 2016, Vol. 36
2016, Vol. 36文章信息
- 郑学博, 樊剑波, 崔键, 徐磊, 祝振球, 周静, 何园球
- ZHENG Xuebo, FAN Jianbo, CUI Jian, XU Lei, ZHU Zhenqiu, ZHOU Jing, HE Yuanqiu.
- 沼液还田对旱地红壤微生物群落代谢与多样性的影响
- Analysis on metabolic characteristics and functional diversity of soil edaphon communities in upland red soil under biogas slurry application
- 生态学报[J]. 2016, 36(18): 5865-5875
- Acta Ecologica Sinica[J]. 2016, 36(18): 5865-5875
- http://dx.doi.org/10.5846/stxb201503230555
-
文章历史
- 收稿日期: 2015-03-23
- 网络出版日期: 2015-12-29





2. 中国科学院大学, 北京 100049;
3. 国家红壤改良工程技术研究中心, 中国科学院红壤生态实验站, 鹰潭 335211;
4. 江西省科学院生物资源研究所, 南昌 330029
2. Universityof Chinese Academy of Sciences, Beijing 100049, China;
3. National Engineering Research and Technology Center for Red Soil Improvement, Red Soil Ecological Experiment Station, Chinese Academy of Sciences, Yingtan 335211, China;
4. Institute of Biology Resource, Jiangxi Academy of Sciences, Nanchang 330029, China
土壤生物是土壤生态系统的核心, 而土壤微生物是土壤生物的重要组成部分, 在土壤结构形成、有机物质分解与转化、营养元素循环与利用、作物生长和生态环境改善、植物病害抑制或拮抗、温室气体排放等方面起着重要的作用[1-4], 是土壤肥力的重要调控因子和土壤地力的关键驱动因子之一[5]。可敏感反映土壤质量变化、表征土壤有机质代谢旺盛程度, 是评价土壤质量的主要指标之一。而其功能多样性信息对于明确不同环境中微生物群落的作用具有重要意义[6]。
Biolog法是描述微生物群落功能变化的重要指标, 其应用始于Garland和Mills[7]。以ECO板碳源利用为基础, 是描述微生物群落功能多样性的一种简单、快速的方法[8-9], 其反应特征能代表实际土壤微生物群落对ECO板底物碳源利用的动力学特征[10-11]。广泛应用于评价不同土壤的微生物群落功能多样性:不同退耕年限[5];不同灌溉水矿化度和施氮量[1];不同有机氯农药污染[12];不同肥料类型[13]等;可以发现肥料类型、土壤类型等因素不同, 微生物群落结构多样性不同。
沼液是畜禽粪便、作物秸秆、工业垃圾、城市垃圾、能源植物植物等厌氧发酵产生的终端产物之一, 含有大量环境有害元素(CODCr, BOD5等), 无序排放可导致水体严重富营养化, 正在成为危害性和危险性不断增大的一种污染源, 因此其如何处理已经成为当前水环境保护中的关键问题[14]。另一方面, 沼液中有机质、大量元素(N、P、K)、微量元素(B、Mn等)等含量丰富[15], 且存在形式多为易被植物吸收利用的速效养分, 是一种缓速兼备的液体有机肥[14]。因此, 沼液被认为是一种良好的土壤改良剂或有机肥料[16]。前人关于沼液还田的研究主要集中在作物产量、肥料利用效率、土壤C, N矿化、微生物数量等, 而且主要以单施沼液为主。Galvez A等研究发现合理的沼液农用可以改善土壤质量, 平衡土壤中各营养元素[17];Abubaker J等研究发现沼液农用提高了小麦产量、土壤有机氮矿化和氨氮氧化潜力[18];同时, 沼液农用提高了土壤微生物活性[19];温晓霞等的研究表明沼液还田可以提高蔬菜、水果产量[20];冯伟等发现沼液还田可以提高小麦根际土壤微生物数量和酶活性[21]。但关于沼液还田, 尤其是沼液化肥配施对土壤微生物群落功能多样性影响的研究却鲜见报道。
因此, 本文采用Biolog技术研究不同沼液还田量下0—20 cm土壤微生物群落功能多样性变化, 旨在探讨土壤微生物功能多样性对不同沼液还田量的响应、定量沼液还田对土壤微生物指标和土壤质量的影响, 从而为旱地红壤质量改善、畜禽废弃物资源化利用及土壤资源的可持续利用提供理论依据和科学指导。
1 材料与方法 1.1 试验区概况试验区位于江西省余江县刘家站鲁王村(116°5′E, 28°12′N)。区内年平均降雨量1750 mm, 80%的降雨集中在3—6月, 年蒸发量1350 mm, 年平均气温18℃, 无霜期267 d左右, 属亚热带湿润季风气候。
试验田为旱平地, 试验时间2013年4月—2014年8月, 前茬作物为花生。试验设置6个处理, 每个处理3次重复, 各小区面积12.4 m×4.8 m=60 m2, 小区间田埂宽50 cm, 高25 cm, 保护行20 cm。供试土壤为第四纪红粘土发育的典型红壤。0—20cm土壤基本理化性质见表 1。
| 土壤类型 Type of soil | 有机质 Organic matter/ (g/kg) | 全氮 Total nitrogen/(g/kg) | 碱解氮 Alkali-hydrolyzale nitrogen/(mg/kg) | 速效磷 Available phosphorus/ (mg/kg) | 速效钾 Available potassium/(mg/kg) | pH (H2O) |
| 红壤Red soil | 12.15 | 0.83 | 35.54 | 15.41 | 169.21 | 4.94 |
各处理N-P2O5-K2O量均为120-90-135 kg/hm2(CK除外)。具体为:Ⅰ对照(不施肥, CK);Ⅱ单施化学氮磷钾肥(NPK);Ⅲ化学氮磷钾肥+沼液(沼液全N占总N 15%, BS15);Ⅳ化学氮磷钾肥+沼液(沼液全N占总N 30%, BS30);Ⅴ化学氮磷钾肥+沼液(沼液全N占总N 45%, BS45);Ⅵ单施沼液(BS100)。具体施肥量见表 2。
氮肥为尿素(N 46%), 磷肥为钙镁磷肥(P2O5 12%), 钾肥为氯化钾(K2O 60%), 沼液发酵原材料主要是猪粪尿等, 发酵3个月以上, 细腻均匀, 透明无明显粪臭味, pH 7.67, 具体养分含量见表 3。
| 指标Indexes | 含量Contents/(mg/L) |
| 有机碳Organic carbon | 162 |
| NH4+-N Ammonium nitrogen | 269 |
| 总磷Total phosphorus | 54 |
| 总氮Total nitrogen | 281 |
| NO3--N Nitrate nitrogen | 0.32 |
| 总钾Total potassium | 188 |
所有肥料全部基施, 沼液还田处理中不足的P2O5、K2O量用化肥补足(表 2)。2013年播种前整地规划小区、翻耕, 播种前2d浇灌沼液, 尿素、钙镁磷肥、氯化钾播种前1d二次翻耕翻入。种植制度为单季花生/a, 每年4月中旬播种, 8月中旬收获。试种品种赣花1号, 密度14万株/hm2, 作物生长期间不再进行追肥。
| 处理 Treatment |
花生Peanut/(kg/hm2) | ||
| Na | P2O5 | K2O | |
| CK | 0/0 | 0/0 | 0/0 |
| BS15 | 18/102 | 6/84 | 27/108 |
| BS45 | 54/66 | 17/73 | 82/53 |
| NPK | 0/120 | 0/90 | 0/135 |
| BS30 | 36/84 | 12/78 | 55/80 |
| BS100 | 120/0 | 37.5/52.5 | 180/0 |
| a:N-P2O5-K2O分别来自沼液/化肥;CK:不施肥No fertilizer; NPK:单施化学氮磷钾肥Chemical fertilizers only;BS15:化学氮磷钾肥+沼液(沼液全N占总N 15%) 85% chemical fertilizer total nitrogen plus 15% biogas slurry total nitrogen; BS30:化学氮磷钾肥+沼液(沼液全N占总N 30%) 70% chemical fertilizer total nitrogen plus 30% biogas slurry total nitrogen; BS45:化学氮磷钾肥+沼液(沼液全N占总N 45%) 55% chemical fertilizer total nitrogen plus 45% biogas slurry total nitrogen; BS100:单施沼液Biogas slurry only | |||
取样时间为2014年8月中旬花生收获前2d。
取样方法采用S形取样, 各小区选取5个样点, 用土钻(直径5.0cm)采集0—20 cm土层土样, 先混匀然后去除其中的杂物、砂砾及作物细根等。过2mm筛后用于测定土壤微生物功能多样性(4℃保存)。
1.4 试验方法 1.4.1 土壤微生物功能多样性分析采用Biolog-ECO微平板法对不同沼液还田量处理土壤微生物群落的功能多样性进行测定。每个ECO板含31种碳源, 分碳水化合物(10种)、羧酸(7种)、氨基酸(6种)、聚合物(4种)、酚类(2种)、胺类(2种)6种类型[22]。试验步骤:称取1g鲜土, 加入到盛有9mL 0.85% NaCl无菌溶液的18mL试管中, 封口后在微型振动仪震荡5min, 按10倍稀释法用0.85% NaCl无菌溶液将其稀释到原来的10-4用于接种。将加样液倾倒在无菌加样槽中, 用8通道加样器将每孔加样150uL, 每样1板, 3次重复, 将接种好的ECO板(EcoPlatesTM, 美国Matrix Technologies Corporation生产)置于25℃暗箱连续培养, 期间每隔24h用Biolog自动鉴定系统(MicroStationTMSystem, 美国Biolog公司)在590nm下读数, 直到光密度值稳定为止, 总共培养时间240h。
1.4.2 计算方法土壤微生物利用碳源的整体能力用平均吸光度值(AWCD)表示:
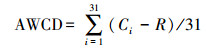 (1)
(1)
式中, Ci为第i孔的吸光度值;R为对照孔的吸光度值;Ci-R < 0时记为0。
土壤微生物利用ECO板中六类碳源的能力用各自的平均吸光度值(AWCD′)[23]表示:
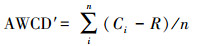 (2)
(2)
式中, Ci为i孔的吸光度值;R为对照孔的吸光度值;n为所选择碳源类型的个数;Ci-R < 0时记为0。
土壤微生物群落功能多样性分析:本研究采用培养120h的AWCD和AWCD′值进行多样性指数计算、主成分分析和聚类分析。土壤微生物群落功能多样性指数包括Shannon指数(H)、丰富度指数(S)、Shannon均匀度指数(E)、Simpson优势度指数(D)。具体计算公式如下[24-25]:
Shannon指数
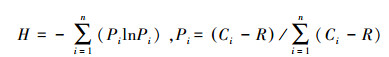 (3)
(3)
丰富度指数S:被利用碳源的总数目(判断标准为Ci-R>0.25);
Shannon均匀度指数
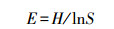 (4)
(4)
Simpson优势度指数
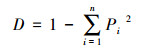 (5)
(5)
式中, Ci为第i孔的吸光度值;R为对照孔的吸光度值;n为ECO板碳源数量, n=31;Pi为第i孔的相对吸光度值与整个ECO板相对吸光度值总和的比率;S为颜色变化的碳源的数量。
1.5 数据处理试验所得数据采用Excel 2007进行处理、绘制图表, SPSS 13.0统计分析软件进行单因素方差分析(One-way ANOVA)、相关性分析(大小采用pearson指数)、主成分分析和聚类分析, 差异显著性分析采用Duncan新复极差方法, 显著性水平P < 0.05。
2 结果与分析 2.1 土壤微生物利用碳源动力学分析 2.1.1 不同沼液还田量下土壤微生物群落整体代谢活性变化(AWCD)平均吸光值AWCD反映了土壤微生物群落利用碳源的整体能力, 是反映微生物代谢活性的有效指标[7, 26]。图 1所示, 不同沼液还田量处理土壤微生物群落代谢AWCD值的变化趋势均为随着时间的延长而升高。0—48h:各处理AWCD值均较低, 说明土壤微生物群落代谢活性较低, 碳源基本未被利用;48—168h:各处理AWCD值迅速升高, 微生物呈指数增长, 说明土壤微生物从48h后开始大量利用碳源, 土壤微生物代谢活性显著升高。168h后:各处理AWCD值基本趋于平缓, 此时土壤微生物活性达到稳定。总体上, 不同沼液还田处理下土壤微生物利用31种碳源的综合能力(AWCD)为:BS30>BS45> NPK > CK >BS100> BS15。BS30和BS45处理120h的微生物活性显著高于BS15处理。

|
| 图 1 土壤微生物群落平均吸光值的变化 Fig. 1 Changes of AWCD of soil edaphon against incubation time under biogas slurry application |
按照化学基团性质, 31种碳源分碳水化合物(10种)、羧酸(7种)、氨基酸(6种)、聚合物(4种)、酚类(2种)、胺类(2种)等共六大类碳源。分析各类碳源的AWCD值可判断各处理土壤微生物群落对特定类型碳源的利用能力。本文选取120h时的AWCD值绘制不同沼液还田量下土壤微生物碳源利用能力图(图 2)。各处理之间土壤微生物群落代谢碳水化合物、羧酸、聚合物的能力存在显著差异(P < 0.05), 代谢氨基酸、酚类、胺类的能力无显著差异(P < 0.05)。其中, 碳水化合物类:BS45>BS30> NPK≈CK >BS100>BS15;羧酸类:BS30> NPK >BS100>BS45> CK >BS15;氨基酸类:BS45>BS30> CK > NPK >BS100>BS15;聚合物类:BS45>BS30 ≈ NPK >BS100> CK >BS15;酚类:BS45>BS30> CK > NPK ≈ BS100>BS15;胺类:BS45>BS30> CK >BS15> NPK >BS100。可以发现, 土壤微生物碳水化合物、氨基酸、酚类AWCD值与上述31种碳源总体利用情况类似。BS30处理土壤微生物对6类碳源的代谢能力高于除BS45处理以外的各处理, 而BS45处理较其它处理则提高了土壤微生物对除羧酸类以外的五类碳源的代谢能力。

|
| 图 2 不同沼液还田量下120h时土壤微生物不同碳源AWCD Fig. 2 AWCD of soil edaphonat 120h of different carbon sources under biogas slurry application |
Shannon物种指数、Shannon均匀度指数、Simpson优势度指数和碳源利用丰富度指数可以反映土壤微生物群落的功能多样性。不同的多样性指数反映土壤微生物群落功能多样性的不同方面, Shannon指数反映群落物种及其个体数;Shannon均匀度指数反映群落个体分布的均匀制度;Simpon优势度指数反映群落中最常见物种的优势度[27]。各处理ECO板培养120h后的土壤微生物群落功能多样性指数见表 2。丰富度指数、Shannon指数、Simpson优势度指数方面均为BS45处理最高, BS30处理次之, BS15处理最低。而Shannon均匀度指数方面则为BS15处理最高, 显著高于其它处理, 而剩余处理间无显著差异。可见, BS30和BS45处理增加了土壤中微生物的种类和优势种群的数量, 但并没有提高微生物种类的平均分布;而BS15处理抑制了土壤中微生物的种类和优势种群的数量, 但提高了土壤中存在的微生物种类的均匀分布。
2.3 不同沼液还田量下土壤微生物群落功能主成分分析(PCA)土壤微生物群落功能多样性反映了群落总体的变化, 但未能反映微生物群落代谢的详细信息, 研究土壤微生物对不同碳源利用能力的差异, 利于更全面地研究微生物群落代谢功能的特性。以培养120h不同处理微生物群落对ECO板31种碳源利用情况做主成分分析(PCA), 31个成分因子中前8个主成分(累积方差贡献率>85%, 特征根>1)的累积方差贡献率达到92.70%。从中提取可以聚集单一碳源变量的数据变异的第1主成分(PC1, 方差贡献率32.34%, 特征根10.02)和第2主成分(PC2, 方差贡献率15.36%, 特征根4.76)来分析土壤微生物群落功能多样性。第3至第8主成分的方差贡献率较小, 分别为12.76%、10.85%、7.93%、5.88%、4.16%、3.43%。
不同沼液还田量下土壤微生物群落碳源利用的PCA排序图如下(图 3)。由图 3可知, 前两个主成分共解释了总变异的47.70%。结合方差分析结果表明, 各处理在PC1上存在明显的空间分异, 6种处理的土壤微生物群落可明显分为4簇:BS30、BS45处理聚为一簇, CK、NPK处理聚为一簇、BS100处理为一簇、BS15处理为一簇。而在PC2上则没有显著的空间差异。PC1方差分析表明, 来自(BS30、BS45)和BS15处理的土壤微生物群落结构存在显著差异。同时, 在PC1上, 各处理土壤微生物群落碳源利用主成分分析得分为BS30>BS45> NPK > CK >BS100>BS15。这与Shannon指数、丰富度指数、Simpson指数的研究结果一致。

|
| 图 3 不同沼液还田量下土壤微生物群落功能主成分分析 Fig. 3 The principal component analysis for soil microbial community function under biogas slurry application |
原始变量与各主成分的相关系数可用载荷值表征, 载荷值越大, 表明该原始变量是该主成分的主要影响因子, 对该主成分的贡献越大[28]。表 5为31种碳源(原始变量)在前2个主成分上的载荷值。从表中可以看到, 对PC1贡献大的碳源(载荷值≥0.50或≤-0.50)有16种, 决定了主成分1的变异, 其中碳水化合物4种(β-甲基-D-葡萄糖苷, D-甘露醇, 葡萄糖-1-磷酸盐, D, L-a-甘油), 羧酸2种(D-葡萄胺酸, 衣康酸), 氨基酸4种(L-精氨酸, L-天冬酰胺酸, L-苯基丙氨酸, L-苏氨酸), 聚合物3种(吐温40, 吐温80, 肝糖), 酚类2种(2-羟基苯甲酸, 4-羟基苯甲酸), 胺类1种(腐胺);对PC2贡献大的碳源(载荷值≥0.50或≤-0.50)有8种, 决定了主成分2的变异, 其中碳水化合物2种(D-木糖, N-乙酰基-D-葡萄胺, ), 羧酸4种(D-葡萄胺酸, D-半乳糖醛酸, y-羟基丁酸, D-苹果酸), 氨基酸1种(甘氨酰-L-谷氨酸), 聚合物1种(a\\环式糊精)。结合图 3, 发现较其他处理, BS30和BS45显著提高了碳水化合物(4种)、羧酸(2种)、氨基酸(4种)、聚合物(3种)、酚类(2种)、胺类(1种)等共16种碳源的代谢能力。
| 处理 Treatment |
多样性指数Diversity indexes | |||
| 丰富度指数S Diversity index S |
Shannon物种指数H Shannon species index H |
Shannon均匀度指数E Shannon evenness index E |
Simpson优势度指数D Simpon dominance index D |
|
| CK | 13.5±2.1ab | 2.66±0.23ab | 1.02±0.03b | 0.92±0.01ab |
| NPK | 14.5±3.5a | 2.86±0.09ab | 1.08±0.07ab | 0.93±0.01a |
| BS15 | 6.5±2.1b | 2.58±0.05b | 1.42±0.29a | 0.90±0.00b |
| BS30 | 16.5±0.7a | 2.89±0.05ab | 1.03±0.03b | 0.94±0.00a |
| BS45 | 17.0±4.2a | 2.95±0.13a | 1.05±0.05b | 0.94±0.01a |
| BS100 | 12.5±2.1ab | 2.70±0.05ab | 1.07±0.05ab | 0.92±0.00ab |
| 同列不同小写字母表示处理间差异显著(P < 0.05) | ||||
| 碳源类别 Carbon sources | 底物 Substrates | 第一主成分 The primary principal component (PC1) | 第二主成分 The secondary principal component (PC2) | 第三主成分 The tertiary principal component (PC3) |
| 碳水化合物 | D-纤维二糖 | 0.442 | 0.147 | -0.164 |
| Carbohydrates | a-D-乳糖 | 0.353 | -0.083 | 0.669 |
| β-甲基-D-葡萄糖苷 | 0.553 | 0.121 | -0.208 | |
| D-木糖 | -0.067 | 0.750 | 0.343 | |
| I-赤藻糖醇 | -0.013 | -0.155 | 0.457 | |
| D-甘露醇 | 0.553 | 0.121 | -0.208 | |
| N-乙酰基-D-葡萄胺 | -0.067 | 0.750 | 0.343 | |
| 葡萄糖-1-磷酸盐 | 0.900 | 0.119 | -0.271 | |
| D, L-a-甘油 | 0.611 | 0.458 | -0.260 | |
| D-半乳糖内酯 | -0.082 | -0.413 | 0.068 | |
| 羧酸类Carboxylic acids | 丙酮酸甲酯 | 0.160 | 0.473 | 0.527 |
| D-葡萄胺酸 | 0.552 | -0.501 | 0.142 | |
| D-半乳糖醛酸 | 0.140 | -0.802 | 0.283 | |
| y-羟基丁酸 | -0.013 | 0.864 | -0.044 | |
| 衣康酸 | 0.442 | -0.151 | 0.664 | |
| a-丁酮酸 | 0.517 | 0.054 | -0.205 | |
| D-苹果酸 | 0.003 | -0.538 | -0.006 | |
| 氨基酸类Amino acids | L-精氨酸 | 0.910 | -0.136 | 0.056 |
| L-天冬酰胺酸 | 0.925 | -0.171 | -0.153 | |
| L-苯基丙氨酸 | 0.682 | -0.190 | -0.326 | |
| L-丝氨酸 | 0.134 | 0.065 | 0.602 | |
| L-苏氨酸 | 0.899 | -0.007 | -0.066 | |
| 甘氨酰-L-谷氨酸 | 0.385 | 0.777 | 0.325 | |
| 聚合物类Polymers | 吐温40 | 0.608 | 0.037 | 0.605 |
| 吐温80 | 0.799 | -0.182 | -0.353 | |
| a-环式糊精 | 0.330 | 0.720 | -0.043 | |
| 肝糖 | 0.500 | -0.269 | -0.301 | |
| 酚类Phenols | 2-羟基苯甲酸 | 0.691 | 0.093 | -0.444 |
| 4-羟基苯甲酸 | 0.558 | -0.404 | 0.463 | |
| 胺类Amines | 苯乙基胺 | 0.372 | -0.148 | 0.335 |
| 腐胺 | 0.835 | 0.052 | -0.422 |
对所提取的8个主成分分别计算各处理6类碳源的主成分得分(Pricipal component scores, F), 结果如图 4。F值的相对大小可以表征不同处理土壤微生物群落对同一类碳源的代谢能力和同一处理土壤微生物对不同类型碳源代谢能力的高低。对F值进行横向比较发现, 不同处理土壤微生物对不同碳源的利用能力不同。CK:代谢碳水化合物能力最强;NPK:代谢碳水化合物、羧酸、聚合物能力最强;BS30、BS45、BS100:代谢碳水化合物、羧酸、氨基酸、聚合物能力较强;BS15:代谢碳水化合物和羧酸能力较强;各处理土壤微生物群落均为代谢酚类和胺类能力较弱。对F值进行纵向比较可知, 代谢碳水化合物、羧酸、氨基酸、聚合物、酚类和胺类能力最弱的都是来自BS15处理的土壤微生物, 代谢以上物质最强的都是来自BS30和BS45处理的土壤微生物。

|
| 图 4 不同沼液还田量处理碳源类型主成分得分 Fig. 4 Pricipal component scores of carbon sources under biogas slurryapplication |
同时对所提取的8个主成分分别计算各处理6类碳源的主成分综合得分(Pricipal component composite scores, ZF) (图 5)。ZF值的相对大小可以反映不同处理土壤微生物对ECO板31种碳源综合代谢能力的高低。图 5显示, 6种处理土壤微生物对ECO板中31种单一碳源的综合代谢能力排序为:BS45>BS30> NPK >CK >BS100>BS15。

|
| 图 5 不同沼液还田量处理碳源类型主成分综合得分 Fig. 5 Pricipal component composite scores of carbon sources underbiogas slurryapplication |
以各处理的前2个主成分得分代替原指标作为评价土壤微生物代谢能力的新指标, 以欧式距离作为衡量各处理土壤微生物代谢能力的差异, 采用最短距离法将各处理按土壤微生物代谢能力的亲疏相似程度进行系统聚类。由聚类树形图(图 6)可直观地看出各处理土壤微生物代谢能力的归类情况(以欧式距离10为分异点):BS30, BS45> CK, NPK >BS100>BS30, 即各处理微生物群落代谢能力分为4个等级:一等为BS30, BS45, 属强土壤微生物群落代谢能力等级;二等为CK, NPK, 属较强土壤微生物群落代谢能力等级;三等为BS100, 土壤微生物群落代谢能力低;四等为BS15, 土壤微生物群落代谢能力最低。这与主成分分析结果一致。

|
| 图 6 聚类树形图 Fig. 6 Arborescence of cluster analysis |
研究表明, 微生物群落碳源代谢强度和代谢类型的变化能较早预测土壤养分的变化趋势, 被认为是预测土壤质量变化最有潜力的敏感性生物指标[29-30]。土壤微生物是土壤生态系统中最重要和最活跃的部分, 是驱动土壤有机物质和养分转化与循环的主要动力, 其活性和多样性变化在很大程度上反映了土壤有机碳库的转化情况[31]。而其数量、群落结构及活性受外源有机物投入种类、存在状态和数量的影响[32]。有机物料作为外源有机碳源进入土壤, 改变了土壤微生物的生存环境和微生物所需能源物质的数量和质量[33], 从而对土壤自身有机碳库的转化产生影响。刘定辉等[34]发现秸秆还田循环利用显著增加了土壤微生物数量和生物量。刘国顺等[35]、李正等[36]发现化肥与绿肥配施还田后土壤微生物活性显著提高, 利于有机物质的转化和土壤质量的提高。AWCD从功能代谢水平上揭示土壤微生物群落结构多样性, 是反映土壤微生物活性, 描述土壤微生物群落功能多样性的重要指标[30]。本文研究表明, 较不施肥处理, 单施化肥和单施沼液处理土壤微生物群落碳源代谢强度无显著变化, 这与程存刚等[32]的研究结果一致;而沼液化肥配施处理提高了土壤微生物群落碳源代谢强度, 特别是沼液全N配施45%和30%处理土壤微生物群落碳源代谢强度显著提高, 碳水化合物4种(β-甲基-D-葡萄糖苷, D-甘露醇, 葡萄糖-1-磷酸盐, D, L-a-甘油), 羧酸2种(D-葡萄胺酸, 衣康酸), 氨基酸4种(L-精氨酸, L-天冬酰胺酸, L-苯基丙氨酸, L-苏氨酸), 聚合物3种(吐温40, 吐温80, 肝糖), 酚类2种(2-羟基苯甲酸, 4-羟基苯甲酸), 胺类1种(腐胺)等16种碳源代谢能力增幅最大。说明一定比例沼液与化肥配施可以改善土壤微生物群落的营养供应, 进而提高微生物群落代谢能力。原因可能是该比例沼液化肥配施为土壤微生物生长提供所需碳源和能源的同时, 使微生物C/N适宜, 土壤速效养分含量多, 促进了花生植株的吸收和根系生长, 从而增加了根系分泌物, 促进了土壤中微生物数量的增长和活性的提高。另一方面, 单施化肥会导致红壤进一步酸化[23], 而土壤中绝大多数微生物适宜在中性环境中生存繁殖, 沼液的强pH(表 2)抑制了这种酸化趋势。不同沼液还田量下, 土壤微域环境发生了改变, 这使得多种微生物的适宜性受到影响, 从而影响了土壤微生物对碳源的选择性利用。
不同的多样性指数反映土壤微生物群落功能多样性的不同方面。丰富度指数S、Shannon均匀度指数、Simpson优势度指数分别从微生物群落物种丰富度、均一性及常见物种优势度等方面反映土壤微生物群落多样性[27]。本研究发现BS45处理Shannon指数、丰富度指数S、Simpson优势度指数较其它处理分别提高2.07%—37%, 2.94%—61.76%和1.08%—4.26%。说明该处理在增加土壤微生物种群物种种类和数量的基础上提高了优势微生物种群的数量, 提高了土壤微生物群落的多样性。
对各试验处理AWCD值进行进一步主成分和聚类分析(图 6), 结果表明各试验处理土壤微生物群落多样性分为4组, BS30、BS45处理微生物群落多样性相似,为一组;不施肥、单施化肥处理为一组;BS100处理为一组;BS15处理为一组。具体为BS45,BS30> CK,NPK >BS100>BS15。这说明沼液还田量显著影响土壤微生物群落多样性, 而对土壤微生物群落结构变化的影响则需要通过PCR-DGGE方法进一步分析。本试验仅针对连续施沼液两年后的土壤微生物群落活性及多样性进行研究, 而关于长期沼液农用对土壤微生物群落活性和多样性的影响以及是否对土壤质量带来负面影响等方面有待进一步研究。
4 结论(1) 与不施肥、单施化肥处理相比, 单施沼液处理对土壤微生物群落代谢活性影响较小, 而沼液化肥配施处理显著影响土壤微生物群落代谢活性。沼液全N配施比例在30%—45%时, 土壤微生物群落整体活性最高, 而且碳源利用类型以碳水化合物(4种)、羧酸(2种)、氨基酸(4种)、聚合物(3种)、酚类(2种)和胺类(1种)等共16种碳源为主;
(2) BS45处理Shannon物种指数、丰富度指数、Simpson优势度指数较其它处理分别提高2.07%—37%, 2.94%—61.76%和1.08%—4.26%, 该处理在增加土壤微生物种群物种种类和数量的基础上提高了优势微生物的数量, 同时提高了土壤微生物群落功能多样性。
(3) 结合主成分分析(PCA)和聚类分析得出:微生物群落多样性分为4个等级:一等为BS30, BS45处理, 属强土壤微生物群落代谢能力等级;二等为CK, NPK处理, 属较强土壤微生物群落代谢能力等级;三等为BS100处理, 土壤微生物群落代谢能力低;四等为BS15处理, 土壤微生物群落代谢能力最低。
综上, 对于旱地红壤而言, 沼液并不能完全取代化肥单独施用, 必须与一定比例的化肥配合施用。该研究中, 沼液化肥氮配比为30%—45%是最佳配施比例, 利于土壤质量的保持和提高。
| [1] | 闵伟, 侯振安, 冶军, 马丽娟, 曹志, 罗慧兰. 长期咸水滴灌棉田土壤微生物活性及群落功能多样性. 生态学杂志 , 2014, 33 (11) : 2950–2958. |
| [2] | 郑亚萍, 吴正峰, 王才斌, 梁晓艳, 陈殿绪, 孙学武, 王建国, 郑永美, 冯昊, 孙秀山. 旱地花生不同土壤类型主要土壤肥力指标季节变异及其相互关系研究. 核农学报 , 2013, 27 (6) : 831–838. |
| [3] | 林先贵, 胡君利. 土壤微生物多样性的科学内涵及其生态服务功能. 土壤学报 , 2008, 45 (5) : 892–900. |
| [4] | Shi W, Miller B E, Stark J M, Norton J M. Microbial nitrogen transformations in responce to treated dairy waste in agricultural soils. Soil Science Society of America Journal , 2004, 68 (6) : 1867–1874. DOI:10.2136/sssaj2004.1867 |
| [5] | 董立国, 蒋齐, 蔡进军, 张源润, 许浩, 李生宝. 基于Biolog-ECO技术不同退耕年限苜蓿地土壤微生物功能多样性分析. 干旱区研究 , 2011, 28 (4) : 630–637. |
| [6] | Preston M J, Boddy L, Randerson P F. Analysis of microbial community functional diversity using sole-carbon-source utilization profiles acritique. FEMS Microbial Ecology , 2002, 42 (1) : 1–14. |
| [7] | Garland J L, Mills A L. Classification and characterization of heterotrophic microbial communities on basis of patterns of community-lever sole-carbon-source utilization. Applied and Environental Microbiology , 1991, 57 : 2351–2359. |
| [8] | Garland J L. Analysis and interpretation of community-level physiological profiles in microbial ecology. FEMS Microbiology Ecology , 1997, 24 (4) : 289–300. DOI:10.1111/j.1574-6941.1997.tb00446.x |
| [9] | Hollibaugh J T. Relationship between thymidine metabolism, bacterioplankton community metabolic capabilities and sources of organic matter. Microbial Ecology , 1994, 28 (2) : 117–131. DOI:10.1007/BF00166800 |
| [10] | 章家恩, 蔡燕飞, 高爱霞, 朱丽霞. 土壤微生物多样性试验研究方法概述. 土壤 , 2004, 36 (4) : 346–350. |
| [11] | 陈承利, 廖敏, 曾路生. 污染土壤微生物群落结构多样性及功能多样性测定方法. 生态学报 , 2006, 26 (10) : 3404–3412. |
| [12] | 郑丽萍, 龙涛, 林玉锁, 于赐刚, 刘燕, 祝欣. Biolog-ECO解析有机氯农药污染场地土壤微生物群落功能多样性特征. 应用与环境学报 , 2013, 19 (5) : 759–765. |
| [13] | 罗希茜, 郝晓晖, 陈涛, 邓婵娟, 吴金水, 胡荣桂. 长期不同施肥对稻田土壤微生物群落功能多样性的影响. 生态学报 , 2009, 29 (2) : 740–748. |
| [14] | 白文波, 王春艳, 李茂松, 鲍顺淑, 何爽, 曹肆林, 宋吉青. 不同灌溉条件下保水剂对新疆棉花生长及产量的影响. 农业工程学报 , 2010, 26 (10) : 69–76. |
| [15] | Zirkler D, Peters A, Kaupenjohann M. Elemental composition of biogas residues:Variability and alteration during anaerobic digestion. Biomass and Bioenergy , 2014, 67 : 89–98. DOI:10.1016/j.biombioe.2014.04.021 |
| [16] | Pinto R, Ilileji K. On the performance of biobased carriers from distillers grains with solubles (DDGS)//ASABE Meeting Presentation, 2009:Paper number 095675. |
| [17] | Galvez A, Sinicco T, Cayuela M L, Mingorance M D, Fornasier F, Mondini C. Short term effects of bioenergy by-products on soil C and N dynamics, nutrient availability and biochemical properties. Agriculture, Ecosystems & Environment , 2012, 160 : 3–14. |
| [18] | Abubaker J, Risberg K, Pell M. Biogas residues as fertilisers-Effects on wheat growth and soil microbial activities. Applied Energy , 2012, 99 : 126–134. DOI:10.1016/j.apenergy.2012.04.050 |
| [19] | Terhoeven-Urselmans T, Scheller E, Raubuch M, Ludwig B, Joergensen R G. CO2 evolution and N mineralization after biogas slurry application in the field and its yield effects on spring barley. Applied Soil Ecology , 2009, 42 (3) : 297–302. DOI:10.1016/j.apsoil.2009.05.012 |
| [20] | 温晓霞, 殷瑞敬, 高茂盛, 艾绥龙. 不同覆盖模式下旱作苹果园土壤酶活性和微生物数量时空动态研究. 西北农林学报 , 2011, 20 (11) : 82–88. |
| [21] | 冯伟, 管涛, 王晓宇, 朱云集, 郭天财. 沼液与化肥配施对冬小麦根际土壤微生物数量和酶活性的影响. 应用生态学报 , 2011, 22 (4) : 1007–1012. |
| [22] | 李志斐, 王广军, 谢骏, 郁二蒙, 余德光, 夏耘, 魏南. 草鱼养殖池塘生物膜固着微生物群落碳代谢Biolog分析. 水产学报 , 2014, 38 (12) : 1985–1994. |
| [23] | 孙凤霞, 张伟华, 徐明岗, 张文菊, 李兆强, 张敬业. 长期施肥对红壤微生物生物量碳氮和微生物碳源利用的影响. 应用生态学报 , 2010, 21 (11) : 2792–2798. |
| [24] | Harch B D, Correll R L, Meech W, Kirkby C A, Pankurst C E. Using the Gini coefficient with Biolog Substrate utilization data to provide an alternative quantitative measure for comparing bacterial soil communities. Journal of Microbiological Methods , 1997, 30 (1) : 91–101. DOI:10.1016/S0167-7012(97)00048-1 |
| [25] | Zak J C, Willig M R, Moorhead D L, Wildman H G. Functional diversity of microbial communities:A quantitative approach. Soil Biology and Biochemistry , 1994, 26 (9) : 1101–1108. DOI:10.1016/0038-0717(94)90131-7 |
| [26] | Haack S K, Garchow H, Klug M J, Forney LJ. Analysis of factors affecting the accuracy, reproducibility, and interpretation of microbial community carbon source utilization patterns. Applied and Environmental Microbiology , 1995, 61 (4) : 1458–1468. |
| [27] | 李忠佩, 吴晓晨, 陈碧云. 不同利用方式下土壤有机碳转化及微生物群落功能多样性变化. 中国农业科学 , 2007, 40 (8) : 1712–1721. |
| [28] | 邱莉萍, 刘军, 王益权, 孙慧敏, 和文祥. 土壤酶活性与土壤肥力的关系研究. 植物营养与肥料学报 , 2004, 10 (3) : 277–280. |
| [29] | 孙波, 赵其国, 张桃林, 俞慎. 土壤质量与持续环境:Ⅲ.土壤质量评价的生物学指标. 土壤 , 1997 (5) : 225–234. |
| [30] | 吴等, 宋志文, 徐爱玲, 郑远, 夏岩, 等. 青岛市不同功能区冬季空气微生物群落代谢与多样性特征. 生态学报 , 2015, 35 (7) : 2277–2284. |
| [31] | 周丽霞, 丁明懋. 土壤微生物学特性对土壤健康的指示作用. 生物多样性 , 2007, 15 (2) : 162–171. |
| [32] | 程存刚, 赵德英, 吕德国, 姜曼, 杜国栋. 植物源有机物料对果园土壤微生物群落多样性的影响. 植物营养与肥料学报 , 2014, 20 (4) : 913–922. |
| [33] | 张海燕, 肖延华, 张旭东, 李军, 席联敏. 土壤微生物量作为土壤肥力指标的探讨. 土壤通报 , 2006, 37 (3) : 422–425. |
| [34] | 刘定辉, 舒丽, 陈强, 陈尚洪, 陈红琳, 朱钟麟. 秸秆还田少免耕对冲积土微生物多样性及微生物碳氮的影响. 应用与环境学报 , 2011, 17 (2) : 158–161. |
| [35] | 刘国顺, 李正, 敬海霞, 叶协锋, 史宏志, 王永, 杨超, 常栋. 连年翻压绿肥对植烟土壤微生物量及酶活性的影响. 植物营养与肥料学报 , 2010, 16 (6) : 1472–1478. |
| [36] | 李正, 刘国顺, 敬海霞, 叶协锋, 解昌盛, 向永光, 张文平, 杨超, 王永, 习相银. 绿肥与化肥配施对植烟土壤微生物量及供氮能力的影响. 草叶学报 , 2011, 20 (6) : 126–134. |