 2016, Vol. 36
2016, Vol. 36文章信息
- 和润莲, 陈亚梅, 邓长春, 杨林, 刘军伟, 杨万勤, 张健, 刘洋
- HE Runlian, CHEN Yamei, DENG Changchun, YANG Lin, LIU Junwei, YANG Wanqin, ZHANG Jian, LIU Yang.
- 川西高山林线三种灌木凋落叶分解中的无脊椎动物多样性
- Invertebrate diversity in foliar litter of three shrubs in the alpine timberline of western Sichuan
- 生态学报[J]. 2016, 36(17): 5497-5507
- Acta Ecologica Sinica[J]. 2016, 36(17): 5497-5507
- http://dx.doi.org/10.5846/stxb201502250380
-
文章历史
- 收稿日期: 2015-02-25
- 网络出版日期: 2015-12-14

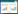




研究区区域位于四川省阿坝藏族、羌族自治州理县米亚罗鹧鸪山(31°50'-31°53'E,102°40'-102°44'N) 处于青藏高原东缘褶皱带最外缘部分,岷江上游支流杂谷脑河上游地区。鹧鸪山山体海拔3 200-4 800 m,山地垂直地带性明显,自河谷至山顶依次分布的植被类型有针阔混交林、暗针叶林、高山疏林灌丛、高山草甸,4 500m以上为高山荒漠和积雪带。气候属于冬寒夏凉的高寒气候,年平均气温6-12 ℃,1月平均气温-8 ℃,7月平均气温12.6 ℃,年积温1 200-1 400 ℃,年降水量600-1 100 mm,年蒸发量1 000-1 900 mm。2012年10月底到2013年11月初一年的日均温变化情况如图 1。由图 1知,一年内林线上温度变化波动很大,而针叶林上温度变化平缓。从2012年10月底到2013年4月底,日均温在0℃以下,从5月份开始逐渐回温,6月份达到峰值,7、8月份的日均温保持较高。地形以石质山地为主,土壤具有粗骨、石砾含量高和薄层等特征。从低海拔到高海拔,土壤类型依次为山地暗棕壤、棕色针叶林土、高山草甸土。主要灌木树种有大理杜鹃(Rhododendron taliense)、皱皮杜鹃(R. wiltonii)、高山杜鹃(R. lapponicum)、红花花楸(Sorbus rufopilosa)、柳叶忍冬(Lonicera lanceolata)、越桔叶忍冬(L. myrtillus)、窄叶鲜卑花(Sibiraea angustata)和高山柳(Salix cupularis)等。每年从4月到10月为生长季末期,冬季雪被末期明显,从10月到次年4月,长达6-7个月。
2 研究方法 2.1 实验设计2012年10月在鹧鸪山针叶林和林线上采集优势灌木物种的凋落叶,包括高山柳、红毛花楸和高山杜鹃3个物种,带回室内将其风干,每个物种称取10 g装入20 cm × 20 cm(孔径为1 mm)的分解袋中,每个物种称取12袋,共36个凋落袋。10月底分别将凋落物袋平铺埋设在海拔3900m的针叶林和海拔4000m的林线地表,每个海拔每个物种放置6袋,并用钉子固定位置。同时每个海拔随机选取一个凋落袋,袋内埋设一个纽扣式温度记录仪(iButton DS1923-F5,Maxim/Dallas Semiconductor,Sunnyvale,CA,USA),设定为每3h记录1次温度数据,计算冻融频次。同时,在放置凋落袋的附近用15竹竿插入土壤中,用作雪被厚度测定的标记,在雪被末期每月观测记录雪被的厚度情况。
2.2 样品处理2013年5月1日和2013年11月7日分别代表雪被末期末和生长季末期,分别采集每个处理的凋落叶分解袋各3袋,将所采的分解袋装入密封而透气的黑布袋里面低温保存,迅速带回室内,先将袋外的泥土残落物轻轻去除,然后将袋内的残落物放置于改良的Tullgren干漏斗中分离提取中小型无脊椎动物,采用体式解剖镜和生物显微镜镜检计数和分类,并参照《中国土壤动物检索图鉴》[18]鉴定到科。分离结束后,清除残落物表面的土壤颗粒和混入的杂物,60℃下烘干至恒重,称量残落物干重。
2.3 数据处理与分析采用Shannon指数H'、Simpson优势度指数C、Pielou均匀度指数J和密度-类群指数DG计算各海拔各无脊椎动物群落多样性。各指数计算公式如下[8]:
(1)Shannon多样性指数
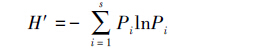
(2)Simpson优势度指数
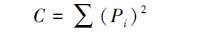
(3)Pielou均匀度指数
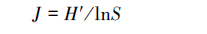
(4)Margalef 丰富度指数
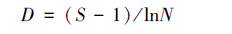
式中,ni为第i 个物种的个体数,S 为类群数,N 为无脊椎动物总个体数,Pi为第i 类群的百分比。
各类群优势度的划分:个体数量占捕获总数10.0% 以上的为优势类群,1.0%-10.0% 的为常见类群,1.0%以下的为稀有类群[1, 5]。数据分析在SPSS17.0中进行,采样方差分析和独立样本T检验比较环境因子和无脊椎动物群落结构间的差异,Pearson相关性分析环境因子与无脊椎动物群落结构特征的相关性,显著性水平设定为a=0.05。相关图表制作在Excel中完成。
3 结果与分析 3.1 凋落叶分解的环境因子2012 年10月30日-2013年11月7日针叶林和林线上的日平均温度变化如图 1所示。针叶林和林线上温度变化趋势一致,但针叶林日均温变化平稳,而林线日均温波动幅度较大。从10月底到次年1月,气温逐渐下降,12月底1月初温度最低,从1月中下旬开始,气温逐渐回升,到5月份日均温达到0℃以上,在7月份达到峰值,到次年10月份温度又开始降到0℃以下。

|
| 图 1 针叶林和林线凋落袋内年日均温度动态 Fig. 1 Dynamics of daily mean temperature in litter bag in the coniferous forest and timberline within a year |
雪被期针叶林和林线上雪被厚度如图 2所示。针叶林和林线上的雪被均在11月份最厚,3月初针叶林和林线上没有监测到雪被厚度。T检验发现,雪被期针叶林和林线上的雪被厚度均差异不显著。两个时期针叶林和林线上的平均温度、雪被厚度、冻融频次、正积温和负积温如表 1所示。雪被期日均温在0℃以下,生长季节日均温在0℃以上,针叶林内平均雪被厚度高于林线上,且两个时期林线上的日均温、冻融频次、正积温和负积温均高于针叶林上,进一步说明林线上温度变化剧烈。

|
| 图 2 雪被期针叶林和林线的雪被厚度 Fig. 2 Snow depth of the alpine forest and timberline during snow-covered season |
| 时期Date | 林线交错带 Ecotone | 平均温度(ST) Average temperature/℃ | 雪被厚度(SC) Snow thickness/cm | 冻融循环次数(FC) Frequency of freeze-thaw cycle/time | 正积温(PAT) Positive accumulated temperature | 负积温(NAT) Negative accumulated temperature |
| 雪被期 | 针叶林 | -2.65 | 7.21 | 135 | 1.69 | -784.06 |
| Snow covered season | 林线 | -1.89 | 6.44 | 270 | 310.63 | -391 |
| 生长季节 | 针叶林 | 6.46 | 0 | 37 | 883.44 | -15.81 |
| Growing season | 林线 | 8.21 | 0 | 73 | 1573.31 | -11 |
雪被末期和生长季末期2次采样凋落物中中小型无脊椎动物共714只,隶属1门3纲12目32类(科)(表2),不同季节林线交错带3种灌木凋落叶中获得的无脊椎动物群落结构有所变化。雪被末期3种凋落叶中获得无脊椎动物18科,共111只,其中针叶林中获得15科,28只,林线上获得10科,83只。生长季末期3种凋落叶中获得无脊椎动物25科,共603只,其中针叶林中获得无脊椎动物14科,335只,林线上获得无脊椎动物21科,268只。若甲螨科和隐翅甲科仅出现在雪被末期针叶林上的高山杜鹃凋落叶中,大蚊科和管蓟马亚科仅出现在雪被末期针叶林上红毛花楸的凋落叶中,罗甲螨科仅出现在雪被末期林线上高山杜鹃凋落叶中,而寄螨科和气肢虫科仅出现在生长季末期林线上的红毛花楸凋落叶中,缨甲科和卷甲虫科仅出现在生长季末期针叶林上高山杜鹃凋落叶中,同翅目仅出现在生长季末期林线上高山杜鹃凋落叶中。两个季节针叶林和林线上无脊椎动物优势类群如表 3。雪被末期,针叶林中弹尾目、中气门亚目、甲螨亚目和双翅目幼虫为优势类群,前气门亚目、鞘翅目和缨翅目为常见类群;林线上以前气门亚目和中气门亚目为优势类群,以弹尾目、甲螨亚目和双翅目幼虫为常见类群。生长季末期,针叶林中以弹尾目和中气门亚目为优势类群,以中气门亚目、甲螨亚目和鳞翅目幼虫为常见类群,以双翅目幼虫、鞘翅目、等足目和啮目为稀有类群;林线上以弹尾目、前气门亚目和中气门亚目为优势类群,以甲螨亚目、钳亚目和同翅目为常见类群,以双翅目幼虫、鳞翅目幼虫、等足目和啮目为稀有类群。缨翅目仅出现在雪被末期,鳞翅目幼虫、双尾目、等足目、啮目和同翅目仅在生长季末期出现。无脊椎动物群落结构同时受到凋落叶物种和季节的影响。
| 时期 Date | 雪被末期 Snow covered season | 生长季末期 Growing season | |||||||||||||
| 林线交错带 Ecotone | 针叶林Coniferous forest | 林线Timberline | 针叶林Coniferous forest | 林线Timberline | |||||||||||
| 类群 Group | Sc | Sr | Rl | Sc | Sr | Rl | Sc | Sr | Rl | Sc | Sr | Rl | |||
| 棘跳科Onychiuridae | 0 | 2 | 2 | 0 | 1 | 0 | 28 | 41 | 1 | 0 | 0 | 4 | |||
| 跳虫科Poduridae | 0 | 0 | 3 | 5 | 0 | 0 | 84 | 84 | 0 | 12 | 9 | 17 | |||
| 长角跳科Entomobryidae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | |||
| 等级跳科Isotomidae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 5 | |||
| 赤螨科Erythraeidae | 0 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 1 | |||
| 矮蒲螨科Pygmephoridae | 0 | 0 | 0 | 41 | 11 | 0 | 0 | 0 | 20 | 0 | 0 | 49 | |||
| 长须螨科Stigmaeidae | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 1 | 1 | 0 | 0 | 2 | |||
| 派盾螨科 Parholaspididae | 0 | 0 | 5 | 0 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | |||
| 美绥螨科Ameroseiidae | 0 | 3 | 2 | 0 | 11 | 4 | 0 | 0 | 0 | 32 | 1 | 35 | |||
| 维螨科Veigaiidae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 48 | |||
| 足角螨科Podocinidae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 5 | |||
| 寄螨科Parasitidae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | |||
| 厉螨科Laelapidae | 0 | 1 | 0 | 0 | 0 | 0 | 34 | 12 | 1 | 0 | 0 | 0 | |||
| 丽甲螨科Liacaridae | 0 | 1 | 0 | 0 | 1 | 0 | 3 | 1 | 5 | 6 | 0 | 9 | |||
| 大翼甲螨科Galumnidae | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | |||
| 若甲螨科Oribatuloidae | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| 步甲螨科Carabodidae | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| 懒甲螨科Nothridae | 0 | 0 | 0 | 1 | 1 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| 古甲螨科Palaeacaridae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 2 | 0 | 3 | |||
| 罗甲螨科Lohmannidae | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| 长足虻科Dolichopodidae | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 2 | 0 | 0 | 0 | |||
| 摇蚊科Chironomidae | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | |||
| 大蚊科Tipulidae | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| 隐翅甲科Staphilinidae | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| 缨甲科Ptiliidae | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | |||
| 尺蛾科Geometridae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 2 | 1 | 0 | 0 | |||
| 管蓟马亚科Phlaeothripinae | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| 铗叭科Japygidae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 2 | |||
| 卷甲虫科Armadillidiidae | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | |||
| 气肢虫科Trachelipidae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | |||
| 鼠啮科Myopsocidae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | |||
| 同翅目Homoptere | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | |||
| 类群数Group number | 0 | 9 | 8 | 4 | 5 | 5 | 9 | 8 | 8 | 8 | 8 | 16 | |||
| 个体数Ind. number/只 | 0 | 12 | 16 | 49 | 25 | 9 | 158 | 144 | 33 | 60 | 19 | 189 | |||
| Sc:高山柳Salix cupularis; Sr:红毛花楸Sorbus rufopilosa; Rl:高山杜鹃Rhododendron lapponicum | |||||||||||||||
| 时期 Date | 雪被末期 Snow covered season | 生长季末期 Growing season | 合计Total | ||||||||||||||||||||
| 林线交错带 Ecotone | 针叶林Coniferous forest | 林线 Timberline | 针叶林Coniferous forest | 林线 Timberline | 针叶林Coniferous forest | 林线 Timberline | |||||||||||||||||
| 类群 Group | 个体数 | % | 优势度Abundance | 个体数 | % | 优势度Abundance | 个体数 | % | 优势度Abundance | 个体数 | % | 优势度Abundance | 个体数 | % | 优势度Abundance | 个体数 | % | 优势度Abundance | |||||
| 弹尾目Collembola | 7 | 25 | + + + | 6 | 7.23 | + + | 238 | 71.04 | + + + | 54 | 20.15 | + + + | 245 | 67.48 | + + + | 60 | 17.09 | + + + | |||||
| 前气门亚目Prostigrnata | 1 | 3.57 | + + | 54 | 65.06 | + + + | 26 | 7.76 | + + | 55 | 20.52 | + + + | 27 | 7.44 | + + | 109 | 31.05 | + + + | |||||
| 中气门亚目Mesostigmata | 11 | 39.29 | + + + | 15 | 18.07 | + + + | 49 | 14.63 | + + + | 127 | 47.39 | + + + | 60 | 16.53 | + + + | 142 | 40.46 | + + + | |||||
| 甲螨亚目Oribatid | 4 | 14.29 | + + + | 7 | 8.43 | + + | 11 | 3.28 | + + | 21 | 7.83 | + + | 15 | 4.13 | + + | 28 | 7.98 | + + | |||||
| 双翅目幼虫Diptera larvae | 3 | 10.71 | + + + | 1 | 1.21 | + + | 3 | 0.9 | + | 1 | 0.37 | + | 6 | 1.65 | + + | 2 | 0.57 | + | |||||
| 鞘翅目Coleoptera | 1 | 3.57 | + + | 0 | 0 | 1 | 0.3 | + | 0 | 0 | 2 | 0.55 | + | 0 | 0 | ||||||||
| 鳞翅目幼虫Lepidoptera larvae | 0 | 0 | 0 | 0 | 5 | 1.49 | + + | 1 | 0.37 | + | 5 | 1.38 | + + | 1 | 0.28 | + | |||||||
| 缨翅目Thysanoptera | 1 | 3.57 | + + | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0.28 | + | 0 | 0 | |||||||||
| 钳亚目Dicellurata | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 1.5 | + + | 0 | 0 | 4 | 1.14 | + + | |||||||||
| 等足目Isopoda | 0 | 0 | 0 | 0 | 1 | 0.3 | + | 1 | 0.37 | + | 1 | 0.28 | + | 1 | 0.28 | + | |||||||
| 啮目Psocoptera | 0 | 0 | 0 | 0 | 1 | 0.3 | + | 1 | 0.37 | + | 1 | 0.28 | + | 1 | 0.28 | + | |||||||
| 同翅目Homoptera | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 1.13 | + + | 0 | 0 | 3 | 0.87 | + | |||||||||
| 合计 Total | 28 | 100 | 83 | 100 | 335 | 100 | 268 | 100 | 363 | 100 | 351 | 100 | + | ||||||||||
| 优势类群:密度占总密度的10%以上 Average density accounted for more than 10% of the total; 常见类群:密度占总密度的1%-10% Average density accounted for 1%-10% of the total; 稀有类群:密度占总密度的1%以下 Average density accounted for less than 1% of the total | |||||||||||||||||||||||
两个时期针叶林和林线上灌木凋落叶中无脊椎动物个体密度和类群密度如图 3所示。个体密度和类群密度在生长季末期均高于雪被末期,并且个体密度在针叶林上高山柳凋落叶中(F =7.682,P <0.05)和林线上高山杜鹃凋落叶中差异显著(F =13.609,P <0.05),类群密度在林线上高山杜鹃凋落叶中差异显著(F =12.914,P <0.05)。说明生长季更适合不同类群无脊椎动物的生存,获得的个体数量也比雪被期高。

|
| 图 3 无脊椎动物个体密度和类群密度 Fig. 3 Individual density and group density of invertebrate |
采用T检验,比较雪被末期和生长季末期林线和针叶林的各物种凋落叶的无脊椎动物多样性(图 4)。在雪被末期,高山柳凋落叶内没有无脊椎动物外,Shannon指数H、Pielou指数J和Margalef指数D在针叶林中高于林线上,Simpson指数C相反,并且高山柳凋落叶中Shannon指数H(F=10.048,P<0.05)、Pielou指数J(F=10.047,P<0.05)和Simpson指数C(F=10.305,P<0.05)在针叶林和林线上差异显著,红毛花楸凋落叶中Pielou指数J(F=9.844,P<0.05)在针叶林和林线上差异显著;在生长季末期,高山柳凋落叶中的Pielou指数J、红毛花楸和高山杜鹃凋落叶中的Simpson指数C针叶林上高于林线上,各凋落叶中的Shannon指数H、Margalef指数D、高山柳和红毛花楸凋落叶中的Pielou指数J和高山柳凋落叶中的Simpson指数C在林线上高于针叶林上,并且差异不显著(P>0.05)。总体来说,生长季末期无脊椎动物较雪被末期丰富,林线上较针叶林丰富。

|
| 图 4 雪被末期和生长季节末期凋落物无脊椎动物群落多样性季节动态 Fig. 4 Seasonal dynamics of invertebrate community diversity of snow covered season and growing season |
凋落物种类、海拔梯度和季节对无脊椎动物群落结构均有一定的影响(表 4)。凋落物物种对多样性指数H(P<0.05)、J和D(P<0.01)有显著的影响;海拔梯度对C指数(P<0.01)和无脊椎动物类群密度(P<0.05)有显著影响;季节差异对无脊椎动物多样性指数H、D、个体密度和类群密度均有极显著的影响(P<0.01)。凋落物种类和海拔梯度的交互作用对多样性指数J、C、类群密度(P<0.05)和个体密度(P<0.01)均有显著影响;凋落物种类和季节的交互作用对多样性指数H和J影响显著;海拔梯度和季节的交互作用对多样性指数H、C、D以及对类群密度均有显著影响;凋落物种类、海拔梯度和季节差异3个因子的交互对无脊椎动物个体密度和类群密度、多样性指数H、J有显著影响。
| Sources | H | J | C | D | 个体密度Individual density | 类群密度Group density |
| 凋落物物种 Litter Species | 4.824* | 10.141** | 0.896 | 6.027** | 0.491 | 0.37 |
| 海拔梯度 Altitude | 0.418 | 0.798 | 10.545** | 0.358 | 2.126 | 4.573* |
| 季节 Season | 31.779** | 3.952 | 2.656 | 15.899** | 16.481** | 89.669** |
| 凋落物物种×海拔梯度 Litter Species×Altitude | 0.853 | 5.090* | 5.474* | 0.965 | 5.936** | 4.703* |
| 凋落物物种×季节 Litter Species×Season | 4.569* | 5.515* | 3.166 | 2.915 | 0.634 | 1.442 |
| 海拔梯度×季节 Altitude×Season | 6.534* | 0.037 | 30.316** | 10.868** | 0.379 | 4.655* |
| 凋落物物种×海拔梯度×季节Litter Species×Altitude×Season | 6.560** | 7.849** | 1.366 | 2.692 | 5.660* | 3.545* |
| H:香浓多样性指数 Shannon index; J:均匀度指数 Pielou index; C:优势度指数 Simpson index; D:丰富度指数Margalef index; * P < 0.05,**P < 0.01 | ||||||
川西高山针叶林和林线上灌木凋落叶中无脊椎动物群落结构与环境因子的相关分析结果如表 5。无脊椎动物群落的多样性指数H、D、个体密度和类群密度均与雪被厚度和冻融频次极显著负相关,与正积温显著正相关,多样性指数J与日均温和正、负积温正相关,与雪被厚度和冻融频次负相关,多样性指数C与冻融频次极显著正相关。相关分析结果说明,温度、雪被和冻融循环频率对无脊椎动物群落结构和多样性的影响显著。
| 环境因子Climatic factors | H | J | C | D | 个体密度Individual density | 类群密度Group density |
| ST | 0.550** | 0.234 | -0.017 | 0.444** | 0.328* | 0.797** |
| SC | -0.560** | -0.217 | 0.118 | -0.439** | -0.483** | -0.789** |
| FC | -0.517** | -0.137 | 0.470** | -0.453** | -0.472** | -0.622** |
| PAT | 0.582** | 0.234 | -0.104 | 0.498** | 0.354* | 0.812** |
| NAT | 0.457** | 0.216 | 0.131 | 0.319 | 0.409* | 0.714** |
| **在0.01水平上显著相关;*在0.05水平上显著相关 | ||||||
在全球尺度上,气候因素对凋落物的分解起主导作用,温度和湿度被认为是影响凋落物分解主要的气候因子[19],其次是凋落物质量和土壤生物作用。最近的一些分解模型研究发现,气候和凋落物质量可以解释全球凋落物分解速率的60%-70%,另外的30%-40%主要是生物因素的作用[20]。从全球无脊椎动物对凋落物分解的贡献来看,如果排除无脊椎动物的影响,凋落物分解速率将显著降低35%[21]。凋落物为土壤生物提供食物来源和能量,而土壤生物的活动对凋落物的分解起促进作用[19]。无脊椎动物群落结构和多样性受到气候、植被类型、土壤、凋落物种类及小气候的影响[2, 8, 14, 22]。研究发现,川西高山森林和林线由于郁闭度和植被类型变化,温度、雪被厚度和冻融频次等环境因子差异明显。因此,导致海拔梯度上无脊椎动物群落结构和多样性的差异,尤其是季节变化对无脊椎动物群落结构和多样性的影响最为显著,生长季末期无脊椎动物个体密度和类群密度、多样性指数H、D均高于雪被末期,这与团队前期研究结果类似[7-9, 23],与David和Gillon[24]与wang[2]等认为温湿度和无脊椎动物多样性密切相关的研究结果一致,进一步证实了季节变化造成的温度、雪被等环境因子影响着无脊椎动物群落组成及多样性[14, 23, 19]。气候是影响无脊椎动物群落结构和多样性的决定性因素[21,25],生长季节更高的温度更适合无脊椎动物的生存繁衍[8]。川西高山地区冬季环境严酷,低温不仅严重制约了无脊椎动物活动,导致无脊椎动物群落表现出较低的活性甚至休眠状态[26-28],同时还冻结了凋落物的养分资源,使得可利用的食物来源严重匮乏[23],无脊椎动物难以取食。然而,雪被末期无脊椎动物数量仍占到总数的15.5%,说明在冬季恶劣环境条件下,仍有部分类群和数量的无脊椎动物生存活动[8]。伴随着生长季的回暖,部分休眠或蛰伏无脊椎动物开始复苏[29],个体数量和类群数显著上升。同时,生长季节水热同季,植被生长旺盛,微生物活性较高,林线交错带土壤微生物数量和生物量表现为生长季末期高于雪被末期[30],这为无脊椎动物的生存和繁衍提供了良好的食物来源[31-32]。
海拔差异导致无脊椎动物所处的微环境发生变化,对高山森林和林线上无脊椎动物多样性及类群密度有重要影响。本研究发现,海拔梯度对凋落物中无脊椎动物优势度指数C和类群密度影响显著。雪被期林线上获得的个体数和类群数较针叶林丰富,生长季节针叶林内获得的个体数较多,但类群数在林线上更丰富(表 2),进一步佐证了林线交错带具有较高的物种丰富度[5, 12]。两个海拔获得的部分优势类群和常见类群相同,但稀有类群差异较大,如鞘翅目、缨翅目仅在针叶林出现,同翅目仅出现在林线上,可能是因为高山林线处于森林向灌丛转变的过渡带上,从暗针叶林过渡到灌丛草甸,植被类型与土壤环境因子发生了显著的变化,导致林线交错区的光强、温湿度、雪被、冻融循环等微环境产生很大的差异,这些变化对无脊椎动物的食物源和栖息环境产生很大的影响,它们直接或间接作用于无脊椎动物群落,从而导致无脊椎动物群落结构和多样性发生了显著变化[2, 5, 14]。在高纬度和高海拔地区,雪被能对这些控制凋落物分解的气候因子产生重要影响[33]。Templer等[34]研究发现,美国北方阔叶林冬季雪被厚度减少和持续时间缩短,降低了森林地面的节肢动物的丰度和多样性,改变了群落结构和组成。在未来气候变暖条件下,随着气温升高,冬季雪线位置的上移[35],高山生态系统中无脊椎动物群落结构势必发生改变,可能使高寒物种、地方特有种或其中的优势种因为生境的改变而濒临灭绝或被其他物种替代[36]。
凋落物的种类、基质质量影响无脊椎动物的数量分布和类群组成[37]。本研究发现,物种对凋落叶中无脊椎动物多样性指数H、J和D影响显著,3种灌木凋落叶中无脊椎动物捕获量以高山杜鹃最多,红毛花楸次之,高山柳最少。长角跳科、若甲螨科、步甲螨科、罗甲螨科和隐翅甲科仅出现在高山杜鹃凋落叶中,寄螨科、摇蚊科、大蚊科、管蓟马亚科和气肢虫科仅出现在红毛花楸凋落叶中,缨甲科和卷甲虫科仅出现在高山柳凋落叶中。值得注意的是,雪被期针叶林内高山柳凋落叶中没有分离到无脊椎动物,可能是无脊椎动物迁移和活动的偶然性造成的。方差分析结果表明,季节、海拔梯度、凋落物种类两两交互或3个因子的交互对无脊椎动物个体密度和类群密度、多样性指数有显著影响,且季节和海拔梯度的交互作用更加显著,说明季节对各海拔的环境因子影响程度不同,对不同凋落物种类的影响也有差异,3个因子综合作用对土壤动物群落结构造成影响。由于高山植物对气候变暖的响应最为敏感[38],气候变暖可能导致林线上移以及灌丛分布密度增加[13, 38],从而影响凋落物种类和基质质量[15-16],势必改变无脊椎动物群落的分布格局和多样性[25],如灌木凋落叶中的特有种罗甲螨科、缨甲科等可能随灌木密度的增加和凋落物输入量增大而增加。同时,未来气候变化将改变无脊椎动物对凋落物分解的作用方式,尤其是高寒地区的气候变化对凋落物分解的影响将更为显著[19],随着降水的增多和最低温的升高,无脊椎动物对凋落物分解的作用增大[21],表现在直接加速凋落物分解和有机碳库的释放,间接地提高无脊椎动物对凋落物分解的贡献率。
| [1] | Swift M J, Heal O W, Anderson J M. Decomposition in Terrestrial Ecosystems. Oxford: Blackwell Scientific Publications, 1979. |
| [2] | Wang S J, Ruan H H, Wang B. Effects of soil microarthropods on plant litter decomposition across an elevation gradient in the Wuyi Mountains. Soil Biology and Biochemistry , 2009, 41 (5) : 891–897. DOI:10.1016/j.soilbio.2008.12.016 |
| [3] | Donoso D A, Johnston M K, Kaspari M. Trees as templates for tropical litter arthropod diversity. Oecologia , 2010, 164 (1) : 201–211. DOI:10.1007/s00442-010-1607-3 |
| [4] | Gavazov K S. Dynamics of alpine plant litter decomposition in a changing climate. Plant and Soil , 2010, 337 (1/2) : 19–32. |
| [5] | 黄旭, 文维全, 张健, 杨万勤, 刘洋, 闫帮国, 黄玉梅. 川西高山典型自然植被土壤动物多样性. 应用生态学报 , 2010, 21 (1) : 181–190. |
| [6] | 夏磊, 吴福忠, 杨万勤. 季节性冻融期间土壤动物对岷江冷杉凋落叶质量损失的贡献. 植物生态学报 , 2011, 35 (11) : 1127–1135. |
| [7] | 夏磊, 吴福忠, 杨万勤, 谭波. 川西亚高山森林凋落物分解初期土壤动物对红桦凋落叶质量损失的贡献. 应用生态学报 , 2012, 23 (2) : 301–306. |
| [8] | 谭波, 吴福忠, 杨万勤, 夏磊, 杨玉莲, 王奥. 川西亚高山/高山森林大型土壤动物群落多样性及其对季节性冻融的响应. 生物多样性 , 2012, 20 (2) : 215–223. |
| [9] | 刘瑞龙, 李维民, 杨万勤, 谭波, 王文君, 徐振锋, 吴福忠. 土壤动物对川西高山/亚高山森林凋落物分解的贡献. 应用生态学报 , 2013, 24 (12) : 3354–3360. |
| [10] | Wu P F, Liu X L, Liu S R, Wang J X, Wang Y. Composition and spatio-temporal variation of soil microarthropods in the biodiversity hotspot of northern Hengduan Mountains, China. European Journal of Soil Biology , 2014, 62 : 30–38. DOI:10.1016/j.ejsobi.2014.02.013 |
| [11] | Holtmeier F K. Ecological aspects of climatically caused timberline fluctuations//Benist M, ed. Mountain Environments in Changing Climates. Routledge. New York:London, 1994, 220-233. |
| [12] | 石培礼, 李文华, 王金锡, 刘兴良. 四川卧龙亚高山林线生态交错带群落的种多度关系. 生态学报 , 2000, 20 (3) : 384–389. |
| [13] | 刘洋, 张健, 杨万勤. 高山生物多样性对气候变化响应的研究进展. 生物多样性 , 2009, 17 (1) : 88–96. |
| [14] | 和润莲, 陈亚梅, 邓长春, 杨万勤, 张健, 刘洋. 雪被期川西高山林线交错带两种地被物凋落物分解与土壤动物多样性. 应用生态学报 , 2015, 26 (3) : 723–731. |
| [15] | Walther G R, Beißner S, Burga C A. Trends in the upward shift of alpine plants. Journal of Vegetation Science , 2005, 16 (5) : 541–548. DOI:10.1111/j.1654-1103.2005.tb02394.x |
| [16] | Klasner F L, Fagre D B. A half century of change in alpine treeline patterns at Glacier National Park, Montana. U.S.A. Arctic, Antarctic, and Alpine Research , 2002, 34 (1) : 49–56. DOI:10.2307/1552508 |
| [17] | Wardle D A, Yeates G W, Barker G M, Bonner K I. The influence of plant litter diversity on decomposer abundance and diversity. Soil Biology and Biochemistry , 2006, 38 (5) : 1052–1062. DOI:10.1016/j.soilbio.2005.09.003 |
| [18] | 尹文英, 胡圣豪, 沈韫芬, 宁应之, 孙希达, 吴纪华, 诸葛燕, 张云美, 王敏, 陈建英, 徐成纲, 梁彦龄, 王洪铸, 杨潼, 陈德牛, 张国庆, 宋大祥, 陈军, 梁来荣, 胡成业, 王慧芙, 张崇洲, 匡溥人, 陈国孝, 赵立军, 谢荣栋, 张骏, 刘宪伟, 韩美贞, 毕道英, 肖宁年, 杨大荣. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998. |
| [19] | Aerts R. The freezer defrosting:global warming and litter decomposition rates in cold biomes. Journal of Ecology , 2006, 94 (4) : 713–724. DOI:10.1111/jec.2006.94.issue-4 |
| [20] | Parton W, Silver W L, Burke I C, Grassens L, Harmon M E, Currie W S, King J Y, Adair E C, Brandt L A, Hart S C, Fasth B. Global-scale similarities in nitrogen release patterns during long-term decomposition. Science , 2007, 315 (5810) : 361–364. DOI:10.1126/science.1134853 |
| [21] | García-Palacios P, Maestre F T, Kattge J, Wall D H. Climate and litter quality differently modulate the effects of soil fauna on litter decomposition across biomes. Ecology Letters , 2013, 16 (8) : 1045–1053. DOI:10.1111/ele.2013.16.issue-8 |
| [22] | Maharning A R, Mills A A A, Adl S M. Soil community changes during secondary succession to naturalized grasslands. Applied Soil Ecology , 2009, 41 (2) : 137–147. DOI:10.1016/j.apsoil.2008.11.003 |
| [23] | Tan B, Wu F Z, Yang W Q, Liu L, Yu S. Characteristics of soil animal community in the subalpine/alpine forests of western Sichuan during onset of freezing. Acta Ecologica Sinica , 2010, 30 (2) : 93–99. DOI:10.1016/j.chnaes.2010.03.008 |
| [24] | David J F, Gillon D. Combined effects of elevated temperatures and reduced leaf litter quality on the life-history parameters of a saprophagous macroarthropod. Global Change Biology , 2009, 15 (1) : 156–165. DOI:10.1111/gcb.2009.15.issue-1 |
| [25] | Wall D H, Bradford M A, John M G S T, Trofymow J A, Behan-Pelletier V, Bignell D E, Dangerfield J M, Parton W J, Rusek J, Voigt W, Wolters V, Gardel H Z, Ayuke F O, Bashford R, Beljakova O I, Bohlen P J, Brauman A, Flemming S, Henschel J R, Johnson D L, Jones T H, Kovarova M, Kranabetter J M, Kutny L, Lin K C, Maryati M, Masse D, Pokarzhevskii A, Rahman H, Sabará M G, Salamon J A, Swift M J, Varela A, Vasconcelos H L, White D, Zou X M. Global decomposition experiment shows soil animal impacts on decomposition are climate-dependent. Global Change Biology , 2008, 14 (11) : 2661–2677. |
| [26] | Wu F Z, Yang W Q, Zhang J, Deng R J. Litter decomposition in two subalpine forests during the freeze-thaw season. Acta Oecologica , 2010, 36 (1) : 135–140. DOI:10.1016/j.actao.2009.11.002 |
| [27] | 王振中, 张友梅, 邢协加. 土壤环境变化对土壤动物群落影响的研究. 土壤学报 , 2002, 39 (6) : 892–897. |
| [28] | Briones M J I, Ostle N J, McNamara N P, Poskitt J. Functional shifts of grassland soil communities in response to soil warming. Soil Biology and Biochemistry , 2009, 41 (2) : 315–322. DOI:10.1016/j.soilbio.2008.11.003 |
| [29] | Campbell J L, Mitchell M J, Groffman P M, Christenson L M, Hardy J P. Winter in northeastern North America:a critical period for ecological processes. Frontiers in Ecology and the Environment , 2005, 3 (6) : 314–322. DOI:10.1890/1540-9295(2005)003[0314:WINNAA]2.0.CO;2 |
| [30] | 刘洋, 张健, 闫帮国, 黄旭, 徐振锋, 吴福忠. 青藏高原东缘高山森林-苔原交错带土壤微生物生物量碳?氮和可培养微生物数量的季节动态. 植物生态学报 , 2012, 36 (5) : 382–392. |
| [31] | 苗雅杰, 殷秀琴. 小兴安岭红松阔叶混交林土壤动物群落研究. 林业科学 , 2005, 41 (2) : 204–209. |
| [32] | Gongalsky K B, Persson T, Pokarzhevskii A D. Effects of soil temperature and moisture on the feeding activity of soil animals as determined by the bait-lamina test. Applied Soil Ecology , 2008, 39 (1) : 84–90. DOI:10.1016/j.apsoil.2007.11.007 |
| [33] | Baptist F, Yoccoz N G, Choler P. Direct and indirect control by snow cover over decomposition in alpine tundra along a snowmelt gradient. Plant and Soil , 2010, 328 (1/2) : 397–410. |
| [34] | Templer P H, Schiller A F, Fuller N W, Socci A M, Campbell J L, Drake J E, Kunz T H. Impact of a reduced winter snowpack on litter arthropod abundance and diversity in a northern hardwood forest ecosystem. Biology and Fertility of Soils , 2012, 48 (4) : 413–424. DOI:10.1007/s00374-011-0636-3 |
| [35] | IP CC. Climate Change 2007:the physical science basis//Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK: Cambridge University Press, 2007. |
| [36] | Klanderud K, Totland Ø. Simulated climate change altered dominance hierarchies and diversity of an alpine biodiversity hotspot. Ecology , 2005, 86 (8) : 2047–2054. DOI:10.1890/04-1563 |
| [37] | Siemann E, Tilman D, Haarstad J, Ritchie M. Experimental tests of the dependence of arthropod diversity on plant diversity. The American Naturalist , 1998, 152 (5) : 738–750. DOI:10.1086/286204 |
| [38] | 陈建国, 杨扬, 孙航. 高山植物对全球气候变暖的响应研究进展. 应用与环境生物学报 , 2011, 17 (3) : 435–446. |