 2016, Vol. 36
2016, Vol. 36文章信息
- 张义飞, 王平, 毕琪, 张忠辉, 杨允菲
- ZHANG Yifei, WANG Ping, BI Qi, ZHANG Zhonghui, YANG Yunfei.
- 不同强度盐胁迫下AM真菌对羊草生长的影响
- The effect of the arbuscular mycorrhizal fungi on the growth of Leymus chinensis under saline stress of different intensities
- 生态学报[J]. 2016, 36(17): 5467-5476
- Acta Ecologica Sinica[J]. 2016, 36(17): 5467-5476
- http://dx.doi.org/10.5846/stxb201502180372
-
文章历史
- 收稿日期: 2015-02-18
- 网络出版日期: 2015-12-14





2. 东北师范大学环境学院, 国家环境保护湿地生态与植被恢复重点实验室, 长春 130117;
3. 东北师大附属中学, 长春 130022;
4. 东北师范大学生命科学学院, 草地科学研究所, 长春 130024
2. State Environment Protection Key Laboratory of Wetland Ecology and Vegetation Restoration, School of Environment, Northeastern Normal University, Changchun 130117, China;
3. High School Attached to Northeast Normal University, Changchun 130022, China;
4. Institute of Grassland Science, School of Life Science, Northeast Normal University, Changchun 130024, China
菌根是植物根系和丛枝菌根真菌(Arbuscular Mycorrhizal Fungi,AMF)形成的共生体,是生物界最为广泛的一种互惠共生现象[1]。借助于AM真菌,宿主植物可在恶劣环境中获得良好的生长效应[2-3]。大量农作物和林木研究表明,AM真菌能够提高植物的耐盐性[4-5],增加作物在盐渍条件下的产量[6],并且筛选出了促进植物在盐渍土壤中生长的高效菌种[7]。随着菌根技术的不断发展,该研究和应用逐渐由农业和林业扩展到草业[8]。AM真菌-植物共生体在逆境环境中表现出来的互惠效应,为治理和恢复退化草地生态系统提供了新的研究途径和方向[9]。
目前,全球范围内共有约9.32亿hm2土地遭受盐化和碱化,这些地区的植被恢复与当地的食品安全、环境健康和经济发展密切相关[10]。作为松嫩草甸草原的优势物种和优良牧草,羊草(Leymus chinensis)与AM真菌共生体的特性和互惠效应在治理大面积盐碱化草地的恢复工作中备受关注。已有研究表明,羊草能与多种AM真菌在自然条件下形成共生体[11-12],并且提高了羊草在重度盐碱化草地上的定居率[9]。但当盐胁迫压力加大时,高盐环境不仅抑制植物生长,同时还阻碍了AM真菌的孢子萌发和菌丝生长[13]。此外,双方对共同资源的竞争可能会加剧降低AM真菌对羊草耐盐性的促进作用,甚至转变为单方或双方受抑[14-15]。因此,了解不同强度盐胁迫下AM真菌对羊草生长的影响,是应用菌根技术治理盐化羊草草地的基础。本研究设置了不同浓度的NaCl盐处理,研究不同强度盐胁迫对AM真菌的侵染能力和羊草菌根效应的影响,并从植物形态和离子含量角度探讨菌根化羊草对盐胁迫的响应机理。研究结果对于应用菌根技术治理和修复盐碱退化羊草草地等实践活动具有理论指导意义。
1 材料与方法 1.1 实验材料供试土壤取自温室周围的农田土。农田土的水溶性无机盐含量为0.81 g/kg,土壤pH为8.49,有机质含量为2.49%,全氮、全磷、全钾分别为1.69、0.91、34.8 g/kg,速效氮、速效磷、速效钾分别为108.65、8.76、23.5 mg/kg。为改善土壤质地,将农田土过2 mm网筛后按1∶3的河沙∶农田土比例混合。将土壤河沙混合均匀后放入高压蒸汽灭菌锅中121 ℃灭菌2h,以消除土壤和河沙中的真菌孢子和其它土壤微生物。
羊草种子来自吉林绿源草业公司的羊草种子田。播种前将羊草种子用10 % H2O2浸泡消毒10min后,放入蒸馏水中浸泡2h,然后取出放置于湿润滤纸上48h催芽备用。
AM真菌菌种取自吉林省西部盐碱化羊草草地。经鉴定,该土壤中的主要菌种为球囊霉属(Glomus)的摩西球囊霉(G. mosseae)和地球囊霉(G. geosporum)[16]。将上述两个菌种在羊草根系上进行5个月的诱集培养,然后将包括菌根真菌菌丝、孢子、侵染根段及含根外菌丝体的根际土壤的混合物作为AM真菌根接种剂。并将一半接种剂进行灭菌处理,方法同土壤灭菌。
1.2 实验设置本研究于2005年6—8月在吉林省林业厅种苗基地温室中进行。实验期间室温20—25 ℃,日光照射,光照时间控制为8:00—16:00的8h。本研究包含NaCl盐和AM真菌接种两个试验因素,分别设置0、1、2、3 g NaCl / kg土的4个盐处理水平,接种(+AM)和不接种(-AM)2个AM真菌处理水平,共计8个处理。每个处理4盆重复,共32盆。
塑料花盆的上口内径为15 cm,盆底内径为12 cm,盆高15 cm,盆底无孔。使用前将花盆用10 % H2O2浸泡消毒15min。每盆装入2 kg已消毒的农田土-河沙混合物。按照设计的盐处理水平,将相应重量的NaCl溶于蒸馏水中,并与花盆中的土壤充分混匀。在+AM处理的花盆中放入30 g菌根接种剂,将其放置在种子下2 cm深的土层中。在-AM处理的花盆中将30 g灭菌的接种剂放入到相同土层中,并放入30 mL不灭菌接种剂的水滤液(滤膜孔径0.25 μm),以保持+AM和-AM处理除AM真菌外一致的微生物区系。
在花盆中均匀播种羊草种子,出苗后间苗,每盆留间隔均匀的50株。生长期间每周浇2次营养液,每次50 ml。营养液含N100 mg/L,K、P、Mg、Zn 各50 mg/L。实验期间维持正常水分状态。
1.3 样品采集和测量幼苗生长75 a后,齐地面收获羊草地上部分,并挑出根系。用自来水冲洗地上部分和根系后,再用去离子水冲洗3—4遍,用滤纸吸干表面水分。记录根系总鲜重。随机选取一部分根系,记录鲜重后,用于后续的菌根侵染测定。将羊草地上部分和剩余根系放入65 ℃下烘至恒重并称重。
采用KOH透明-乳酸甘油酸性品红染色法染色。将冲洗干净的根系切成1cm左右长度的根段,将根段放入10 % KOH溶液中,水浴(90 ℃)60min,蒸馏水冲洗2次。随后放入碱性双氧水中软化20min,水洗后在2 %的盐酸中酸化3—4min。然后在90 ℃水浴锅中用酸性品红染色30min。取出根段后放入乳酸甘油(1∶1)中浸泡脱色。随机选取50条根段压片,在显微镜下观察每条根的侵染长度(以mm记录),并记录被侵染的根系数量。
将羊草地上部分粉碎过40目筛。测量羊草地上部分的N、P、K+、Na+、Ca2+、Cl-元素的浓度。采用凯氏测氮仪(Kjeldahl,Foss Tractor)测N,紫外分光光度计(UV2201)测P(NaHCO3浸提-钼锑抗比色法),原子吸收分光光度计(AA-220 FS)测K+、Ca2+、Na+,离子色谱仪(DX-300)测Cl-。
1.4 数据处理和统计分析依据下列公式计算AM真菌对羊草根系的侵染情况[17-18]:
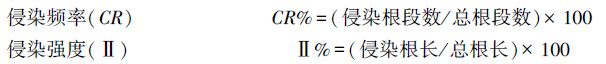
以羊草的单位面积生物量为基础,按下列公式计算每个盐处理的盐胁迫效应(SR)、菌根生长效应(MGR)、磷依赖性(MPR)和钾依赖性(MKR)[19]:


使用混合模型(Mixed Models)分别检验NaCl和AM真菌因素对各响应变量的影响,以及两者是否存在交互作用。以重复盆作为随机因素纳入模型中。对-AM和+AM处理,使用t检验进行均值比较。显著水平均为P < 0.05。本实验数据分析采用SPSS 19.0软件。
2 结果 2.1 羊草生物量、根茎比和盐胁迫效应盐和菌根处理对羊草总生物量影响显著,但两者的交互作用不明显(表 1)。随NaCl浓度的增加,菌根化羊草生物量与非菌根化羊草的差异增加,至3 mg NaCl/kg水平时差异达到显著水平(图 1)。盐对羊草根茎比的影响不显著,虽然菌根化羊草的根茎比随NaCl浓度的增加而降低,但方差检验未达到显著水平(F=2.704,P=0.092)(表 1,图 1)。菌根化羊草的根茎比显著高于非菌根羊草,但随着盐胁迫的增加,这种差异逐渐减小(图 1)。
| 指标Index | 盐NaCl | 菌根AM | 盐×菌根NaCl×AM |
| 总生物量Total Biomass | 12.72(3,21)*** | 10.54(1,21)** | 1.25(3,21) |
| 根茎比Root/shoot | 2.13(3,21) | 63.02(1,21)*** | 2.54(3,21) |
| 盐胁迫效应Salinity response | 7.30(2,15)** | 7.28(1,15)* | 0.93(2,15) |

|
| 图 1 盐对菌根化(+AM)和非菌根化(-AM)羊草的生物量和根茎比的影响及盐胁迫效应 Fig. 1 The biomass and root/shoot ratio of mycorrhizal (+AM) and non-mycorrhizal (-AM) L. chinensis under different salt treatment and the salinity response |
随着盐浓度的增加,盐胁迫效应随之增加,在3 mg NaCl/kg水平下,盐胁迫效应近40 %(表 1,图 1)。AM真菌降低了羊草的盐胁迫效应,并在高盐条件下达到显著水平(图 1)。
2.2 羊草的侵染频率、侵染强度和菌根生长效应盐胁迫显著影响AM真菌对羊草根系的侵染频率(F=11.12,P= 0.007)和侵染强度(F=6.72,P=0.014),尤其是侵染强度随盐浓度的增加而持续降低(图 2)。羊草菌根生长效应在无盐条件下近于零,但随NaCl浓度的增加而显著增加(F=4.19,P = 0.041,图 2)。

|
| 图 2 不同盐胁迫条件下AM真菌侵染情况和羊草的菌根生长效应 Fig. 2 The colonization rate,infection intensity and mycorrhizal growth respone of mycorrhizal L. chinensis under different salt stress |
盐和菌根处理对羊草N、P浓度的影响达到极显著水平,并且存在显著的交互作用(表 2)。菌根化羊草的N、P浓度始终高于非菌根化羊草,但这种差异随着盐胁迫的增加而不断减少,至3 mg/kg高盐时,-AM和+AM羊草的P浓度差异不显著(图 3)。AM真菌促进羊草累积更多N、P(表 2,图 4),但盐胁迫对羊草N累积量的影响不显著,却显著降低了P累积量(表 2)。
| 指标 Index | 离子 Ion | 盐 NaCl | 菌根 AM | 盐×菌根 NaCl×AM |
| 离子浓度Ionic content | N | 26.39(3,21)*** | 477.53(1,21)*** | 9.45(3,21)*** |
| P | 18.05(3,21)*** | 171.30(1,21)*** | 29.42(3,21)*** | |
| K+ | 141.76(3,21)*** | 1.37(1,21) | 15.71(3,21)*** | |
| Ca2+ | 68.90(3,21)*** | 7.18(1,21)* | 57.19(3,21)** | |
| Na+ | 5596.17(3,21)*** | 702.99(1,21)*** | 54.82(3,21)*** | |
| Cl- | 3060.66(3,21)*** | 212.07(1,21)*** | 61.67(3,21)*** | |
| 离子累积量Ionic amount | N | 2.84(3,21) | 142.81(1,21)*** | 0.42(3,21) |
| P | 9.58(3,21)*** | 33.17(1,21)*** | 0.70(3,21) | |
| K+ | 13.70(3,21)*** | 7.26(1,21)* | 6.43(3,21)** | |
| Ca2+ | 26.92(3,21)*** | 18.05(1,21)*** | 1.17(3,21) | |
| Na+ | 6.36(3,21)** | 2.11(1,21) | 2.81(3,21) | |
| Cl- | 16.10(3,21)*** | 2.28(1,21) | 3.65(3,21)* | |
| 离子比 | K+/Na+ | 253.26(3,21)*** | 15.32(1,21)*** | 12.66(3,21)*** |
| P/Na+ | 769.63(3,21)*** | 592.68(1,21)*** | 48.18(3,21)*** |

|
| 图 3 不同盐条件下菌根化(+AM)和非菌根化(-AM)羊草的离子浓度含量 Fig. 3 The ionic contents of mycorrhizal (+AM) and non-mycorrhizal (-AM) L. chinensis under different salt treatments |

|
| 图 4 不同盐条件下菌根化(+AM)和非菌根化(-AM)羊草的离子累积量 Fig. 4 The ionic amounts of mycorrhizal (+AM) and non-mycorrhizal (-AM) L. chinensis under different salt treatments |
方差分析显示AM真菌对羊草K+浓度影响不显著,但NaCl和AM真菌的交互作用表明,在不同盐水平下,AM真菌对羊草K+浓度的影响不同(表 2)。在无盐条件下,菌根化羊草K+离子浓度显著低于非菌根化羊草,但在2 mg/kg盐条件下,菌根化羊草K+离子浓度却显著高于非菌根化羊草(图 3),并且在高盐条件下K+累积量显著高于非菌根化羊草(图 4)。
AM真菌对羊草Ca2+浓度的作用受盐胁迫强度的影响(表 2)。在0和1 mg/kg盐水平下,菌根化羊草Ca2+浓度高于非菌根化羊草,但在2和3 mg/kg盐水平下,显著低于非菌根化羊草(图 3)。无论是菌根化羊草还是非菌根化羊草,Ca2+累积量随盐浓度的增加而降低。在0和3 mg NaCl/kg条件下,菌根化羊草吸收累积的Ca2+离子高于非菌根化羊草(图 4)。
无论是菌根化还是非菌根化,羊草Na+离子浓度随盐浓度的增加而增加。并且菌根化羊草Na+浓度一般低于非菌根化羊草(图 3)。在无盐条件下,菌根化羊草钠累积量低于非菌根化羊草,而在3 mg NaCl/kg高盐条件下,菌根化羊草吸收累积的Na2+离子量高于非菌根羊化草(图 4)。
无论是Cl-浓度还是Cl-积累量,均随着盐浓度的增加而逐渐增加。除2 mg/kg处理外,非菌根化羊草的Cl-浓度显著高于菌根化羊草(图 3)。与非菌根化羊草相比,菌根化羊草在无盐条件下累积较少的氯离子,但在高盐条件下吸收累积较多氯离子(图 4)。
无论是K+/Na+还是P/Na+,均在有盐条件下低于无盐条件下(表 2,图 5)。AM真菌对这两个指标的影响在不同盐水平下表现不一。在无盐和低盐条件下,有无菌根对羊草的K+/Na+没有显著影响,但在2、3 mg/kg高盐水平下,AM真菌显著提高了羊草的K+/Na+(图 5)。P/Na+与此相反,在无盐和低盐条件下,AM真菌显著提高了羊草的P/Na+,但在2、3 mg /kg高盐水平下有无菌根对该指标无显著影响(图 5)。

|
| 图 5 不同盐条件下菌根化和非菌根化羊草的钾钠比和磷钠比 Fig. 5 The K+/Na+ and P/Na+ ratios of mycorrhizal and non-mycorrhizal L. chinensis under different salt stress |
磷依赖性指标在所有盐处理中表现为正值,表明无论土壤中有无NaCl盐,菌根化羊草对磷的依赖性始终存在。但在低盐条件下(1 mg/kg)菌根化羊草对磷的依赖性最强,随着盐胁迫的不断增加,磷依赖性随之下降(图 6)。菌根化羊草仅在2、3 mg/kg高盐水平条件下表现出钾依赖性(图 6)。

|
| 图 6 不同盐胁迫条件下菌根化羊草的磷依赖性和钾依赖性 Fig. 6 The mycorrhizal P response and mycorrhizal K response of L. chinensis under different salt stress |
一般而言,丛枝菌根真菌侵染可改善植物的生长状况,但改善程度取决于土壤性质[2, 20]。当生长环境良好,植物未受到胁迫或胁迫较低时,AM真菌与植物的互利现象并不明显。如本研究中的无盐和低盐处理,AM真菌对羊草生物量的促进作用并未达到显著水平,即羊草的菌根生长效应并不明显。当然,该研究结果也可能由于数据来源自羊草生物量积累较为缓慢的幼苗阶段,AM真菌的菌根生长效应并为完全发挥出来。但即使是在幼苗阶段,当羊草植株面临较高的盐胁迫时,AM真菌显著降低了NaCl对羊草产生的盐胁迫效应,这充分证明了AM真菌-植物共生体对植物在逆境中生存的重要性。
但当逆境中的胁迫强度加大时,不仅植物生长受到抑制,同时还阻碍了AM真菌的孢子萌发和菌丝生长[13]。虽然本研究使用的摩西球囊霉(G. mosseae)和地球囊霉(G. geosporum)是重度盐土中最优势的AM真菌物种[4],但在高钠盐胁迫下,其孢子萌发和菌丝生长也会受到抑制[21]。这不仅降低了AM真菌对羊草根系的侵染能力,并且有可能导致AM真菌和羊草为争夺有限资源而形成相互抑制的竞争关系。虽然羊草的菌根生长效应并没有随盐分浓度的增加而下降,但AM真菌对羊草根系侵染频率和侵染强度却随盐分的增加而显著下降。因此,随着胁迫继续加剧,AM真菌对羊草耐盐性的促进作用将会降低,甚至转变为单方或双方受抑[14-15]。事实上,在环境恶劣的生态系统中,AM真菌和植物共生体面临着多种影响其互利关系的生物和非生物因素,其综合作用可能会加速削弱AM真菌对植物生长的促进作用。
3.2 菌根化羊草的根茎比增加一些研究证明AM真菌能够增强植物的存活率和生物量[22-23]。在无盐或盐胁迫较低时,AM真菌并没有显著改变羊草的生物量,却极大改变了羊草生物量的地上地下分配比例,并且在所有盐处理中,菌根化植物一直保持较高的根茎比。有研究表明增加的根生长能够促进水分和营养的吸收[24-26]。菌根化羊草将更多的光合产物分配到根系,为植株的后期生长发育打下了坚实的基础和保障。相对增加的根系将比无菌根化羊草植株具有更强的水分和营养元素吸收和传输能力,从而能为菌根化植株拓展更为广阔的资源空间和生存机会。因此,菌根化植株重新分配根茎比可能是AM真菌促进植物生长的一条重要作用途径。
3.3 菌根化羊草的离子含量和离子比发生变化除形态变化外,菌根化植物的营养元素含量也发生变化,从而增强了其生长能力[27]。在盐胁迫条件下,因土壤中盐离子过多而排斥植物对其他离子的吸收,导致植物营养失调,抑制生长。一些研究表明,接种AM真菌能够提高植株的营养元素含量[27-28]。本实验中的菌根化羊草一直维持较高的N、P浓度,但随着盐胁迫的加大,该优势逐渐减少。一方面可能由于盐分增加抑制了AM真菌对羊草根系的侵染能力,从而降低其营养吸收能力;另一方面可能由于AM真菌在恶劣环境里与寄主植物之间的营养竞争较激烈,从而减少了寄主一方的利益。虽然高盐条件下羊草P浓度无差异,但在整个实验过程中,菌根化羊草比非菌根化羊草吸收积累了更多的P元素,证明了AM真菌能够促进植物吸收利用更多的N、P元素。
AM真菌有效减轻了盐胁迫条件下的离子毒害作用[29]。菌根化羊草一直维持着较低的Na+和Cl-离子浓度。有关AMF减缓盐离子对植物毒害作用的机理还未有一致的结论。可能是由于菌根化植株的P、K+和Ca2+等离子含量增加,不仅降低了细胞渗透势,还有效降低了盐离子的相对含量,从而减轻盐离子的毒害作用[30]。也可能由于AM真菌促进植株生长,由此相对稀释了有毒离子的浓度,减缓了离子毒害[13]。
盐胁迫对植物的危害还表现在渗透势增加所导致的生理缺水[31]。菌根化植物除了大量吸收外界营养元素促进生长外,还大量吸收一些其它离子来调节渗透势,诸如K+,从而提高植物的耐盐能力[32]。低盐条件下菌根化羊草的Ca2+离子浓度和中高盐条件下的K+离子浓度均高于非菌根化羊草。Ca2+ 、K+离子浓度的增加有效降低了细胞渗透势,其中K+在高盐胁迫环境下对改善细胞渗透势更为重要。此外,低盐条件下菌根化羊草的P/Na+比,和中高盐条件下的K+/Na+比均高于非菌根化羊草。因此,维持较高的P/Na+、K+/Na+可能也是AM真菌增强羊草耐盐性的机制之一[33]。随着盐胁迫的增加,菌根化羊草对磷的依赖性逐渐转换为对钾的依赖性,羊草的抗盐能力随着盐胁迫的增加越来越依赖于钾元素。
| [1] | Smith S E, Facelli E, Pope S, Smith A F. Plant performance in stressful environments:interpreting new and established knowledge of the roles of arbuscular mycorrhizas. Plant and Soil , 2010, 326 (1/2) : 3–20. |
| [2] | Evelin H, Kapoor R, Giri B. Arbuscular mycorrhizal fungi in alleviation of salt stress:a review. Annals of Botany , 2009, 104 (7) : 1263–1280. DOI:10.1093/aob/mcp251 |
| [3] | 李晓林, 冯固. 丛枝菌根生态生理. 北京: 华文出版社, 2001. |
| [4] | Bothe H. Arbuscular mycorrhiza and salt tolerance of plants. Symbiosis , 2012, 58 (1/3) : 7–16. |
| [5] | Talaat N B, Shawky B T. Protective effects of arbuscular mycorrhizal fungi on wheat (Triticum aestivum L.) plants exposed to salinity. Environmental and Experimental Botany , 2014, 98 : 20–31. DOI:10.1016/j.envexpbot.2013.10.005 |
| [6] | Talaat N B, Shawky B T. Influence of arbuscular mycorrhizae on root colonization,growth and productivity of two wheat cultivars under salt stress. Archives of Agronomy and Soil Science , 2012, 58 (1) : 85–100. DOI:10.1080/03650340.2010.506481 |
| [7] | Estrada B, Aroca R, Barea J M, Ruiz-Lozano J M. Native arbuscular mycorrhizal fungi isolated from a saline habitat improved maize antioxidant systems and plant tolerance to salinity. Plant Science , 2013, 201-202 : 42–51. DOI:10.1016/j.plantsci.2012.11.009 |
| [8] | Karanika E D, Mamolos A P, Alifragis D A, Kalburtji K L, Veresoglou D S. Arbuscular mycorrhizas contribution to nutrition,productivity,structure and diversity of plant community in mountainous herbaceous grassland of northern Greece. Plant Ecology , 2008, 199 (2) : 225–234. DOI:10.1007/s11258-008-9427-9 |
| [9] | Zhang Y F, Wang P, Yang Y F, Bi Q, Tian S Y, Shi X W. Arbuscular mycorrhizal fungi improve reestablishment of Leymus chinensis in bare saline-alkaline soil:Implication on vegetation restoration of extremely degraded land. Journal of Arid Environments , 2011, 75 (9) : 773–778. DOI:10.1016/j.jaridenv.2011.04.008 |
| [10] | Rengasamy P. World salinization with emphasis on Australia. Journal of Experimental Botany , 2006, 57 (5) : 1017–1023. DOI:10.1093/jxb/erj108 |
| [11] | Carvalho L M, Caçador I, Martins-Loução M. Temporal and spatial variation of arbuscular mycorrhizas in salt marsh plants of the Tagus estuary (Portugal). Mycorrhiza , 2001, 11 (6) : 303–309. DOI:10.1007/s00572-001-0137-6 |
| [12] | 张伟溪, 王洪峰, 赵昕, 阎秀峰. 松嫩盐碱草地植物丛枝菌根的初步调查. 土壤通报 , 2010, 41 (6) : 1380–1385. |
| [13] | Juniper S, Abbott L. Vesicular-arbuscular mycorrhizas and soil salinity. Mycorrhiza , 1993, 4 (2) : 45–57. DOI:10.1007/BF00204058 |
| [14] | Johnson N C, Graham J H, Smith F A. Functioning of mycorrhizal associations along the mutualism-parasitism continuum. New Phytologist , 1997, 135 (4) : 575–586. DOI:10.1046/j.1469-8137.1997.00729.x |
| [15] | Karst J, Marczak L, Jones M D, Turkington R. The mutualism-parasitism continuum in ectomycorrhizas:A quantitative assessment using meta-analysis. Ecology , 2008, 89 (4) : 1032–1042. DOI:10.1890/07-0823.1 |
| [16] | 张义飞. 松嫩平原盐碱化草地丛枝菌根真菌资源及其生态作用的研究[D]. 长春:东北师范大学,2011. http://cdmd.cnki.com.cn/article/cdmd-10200-1012291123.htm |
| [17] | McGonigle T P, Miller M H, Evans D G, Fairchild G L, Swan J A. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytologist , 1990, 115 (3) : 495–501. DOI:10.1111/nph.1990.115.issue-3 |
| [18] | Biermann B, Linderman R G. Quantifying vesicular-arbuscular mycorrhizae:a proposed method towards standardization. New Phytologist , 1981, 87 (1) : 63–67. DOI:10.1111/nph.1981.87.issue-1 |
| [19] | Colla G, Rouphael Y, Cardarelli M, Tullio M, Rivera C M, Rea E. Alleviation of salt stress by arbuscular mycorrhizal in zucchini plants grown at low and high phosphorus concentration. Biology and Fertility of Soils , 2008, 44 (3) : 501–509. DOI:10.1007/s00374-007-0232-8 |
| [20] | 江龙, 王茂胜, 黄建国, 袁玲. 丛枝菌根真菌对烟苗吸收基质养分的影响. 贵州农业科学 , 2010, 38 (6) : 50–54. |
| [21] | Carvalho L M, Correia P M, Caçador I, Martins-Loução M A. Effects of salinity and flooding on the infectivity of salt marsh arbuscular mycorrhizal fungi in Aster tripolium L.. Biology and Fertility of Soils , 2003, 38 (3) : 137–143. DOI:10.1007/s00374-003-0621-6 |
| [22] | Kaya C, Ashraf M, Sonmez O, Aydemir S, Tuna A L, Cullu M A. The influence of arbuscular mycorrhizal colonisation on key growth parameters and fruit yield of pepper plants grown at high salinity. Scientia Horticulturae , 2009, 121 (1) : 1–6. DOI:10.1016/j.scienta.2009.01.001 |
| [23] | Sheng M, Tang M, Chen H, Yang B W, Zhang F F, Huang Y H. Influence of arbuscular mycorrhizae on the root system of maize plants under salt stress. Canadian Journal of Microbiology , 2009, 55 (7) : 879–886. DOI:10.1139/W09-031 |
| [24] | Müller I, Schmid B, Weiner J. The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plants. Perspectives in Plant Ecology,Evolution and Systematics , 2000, 3 (2) : 115–127. DOI:10.1078/1433-8319-00007 |
| [25] | Eissenstat D M, Graham J H, Syvertsen J P, Drouillard D L. Carbon economy of sour orange in relation to mycorrhizal colonization and phosphorus status. Annals of Botany , 1993, 71 (1) : 1–10. DOI:10.1006/anbo.1993.1001 |
| [26] | Johnson D, Leake J R, Ostle N, Ineson P, Read D J. In situ 13CO2 pulse-labelling of upland grassland demonstrates a rapid pathway of carbon flux from arbuscular mycorrhizal mycelia to the soil. New Phytologist , 2002, 153 (2) : 327–334. DOI:10.1046/j.0028-646X.2001.00316.x |
| [27] | Rabie G H. Influence of arbuscular mycorrhizal fungi and kinetin on the response of mungbean plants to irrigation with seawater. Mycorrhiza , 2005, 15 (3) : 225–230. DOI:10.1007/s00572-004-0345-y |
| [28] | Giri B, Kapoor R, Mukerji K G. Improved tolerance of Acacia nilotica to salt stress by arbuscular mycorrhiza,Glomus fasciculatum may be partly related to elevated K/Na ratios in root and shoot tissues. Microbial Ecology , 2007, 54 (4) : 753–760. DOI:10.1007/s00248-007-9239-9 |
| [29] | Garg N, Chandel S. Role of arbuscular mycorrhizal (AM) fungi on growth,cadmium uptake,osmolyte,and phytochelatin synthesis in Cajanus cajan (L.) millsp. under NaCl and Cd stresses. Journal of Plant Growth Regulation , 2012, 31 (3) : 292–308. DOI:10.1007/s00344-011-9239-3 |
| [30] | 冯固, 张福锁. 丛枝菌根真菌对棉花耐盐性的影响研究. 中国生态农业学报 , 2003, 11 (2) : 21–24. |
| [31] | 冯固, 李晓林, 张福锁, 李生秀. 盐胁迫下丛枝菌根真菌对玉米水分和养分状况的影响. 应用生态学报 , 2000, 11 (4) : 595–598. |
| [32] | 贺忠群, 贺超兴, 张志斌, 邹志荣. 丛枝菌根真菌提高植物耐盐性的作用机制. 西北植物学报 , 2007, 27 (2) : 414–420. |
| [33] | 冯固, 杨茂秋, 白灯莎. 盐胁迫下VA菌根真菌对无芒雀麦体内矿质元素含量及组成的影响. 草业学报 , 1998, 7 (3) : 21–28. |