 2016, Vol. 36
2016, Vol. 36文章信息
- 付贵全, 徐先英, 徐梦莎, 孟源源, 赵鹏, 刘江
- FU Guiquan, XU Xianying, XU Mengsha, MENG Yuanyuan, ZHAO Peng, LIU Jiang.
- 民勤绿洲边缘2种生境红砂种群分形特征及影响因素
- A study of fractal properties of spatial distribution of Reaumuria soongorica population and its influencing factors in Minqin Oasis Fringe
- 生态学报[J]. 2016, 36(18): 5768-5776
- Acta Ecologica Sinica[J]. 2016, 36(18): 5768-5776
- http://dx.doi.org/10.5846/stxb201504120741
-
文章历史
- 收稿日期: 2015-04-12
- 网络出版日期: 2016-01-05





2. 甘肃省治沙研究所, 荒漠化与风沙灾害防治国家重点实验室培育基地, 兰州 730070;
3. 中国林业科学研究院, 国家林业局泡桐研究开发中心, 郑州 450003;
4. 浙江农林大学, 环境与资源学院, 临安 311300
2. Gansu Desert Control Research Institute, State Key Laboratory Breeding Base of Desertification and Aeolian Sand Disaster Combating, Lanzhou 730070, China;
3. Chinese Academy of Forestry, China Paulownia Research and Development Center, Zhengzhou 450003, China;
4. Zhejiang Agricultural and Forestry University, College of Environmental and Resource Sciences, Lin'an 311300, China
种群格局是种群生物学特性、种间种内关系和环境综合作用的结果[1-2]。以前在研究格局分析的过程中由于受到研究方法的限制, 常常会出现理论拟合与其在解释生物学意义上相互矛盾的情况[3], 之后有一些方法对此做出了改进, 但在解决尺度依赖问题上仍显不足[4]。分形模型是目前国内外应用相当广泛的非线性模型, 它能够刻画个体随尺度变化的本质特征[2, 5-6], 该理论由Loehle首次引入生态学领域[7], 随后在树木的分枝格局、植冠格局、种群格局、群落格局和景观格局[8-13]等方面取得了丰硕的成果。但是面对实际问题, 不能够局限于空间格局的研究, 还应该把所处的环境特征联系起来, 这样更有利于我们弄清楚现象的本质。
红砂(Reaumuria soongorica)又称枇杷柴, 是广泛分布于我国干旱荒漠区的多年生、耐旱、耐碱、耐贫瘠和较强集沙能力的C3小灌木, 其具有很强的抗逆性和生态可塑性, 是草原化荒漠和典型荒漠地区的重要建群种和优势种之一[14-15]。然而由于生态环境的变化以及人为活动的干扰, 红砂的生存环境和发展受到了极大的影响, 近年来众多学者对红砂做了比较多的研究, 主要集中在:水、盐胁迫下等其它方面的生理生态[16-24]、种子的萌发和繁殖特性[25-30]、生物学特征[14-15, 31-32]等, 但关于红砂种群空间格局分析的研究报道不多, 尤其是在这两种生境下的研究更少, 而且在以往对种群格局的研究过程中只选择某一个或两个分形维数参数进行研究, 有一定的局限性, 另外该地区梭梭不同程度退化但红砂却能够定居其中。鉴于此, 本文在分析红砂种群结构特征的基础上, 通过综合运用分形维数中的计盒维数、信息维数和关联维数分别对两种生境下红砂种群空间格局进行研究, 并对其影响因素做了主成分分析, 目的在于揭示分形维数在红砂种群格局研究中的生态学意义以及不同生境下红砂种群对环境的适应机理和途径, 以期为干旱区绿洲边缘生态修复和红砂种群的保护利用与培育提供理论支撑。
1 研究区概况及研究方法 1.1 研究区概况研究区位于石羊河下游的甘肃民勤荒漠草地生态系统国家野外科学观测研究站(38°34′1″N, 102°58′19″E), 气候属温带大陆性荒漠气候, 年均气温7.4 ℃, 年均降水量115.2 mm, 多集中于7—9月份, 年平均蒸发量2419.6 mm, 是降水量的24倍, 多年平均风速2.35 m/s, 多年平均日照时数为2832.1 h;土壤为风沙土;地貌以粘土沙障+梭梭林固定沙丘和丘间底地为主, 固定沙丘出现了不同程度的活化, 人工梭梭林衰退;植被由梭梭(Haloxylon ammodendron)、白刺(Nitraria tangutorum)、红砂、芦苇(Phragmites australis)、沙蓬(Agriophyllum squarrosum)、沙拐枣(Calligonum arborescens)等组成。
1.2 研究方法 1.2.1 样地设置2014年8月和2015年6月在民勤西沙窝选择丘间低地和梭梭林固定沙丘两种生境下有代表性样地2处, 然后在每个样地内选取3个面积为10 m×10 m的样方。丘间低地生境(38°35′1”N, 102°59′19”E)下群丛盖度为21.24%, 北面为线型流动沙丘, 南面、西面为粘土+梭梭固沙林, 东面是农田, 地表以物理结皮为主, 结皮盖度在50—75%左右, 主要伴生物种有画眉草(Eragrostis pilosa)、盐生草(Halogeton glomeratus)、猪毛菜(Salsola collina)和白刺;固定沙丘生境(38°34′27″N, 102°59′7″E)下群丛盖度35.95%, 粘土+梭梭固沙林坡地, 地表以生物结皮为主, 结皮盖度15%—30%左右, 主要伴生物种有画眉草、盐生草、猪毛菜、沙蓬、黄花矶松( Limonium aureum)和虎尾草(Chloris virgata)。样地内设置邻接格子样方, 每个格子样方1 m×1 m, 共计600个格子样方, 测定每株红砂的空间二维坐标、冠幅、株高等指标, 并分别在样地内每隔10 cm采集0—40 cm土样, 每个样方取3—5个重复, 测定土壤含水量、全盐和有机质等理化性质。
1.2.2 龄级的确定本文采取空间序列代替时间序列的方法, 也就是大小结构分析法, 以植株的冠幅与株高之积(即植株体积)的立方根和其立方根的分布情况将红砂种群分为3个龄级, 即发育的3个不同阶段, Ⅰ级d < 11 cm;Ⅱ级11 cm≤d < 31 cm;Ⅲ级d ≥31 cm。
1.2.3 分形分析(1) 计盒维数
将样方逐次进行格栅化, 统计给定网格边长值下相对应的非空格子数(Ne), 将每次划分所统计的非空格子数与相对应的网格边长在双对数坐标下进行(分段)直线拟合, 其斜率的绝对值即为计盒维数。本文将样方边长由2等分(对应的网格边长为5 m)划分到100等分(对应的网格边长为0.1 m), 划分99次:
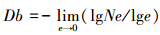
式中, e为划分的网格边长, Ne为网格边长为e时的非空格子数。
(2) 信息维数
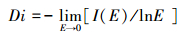
具体原理:首先对样方进行逐步的网格覆盖, 分别统计每一个非空格子中的个体数(Ni)和样方内个体总数(N), 得出某一非空格子的概率(Pi=Ni/N)和信息量(Ii=-PilnPi), 则在该网格边长的信息总量为I(E)=∑Ii;然后通过I(E)与相对应的网格边长E在双对数坐标下进行(分段)拟合直线, 其斜率的绝对值即为信息维数。网格边长的划分和计盒维数相同。
(3) 关联维数

步骤如下:①逐一计算样方内每一个体与其它个体之间的欧氏距离;②给定一系列的距离值, 分别统计小于该系列相对应距离值的欧氏距离个数Mi;③由给定距离值所对应的欧氏距离个数Mi与该系列距离总数M的比值求得C(i);④通过C(i)与相对应的距离值在双对数坐标下进行(分段)拟合直线, 其斜率的绝对值即为关联维数。根据本文的实际研究情况, 距离值的变化从0 m到15 m, 步长为0.1 m。
1.2.4 数据分析样地环境因子的主成分分析以及均值间差异的t检验在Excel 2013和PASW Statistics 18.0中进行;分形维数的分析根据原理, 通过自编程序在MATLAB中进行统计分析;并采用SigmaPlot 12.5作图。
2 结果与分析 2.1 红砂种群的结构特征不同生境条件下红砂种群的龄级结构特征, 反映了红砂种群对环境的适应以及生境条件对红砂种群更新的影响。由表 1得知, 丘间低地生境下红砂种群个体主要集中在Ⅰ级, 占总数的84.8%, 大部分集中在龄级Ⅰ级和Ⅱ级, 共占总数的96.3%, 表明红砂种群更新状况良好;而固定沙丘生境下红砂种群主要集中在Ⅲ级, 占总数的54.4%, Ⅱ级和Ⅲ级共占总数的91.6%, 表明红砂种群的更新受到抑制;两种生境下红砂株高的极小值和极大值、株数以及不同龄级个数相比差异显著(P≤0.05)或极显著(P≤0.01), 且丘间低地生境下红砂种群的密度(9.4株/m2)是固定沙丘生境下红砂种群的密度(2.2株/m2)的4.3倍;但固定沙丘下红砂株高均值为31.3 cm, 变异系数为55.9%, 丘间低地5.3 cm, 变异系数为109.4%, 相比差异显著(P≤0.05)或极显著(P≤0.01)。
| 生境Habit | 株数Number | 极小值Minimum | 极大值Maximum | 均值Mean | 标准偏差Standard deviation | 龄级数Number of age class | ||
| Ⅰ | Ⅱ | Ⅲ | ||||||
| 丘间低地Interdunal depression | 940a** | 0.5a** | 50.0a** | 5.3a** | 5. 8 | 798a** | 107a** | 35a** |
| 固定沙丘Fixed dunes | 215b | 5.0b | 75.0b | 31.3b | 17.5 | 18b | 80b | 117b |
| 字母不相同则表明差异显著(P≤0.05), **表明差异极显著(P≤0.01) | ||||||||
由图 1可以看出, 拟合直线分为两个线性区域, 而只有拐点以后的尺度范围才有研究意义, 因此可以得知在丘间低地生境下红砂种群的计盒维数分别为1.8977(A1)、1.9010(A2)和1.9294(A3), 均值为1.9094;梭梭林固定沙丘生境下红砂种群的计盒维数分别为1.7905(B1)、1.6685(B2)和1.7605(B3), 均值为1.7398。表明梭梭林固定沙丘生境下红砂种群具有较高的生态空间占据程度和生态空间利用能力, 而丘间低地生境下红砂种群具有极强的生态空间占据程度和生态空间利用能力。

|
| 图 1 两种生境下红砂种群的计盒维数 Fig. 1 Box-counting dimension of Reaumuria soongorica population in the two habitats |
由图 2可知, 在分析过程中与计盒维数一样出现了拐点, 分为两个不同的线性区域, 和计盒维数一样只有拐点以后的尺度才具有研究意义, 因此得知丘间低地生境下红砂种群的信息维数分别为1.7329(A1)、1.7645(A2)和1.7293(A3), 均值为1.7422;梭梭林固定沙丘生境下红砂种群的信息维数分别为1.6313(B1)、1.7031(B2)和1.6820(B3), 均值为1.6721。表明与梭梭林固定沙丘生境下相比, 丘间低地生境下红砂种群格局强度较大, 系统结构较复杂。

|
| 图 2 两种生境下红砂种群的信息维数 Fig. 2 Information dimension of Reaumuria soongorica population in the two habitats |
由图 3可见, 在直线拟合过程中出现了两个不同的线性区域, 表明种群内的个体在拐点前后出现了不同的空间自相似性, 即在小尺度(拐点以前)上空间相关性较高, 大尺度(拐点以后)上空间相关性较小;在丘间低地生境下, 红砂种群的关联维数分别为1.4743(A1)、1.5036(A2)和1.4587(A3), 均值为1.4789;在固定沙丘生境下, 红砂种群的关联维数分别为1.7475(B1)、1.8075(B2)和1.7287(B3), 均值为1.7612。表明与丘间低地生境相比, 固定沙丘生境下红砂种群的关联维数较大, 种群个体间竞争较强。

|
| 图 3 两种生境下红砂种群的关联维数 Fig. 3 Correlation dimension of Reaumuria soongorica population in the two habitats |
种群格局的分形特征不但与种群的自身结构特征有关, 而且还与所处的环境因子密切相关。在对环境因子进行主成分分析的过程中, 首先对生物结皮和物理结皮进行量化并分别赋值2和1。研究结果表明, 3个主成分的特征值分别为4.1、1.7和1.4, 第一主成分是土壤的结皮类型, 贡献率为45.9%, 第二主成分是土壤含水量, 贡献率为18.5%, 第三主成分为土壤全盐含量, 贡献率为15.6%, 这3个主成分累计贡献率为80.002%。
3 讨论 3.1 红砂种群格局的分形特征在植物群落中的每个种群生长期间, 都需要有一定的空间生态资源, 但由于受到生境质量、群落地位、竞争和适应能力以及种群发育程度差异的影响[33], 其利用和占据生态空间的能力以及格局强度亦有差别, 这种差别分别可以通过计盒维数和信息维数来量化。计盒维数是用来表征种群占据和利用生态空间能力的量度, 但是当计盒维数小于1时, 一般不予以考虑, 因为此时表征的是个体特征, 而只有计盒维数大于1时才具有生态意义, 可以依此来解释种群的空间分布特征[11], 信息维数是计盒维数的进一步深化和延伸, 与计盒维数相比, 它还考虑到非空格子的信息量大小, 因而表征了系统结构复杂程度随尺度的变化规律, 即揭示出种群格局强度的尺度变化特征[34]。从图 1和2的统计结果来看, 丘间低地生境下比固定沙丘生境下的红砂种群具有更强的生态空间占据程度和生态空间利用能力以及较复杂的空间格局强度, 这主要是由于丘间低地生境下红砂个体的数量较多聚集分布明显, 密度较大是固定沙丘的4.3倍;而且丘间低地生境下红砂幼苗很多, 占总株数的84.8%, 种群更新良好, 但固定沙丘生境下红砂幼苗只占总株数的8.4%(表 1), 种群更新受到抑制。这与宋宇洋[3]、向悟生[35]和郝朝运[33]等对梭梭种群、红豆杉(Taxus chinensis)种群和永瓣藤(Monimopetalum chinense)种群的研究结果相一致, 进一步表明了计盒维数和信息维数对种群的个体数量、更新状况和分布情况反映灵敏, 即计盒维数和信息维数愈大, 表明其种群个体数量愈多、更新愈好、分布更加聚集, 反之则种群个体数量少、更新差、分布更加随机。
关联维数的生态意义在于刻画出种群空间关联性随尺度的变化规律, 关联维数愈大(接近2), 表明种群内个体的空间关联程度愈高, 个体间的竞争愈加强烈;反之, 关联维数越小(远离2), 表明种群内个体的空间相关程度越低, 个体间的竞争越弱[12]。从图 3的分析结果来看, 固定沙丘生境下红砂种群的关联维数比丘间低地生境下的要大, 与丘间低地生境相比, 固定沙丘生境下的水土资源相对贫瘠, 加之风沙流的危害, 个体在此生境下对资源竞争更加强烈。研究发现[12, 36], 在适宜的生境下, 关联维数越大表明种群个体越聚集, 竞争越强烈, 种群整体对生境的空间占据和利用能力愈强, 但本文的研究结果与此不尽相同, 却与宋宇洋[3]等对梭梭种群的研究结果一致, 这是因为与适宜的环境条件相比, 在恶劣的干旱荒漠地区, 生境差, 这进一步说明了关联维数在恶劣的生境下有其特定的生态意义, 即只表示种群竞争的强弱, 需要在以后的研究过程中进一步关注。
| 环境因子Habit factor | 主成份Component | ||
| 1 | 2 | 3 | |
| 特征值Eigenvalue | 4.129 | 1.665 | 1.407 |
| 贡献率Proportion/% | 45.873 | 18.498 | 15.631 |
| 累计贡献率Cumulative proportion/% | 45.873 | 64.371 | 80.002 |
| 全氮Total nitrogen/(g/kg) | 0.746 | 0.579 | 0.099 |
| 地下水位Underground water level/m | -0.637 | -0.099 | 0.511 |
| PH | 0.802 | -0.393 | 0.259 |
| 全盐Total salt/(g/kg) | 0.108 | -0.108 | 0.912 |
| 有机质Organic matter/(g/kg) | -0.801 | 0.178 | 0.002 |
| 容重Bulk density/(g/cm3) | 0.583 | 0.097 | -0.403 |
| 含水量Water content/% | -0.239 | 0.949 | 0.177 |
| 全磷Total phosphorus/(g/kg) | 0.729 | 0.427 | 0.187 |
| 结皮类型Crust type | -0.970 | 0.172 | -0.091 |
综上所述, 根据红砂种群的分形维数特征, 建议在防风固沙和植被恢复时, 丘间低地生境下可以通过增大红砂Ⅰ小龄级个体的密度来提高整体的防风固沙能力和幼苗的成活量, 但在梭梭林固定沙丘生境下如何提高红砂Ⅰ小龄级个体的成活率是保护和利用红砂的关键。
3.2 红砂种群格局分形特征的影响因素植物种群空间格局的分形维数不但取决于种群结构(密度、幼体数量等), 还与种群的生境因子有紧密的联系。陈育[37]、王国严[38]和李海东[39]、杨洪晓[40]、曹宴宁[41]以及宋于洋[3]等分别对不同生境下半日花(Helianthemum songaricum)、砂生槐(Sophora moorcroftiana)、油蒿(Artemisia ordosica)、差不嘎蒿(Artemisia halodendron )以及梭梭种群的空间格局及关联性进行研究, 结果表明生境对种群格局的影响很大, 在不同的生境下植物种群通过随机或非随机的分布方式以及不同龄级间的正(负)关联性来适应不同生境下水土等条件的差异, 并以此来指导干旱沙区防风固沙林的建设。本文研究结果表明两种生境下空间格局的分形维数有一定的差异, 这种差异是由于生境的异质性以及环境因子引起的。主成分分析表明, 土壤结皮类型是红砂种群格局分形维数的第一主成分, 土壤含水量是第二主成分, 土壤全盐含量是第三主成分, 这3个主成分的累计贡献率为80.002%。
丘间低地多为物理结皮且比较薄, 地表结皮在降水后强烈的蒸发导致地表多形成龟裂空隙, 这些空隙为红砂种子保存和萌发创造便利条件;但固定沙丘表面形成大量的生物结皮, 阻碍水分的下渗[42-43]和种子[44]的保存, 不利于红砂种子的着床和萌发, 这就导致两种生境下红砂种群的密度和幼体数量产生较大的差异, 其计盒维数和信息维数的差异就是具体的表现形式。
在干旱荒漠地区, 由于降水稀少、蒸发强烈, 导致植物大多数情况下处于水分亏缺和高盐含量的胁迫下, 但诸多学者的研究表明红砂可以通过多方位自身调节来最大限度地获取和维持水分以及抵抗Na+的毒害, 从而更好地适应生存环境, 有关学者研究发现红砂可以通过调节超氧化物歧化酶和过氧化物酶的活性、叶绿素的含量、游离脯氨酸的含量以及硝酸还原酶的含量来抵抗不同生境下干旱缺水的环境[24]并且其幼苗在不同生境下可以通过优化根系空间的分布构型来调节和利用水分[31-32];另外研究还发现红砂能够将体内的Na+排除体外来减轻Na+的毒害, 从而维持植株的水分[20]并且红砂种子在萌发的过程中可能进化出了干旱胁迫与盐胁迫交叉适应的策略[25]。
4 结论基于对民勤绿洲边缘两种生境下有代表性红砂种群样地的野外调查, 采用空间替代时间和分形理论探讨了两种生境下的红砂种群结构特征和空间格局的分形特征, 并对其影响因素做了主成分分析, 研究结果:
(1) 红砂在丘间低地生境下主要集中在龄级Ⅰ, 种群个体数量多、密度大、更新较好;在固定沙丘生境下, 主要集中在龄级Ⅲ, 种群个体数量少、密度小、更新受到限制。
(2) 丘间低地生境下比固定沙丘生境下的红砂种群具有更强的生态空间占据程度和生态空间利用能力以及较复杂的空间格局强度, 这进一步说明计盒维数和信息维数对种群的个体数量、更新状况和分布情况反映灵敏;但固定沙丘生境下红砂种群的关联维数比丘间低地生境下的要大, 表明种群在固定沙丘生境下竞争更强, 和生境较好地区种群格局的关联维数相比, 进一步证明关联维数在干旱地区有其特定的生态学意义, 即只表示种群竞争的强弱。
(3) 红砂种群的分形特征与生境的异质性以及环境因子和群落结构有关。主成分分析表明, 土壤结皮类型是红砂种群格局分形维数的第一主成分, 土壤含水量和土壤全盐含量分别是第二主成分和第三主成分, 这3个主成分的累计贡献率为80.002%。
(4) 在进行防风固沙和植被恢复时, 丘间低地下可以通过增大小龄级个体的密度来提高整体的防风固沙能力和幼苗的成活量, 但在贫瘠、恶劣的退化梭梭林固定沙丘提高小龄级个体的成活率是保护和利用红砂的关键。
致谢: 感谢甘肃省治沙研究所常丽博士对写作的帮助。| [1] | 惠刚盈, GadowK V, 胡艳波, 陈伯望. 林木分布格局类型的角尺度均值分析方法. 生态学报 , 2004, 24 (6) : 1225–1229. |
| [2] | 刘小恺, 刘茂松, 黄峥, 徐驰, 张明娟, 王汉杰. 宁夏沙湖4种干旱区群落中主要植物种间关系的格局分析. 植物生态学报 , 2009, 33 (2) : 320–330. |
| [3] | 宋于洋, 李园园, 张文辉. 基于Ripley的K(r)函数和分形维数的梭梭种群空间格局. 应用生态学报 , 2010, 21 (4) : 827–835. |
| [4] | 马克明, 祖元刚. 植被格局的分形特征. 植物生态学报 , 2000, 24 (1) : 111–117. |
| [5] | Burrough P A. Fractal dimensions of landscapes and other environmental data. Nature , 1981, 294 (5838) : 240–242. DOI:10.1038/294240a0 |
| [6] | Palmer M W. Fractal geometry:a tool for describing spatial patterns of plant communities. Vegetation , 1988, 75 (1/2) : 91–102. |
| [7] | Loehle C, Li B L, Sundell R C. Forest spread and phase transitions at forest-prairie ecotones in Kansas, U.S.A.. Landscape Ecology , 1996, 11 (4) : 225–235. DOI:10.1007/BF02071813 |
| [8] | Berezovskaya F S, Karev G P, Kisliuk O S, Khlebopros R G, Tsel'niker Y L. A fractal approach to computer-analytical modelling of tree crowns. Trees , 1997, 11 (6) : 323–327. DOI:10.1007/s004680050091 |
| [9] | Zeide B. Fractal geometry in forestry applications. Forest Ecology Natural Management , 1991, 46 (3-4) : 179–188. DOI:10.1016/0378-1127(91)90230-S |
| [10] | Johnson A R, Milne B T, Wiens J A. Diffusion in fractal landscapes:simulations and experimental studies of tenebrionid beetle movements. Ecology , 1992, 73 (6) : 1968–1983. DOI:10.2307/1941448 |
| [11] | 马克明, 祖元刚. 兴安落叶松种群格局的分形特征:计盒维数. 植物研究 , 2000, 20 (1) : 104–111. |
| [12] | 马克明, 祖元刚, 倪红伟. 兴安落叶松种群格局的分形特征——关联维数. 生态学报 , 1999, 19 (3) : 353–358. |
| [13] | 张立杰, 赵文智, 何志斌. 青海云杉(Picea crassifolia)种群格局的分形特征及其影响因素. 生态学报 , 2008, 28 (4) : 1383–1389. |
| [14] | 刘家琼, 邱明新, 蒲锦春, 鲁作民. 我国荒漠典型超旱生植物——红砂. 植物学报 , 1982, 24 (5) : 485–488. |
| [15] | 马茂华, 孔令韶. 新疆呼图壁绿洲外缘的琵琶柴生物生态学特性研究. 植物生态学报 , 1998, 22 (3) : 237–244. |
| [16] | 赵昕, 杨小菊, 石勇, 何明珠, 谭会娟, 李新荣. 盐胁迫下荒漠共生植物红砂与珍珠的根茎叶中离子吸收与分配特征. 生态学报 , 2014, 34 (4) : 963–972. |
| [17] | 谭会娟, 李新荣, 刘玉冰, 赵昕. 盐胁迫下红砂愈伤组织的抗氧化能力与耐盐性研究. 中国沙漠 , 2013, 33 (2) : 549–553. |
| [18] | 高茜, 李毅, 苏世平, ЖигуновA B, 单立山, 种培芳. 盐胁迫对红砂(Reaumuria soongorica)种子吸胀过程中生理特性的影响. 中国沙漠 , 2014, 34 (1) : 83–87. |
| [19] | 耿东梅, 单立山, 李毅, ЖигуновA B. 土壤水分胁迫对红砂幼苗叶绿素荧光和抗氧化酶活性的影响. 植物学报 , 2014, 49 (3) : 282–291. |
| [20] | 周航宇, 包爱科, 杜宝强, 王锁民. 荒漠植物红砂响应高浓度NaCl的生理机制. 草业科学 , 2012, 29 (1) : 71–75. |
| [21] | 肖生春, 肖洪浪, 宋耀选, 段争虎, 陆明峰. 荒漠植被红砂(Reaumurta soongorica)水热响应的年轮学研究. 中国沙漠 , 2006, 26 (4) : 548–552. |
| [22] | 李骁, 王迎春, 征荣. 西鄂尔多斯地区强旱生小灌木水分参数的研究(I). 中国沙漠 , 2005, 25 (4) : 581–586. |
| [23] | 刘美玲, 曹波, 刘玉冰, 谭会娟, 陈永乐, 张亚峰. 红砂(Reaumuria soongorica)黄酮类物质代谢及其抗氧化活性对UV-B辐射的响应. 中国沙漠 , 2014, 34 (2) : 426–432. |
| [24] | 周紫鹃, 苏培玺, 解婷婷, 张海娜, 李善家. 不同生境下红砂(Reaumuria soongorica)的生理生化特征及适应性. 中国沙漠 , 2014, 34 (4) : 1007–1014. |
| [25] | 宋雪梅, 杨九艳, 吕美婷, 杨明, 张作如. 红砂种子萌发对盐胁迫及适度干旱的响应. 中国沙漠 , 2012, 32 (6) : 1674–1680. |
| [26] | 王万鹏, 朱恭, 李正平. 红砂育苗的土壤水分管理初步研究. 中国沙漠 , 2006, 26 (4) : 596–599. |
| [27] | 朱恭, 王万鹏, 李正平. 超旱生小灌木——红砂大田育苗技术研发. 中国沙漠 , 2006, 26 (4) : 612–618. |
| [28] | 曾彦军, 王彦荣, 张宝林, 庄光辉. 红砂种群繁殖特性的研究. 草业学报 , 2002, 11 (2) : 66–71. |
| [29] | 曾彦军, 王彦荣, 保平, 塔拉腾, 苏勒德. 几种生态因子对红砂和霸王种子萌发与幼苗生长的影响. 草业学报 , 2005, 14 (5) : 24–31. |
| [30] | 苏世平, 李毅, 种培芳. 河西走廊不同红砂天然群体种子活性相关性. 生态学报 , 2012, 32 (1) : 135–141. |
| [31] | 单立山, 李毅, 段雅楠, 耿东梅, 李真银, 张荣, 段桂芳, ЖигуновА В. 红砂幼苗根系形态特征和水分利用效率对土壤水分变化的响应. 西北植物学报 , 2014, 34 (6) : 1198–1205. |
| [32] | 单立山, 李毅, 董秋莲, 耿东梅. 红砂根系构型对干旱的生态适应. 中国沙漠 , 2012, 32 (5) : 1283–1290. |
| [33] | 郝朝运, 张小平, 李文良, 张昱. 不同类型群落中濒危植物永瓣藤(Monimopetalum chinense)种群的空间分布格局. 生态学报 , 2008, 28 (6) : 2900–2908. |
| [34] | 马克明, 祖元刚. 兴安落叶松种群格局的分形特征——信息维数. 生态学报 , 2000, 20 (2) : 187–192. |
| [35] | 向悟生, 李先琨, 苏宗明, 欧祖兰, 陆树华. 元宝山南方红豆杉克隆种群分布格局的分形特征. 植物生态学报 , 2007, 31 (4) : 568–575. |
| [36] | 吴承祯, 洪滔, 宋萍, 范海兰, 游爱华. 长苞铁杉种群空间格局的分形特征. 福建林学院学报 , 2008, 28 (1) : 1–5. |
| [37] | 陈育, 杨劼, 张璞进, 清华, 赵利清, 张雷. 西鄂尔多斯半日花(Helianthemum soongoricum)种群结构和点格局分析. 中国沙漠 , 2014, 34 (1) : 75–82. |
| [38] | 王国严, 方江平, 许新勇, 尼玛曲珍, 钟蕊, 赵旷. 雅鲁藏布江中游干旱沙地砂生槐种群结构与点格局分析. 中国沙漠 , 2010, 30 (5) : 1092–1098. |
| [39] | 李海东, 沈渭寿, 佘光辉, 孙明, 袁磊, 林乃峰. 西藏砂生槐种群结构与点格局分析. 中国沙漠 , 2011, 31 (6) : 1443–1448. |
| [40] | 杨洪晓, 张金屯, 吴波, 李晓松, 张友炎. 毛乌素沙地油蒿种群点格局分析. 植物生态学报 , 2006, 30 (4) : 563–570. |
| [41] | 曹晏宁, 史利莎, 韩烁, 侯安燕, 涂凡, 张金屯. 科尔沁沙地植被恢复中差不嘎蒿种群不同龄级个体的点格局分析. 植物学通报 , 2008, 25 (4) : 437–442. |
| [42] | 李新荣, 龙利群, 王新平, 张景光, 贾玉奎. 干旱半干旱地区土壤微生物结皮的生态学意义及若干研究进展. 中国沙漠 , 2001, 21 (1) : 4–11. |
| [43] | 李守中, 肖洪浪, 宋耀选, 李金贵, 刘立超. 腾格里沙漠人工固沙植被区生物土壤结皮对降水的拦截作用. 中国沙漠 , 2002, 22 (6) : 612–616. |
| [44] | 陈荣毅, 魏文寿, 张元明, 吴楠, 张静. 干旱区生物土壤结皮对种子植物多样性的影响. 中国沙漠 , 2008, 28 (5) : 868–873. |