in alpine meadow ecosystems on the Tibetan Plateau. Mammal Review , 2003, 33 (3) : 284–294. |
| [31] | 张堰铭. 高原鼢鼠对高寒草甸群落特征及演替的影响. 动物学研究 , 1999, 20 (6) : 435–440. |
| [32] | 王刚, 杜国祯. 鼢鼠土丘植被演替过程中的种的生态位分析. 生态学杂志 , 1990, 9 (1) : 1–6. |
| [33] | 宗文杰, 江小雷, 严林. 高原鼢鼠的干扰对高寒草地植物群落物种多样性的影响. 草业科学 , 2006, 23 (10) : 68–72. |
| [34] | Horn H S, Mac Arthur R H. Competition among fugitive species in a harlequin environment. Ecology , 1972, 53 (4) : 749–752. DOI:10.2307/1934797 |
| [35] | Denslow J S. Disturbance-mediated coexistence of species//Pickett S T A, White P S, eds. The Ecology of Natural Disturbance and Patch Dynamics. Orlando, Fla:Academic Press, 1985:307-323. |
| [36] | 农业部. 草原鼠荒地治理技术. 中国牧业通讯 , 2008 (13) : 40–44. |
| [37] | 张军, 葛庆征, 张卫国, 江小雷. 植被性状与高原鼢鼠栖息地适合度的关系. 草业科学 , 2011, 28 (5) : 836–840. |
| [38] | 孔照芳. 草原鼠虫病害研究. 兰州: 甘肃民族出版社, 1996100–102. |
| [39] | 贠旭疆, 农业部畜牧业司, 全国畜牧总站. 草原植保实用技术手册. 北京: 中国农业出版社, 201037–39. |
| [40] | Laundré J W. Effect of ground squirrel burrows on plant productivity in a cool desert environment. Journal of Range Management , 1998, 51 (6) : 638–643. DOI:10.2307/4003605 |