 2016, Vol. 36
2016, Vol. 36文章信息
- 张宪德, 徐婷婷, 赵婷婷, 万冬梅, 殷江霞
- ZHANG Xiande, XU Tingting, ZHAO Tingting, WAN Dongmei, YIN Jiangxia.
- 配偶选择中“听众效应”及鸣唱对雌性灰文鸟行为的影响
- Effects of “audience effects” and song on female Padda oryzivora mate choice behavior
- 生态学报[J]. 2016, 36(16): 5247-5255
- Acta Ecologica Sinica[J]. 2016, 36(16): 5247-5255
- http://dx.doi.org/10.5846/stxb201501290238
-
文章历史
- 收稿日期: 2015-01-29
- 网络出版日期: 2015-11-30

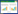




繁殖行为是鸟类行为的重要组成部分[1],而配偶选择是繁殖行为研究的重要内容[2],是性选择的重要方式[3],更是性选择研究的中心问题[4]。配偶选择是一性个体对另一性个体的主动选择,大多数由雌性主导[3],并且具有较强的可塑性以适应外界环境和内部条件的变化。雌性配偶选择能够影响进化的强度和方向[5],有利于雌性获得一些直接利益,如获得所选雄性提供的一些资源、营养、保护或在后代抚育中提供的帮助,并且可以通过遗传间接为后代获取利益[6]。 雌性通常会选择高质量的雄性进行交配,以获得更好的基因或具有吸引力的后代[7]。它们会根据不同的信号对雄性质量进行评估,包括鸣声、行为表现、信息素和形态学特征[8]。在雌性选择为主导的配偶选择中,雄鸟会通过鸣声或其他信息吸引雌性,雌性根据信息对雄鸟质量做出评价,最终选择高质量的雄性作为配偶[9]。鸣禽中,雄鸟鸣声在吸引和刺激雌鸟中发挥重要作用[10]。许多鸟类偏爱较复杂鸣声[10],如水蒲苇莺(Acrocephalus schoenobaenus)[11],北美歌雀(Melospiza melodia)[12]等;有些雌性偏爱较大的鸣声输出,如欧洲椋鸟(Sturnus vulgaris)[13]、白喉带鹀(Zonotrichia albicollis)[14]等。鸣声是一种求爱信号,能够向雌性成鸟传递求爱信息[15],具有物种特异性和个体差异[16-18],为其用于物种识别、区分个体提供了基础[19],是许多鸟类选择同种个体作为配偶的重要依据,对于鸟类物种的形成具有重要意义[20-22]。与此同时,鸣声也是雄性质量的可靠指示器,有利于雌性选择优秀的雄性作为配偶[23-24],提高繁殖成功率并获得优秀的子代,有利于种族的进化和延续[3]。
大多数动物生活在一个信息网络中[25],个体可以通过“偷听”获得有用的信息,如竞争对手和潜在配偶的活力、地位和品质等[25-26]。在许多动物中,雄性都有通过“偷听”从雄性-雄性竞争行为中收集信息的现象,如新疆歌鸲(Luscinia megarhynchos)[27-28]、泰国斗鱼(Betta splendens)[29]、大山雀(Parus major)[30]、金丝雀(Serinus canaria)[31-32]等。雌性也会在早期配偶选择中通过“偷听”对潜在配偶进行评估选择,如泰国斗鱼[33],日本鹌鹑(Coturnix Japonica)[34-35],金丝雀[36-37]等。在婚外交配尝试中也有这一行为,如大山雀[38],黑冕山雀(Poecile atricapilla)[39]等。在“偷听”行为发生的过程中,被偷听者也会由于听众的存在而调节自身信息的行为,即“听众效应(audience effects)”[40-42]。“听众效应”能够隐藏[40]或突出[43]被偷听者的某些信息,或降低信息的可信度[44]。例如在对雄性家养金丝雀在有雌性“听众效应”下的婚外行为和同性攻击行为的研究中发现,雄性在“听众效应”下会隐藏他们的婚外行为,但会增强他们的同性竞争行为[45],充分说明个体会在第三方偷听的条件下适当调节自身的行为。 目前,绝大多数对“听众效应”的研究主要集中于雌性配偶选择中雌性“听众效应”对雄-雄竞争行为的影响,而雄性“听众效应”对雌-雌竞争行为的研究尚未开展。雌性配偶选择研究对于研究动物的性选择机制[6]和进化过程均具有重要意义[3, 6]。以灰文鸟(Padda oryzivora)为研究对象,从雄性鸣唱及雄性“听众效应”对雌性配偶选择的影响两个方面展开研究。灰文鸟,雀形目,文鸟科,是一种常见观赏鸟类,容易购买且实验成本较低,适应性强极易饲养,适合实验室养殖。雄鸟叫声尖而长,雌鸟叫声短促,声音采集较为方便。灰文鸟繁殖周期短,且在北方地区,秋、冬、春三季均可繁殖,产卵频率高,育雏时间短,适宜作为模式物种进行实验研究[46-48]。
1 研究对象和研究方法 1.1 研究对象将10只没有亲缘关系的成年雄性灰文鸟与10只成年雌性灰文鸟随机配对,所有个体均在实验室孵化成长,具有繁殖经验,且之前从未进行过此类实验[45]。每对鸟饲养在一个笼子(35cm×30cm×25cm)中,笼子中有供鸟自由采食的食槽和水槽,活动跳跃的塑料栖木,繁殖产卵的草编鸟巢。首先将10个鸟笼放置于同一房间中饲养7d,相互之间能够看到和听到,使其相互熟悉,以避免雌鸟对配偶和其他雄鸟熟悉程度的不同对实验的影响,之后在15∶9(光照∶黑暗)的光照周期[49]下喂食灰文鸟发情饲料(鸡蛋米)刺激其繁殖[47],使灰文鸟经历第1次繁育周期以巩固配偶关系。第1次繁育周期过程中以塑料模型替换产下的卵,直到最后一枚卵产下一周后将巢移走[45],这样可以使所有鸟共同开始第2次繁育周期,以避免第2次繁育周期开始时间不同可能对性选择偏爱造成影响[50]。待灰文鸟开始第2次繁育时,即可开始实验。
1.2 实验一 雄鸟鸣唱对雌鸟配偶选择的影响在饲养环境下,使用HYUNDAI便携式录音笔对实验所用到的10只雄鸟进行了鸣声采集,以雄鸟做出求偶炫耀动作,并且雌鸟做出CSD反应时的鸣唱作为求偶鸣唱,反之则为非求偶鸣唱。非求偶鸣唱句子时长(4.63±1.64)s(n=10),求偶鸣唱句子时长(7.75±2.06)s(n=10)。以格式工厂软件对录音进行了筛选、音量调节等处理后的录音作为播放录音。实验时将雌鸟置于双向选择实验装置中,类似Collins等在研究中使用的装置[51]。该实验装置是一个(15cm×90cm×15cm)的长方体实验笼,笼子两端及中心区域均有食槽和水槽,两端各有一个播放器(图 1)。由于上午是鸟类鸣唱的高峰期,所以实验每天从08:00开始。实验开始后先将雌鸟置于装置中10 min熟悉环境,之后两端同时以先科多功能扩音器12 min内无间隔连续播放录音,其中一端播放其配偶鸣唱(求偶鸣唱或非求偶鸣唱),另一端播放非配偶鸣唱(求偶鸣唱或非求偶鸣唱)。为避免雌性偏侧效应,每种播放组合在播放完毕后需颠倒播放并记录,即按照随机顺序在实验装置两端按以下组合方式均衡不同的鸣唱声音:

|
| 图 1 实验一中使用的双向选择装置 Fig. 1 Two-way choice device used in experiment 1 |
(1) 灰文鸟雌鸟配偶的非求偶鸣唱与熟悉雄鸟的非求偶鸣唱;(2)配偶的非求偶鸣唱与熟悉雄鸟的求偶鸣唱;(3)配偶的求偶鸣唱与熟悉雄鸟的非求偶鸣唱;(4)配偶的求偶鸣唱与熟悉雄鸟的求偶鸣唱。
以雌鸟位置偏向一侧并且面向这一侧定义为积极偏爱[52]。播放开始后,给雌鸟2 min时间让其定位,随后观察者于距离装置1.5m的帷幕后[52]观察记录雌鸟的积极偏爱次数,15s记录1次,每组记录10 min。通过比较雌鸟C偏向A、B两个方向的积极偏爱次数,确定雌鸟的偏爱选择。
1.3 实验二 灰文鸟雌-雌竞争实验前对实验用鸟进行10h饥饿处理,将实验雌鸟和竞争雌鸟放置到一个(35cm×30cm×25cm)笼即实验笼中,中间以铁丝网隔开,以避免其身体接触。实验笼旁放置一听众笼(15cm×15cm×15cm),适应30 min后,撤去隔网,投食开始后记录。根据听众笼分为3个连续的实验阶段,即:(1)无听众,(2)一只熟悉雄性做听众(竞争对手的配偶),(3)雄性配偶做听众(竞争对手的熟悉雄性),每个阶段均持续10 min。实验中,允许实验笼和听众笼中鸟的视觉和听觉交流,通过记录不同听众笼条件下雌性的攻击行为次数来评估雌性的反映。攻击行为包括掠食尝试、防止掠食成功和反击追逐。为避免实验者偏向,观察者并不知道听众的身份直到数据被分析[45]。
1.4 数据处理实验一样本容量等于30和实验二样本容量大于30,均可近似按正态分布处理。实验一数据使用成对样本T检验进行统计分析;实验二数据以听众类别为自变量以攻击行为次数为因变量进行单因素方差分析(One-way ANOVA),主要比较各方的均值关系及差异显著性,统计性显著水平设为0. 05。以上分析均使用统计软件SPSS(version 17. 0)进行。
2 结果 2.1 实验一鸣唱对配偶选择的影响 2.1.1 雌性偏爱鸣唱的类型和来源雌性灰文鸟对配偶非求偶鸣唱的积极偏爱次数显著高于非配偶非求偶鸣唱(t=2.959,P<0.05;图 2),对配偶求偶鸣唱的积极偏爱次数显著高于非配偶非求偶鸣唱(t=7.681,P<0.05;图 2),对配偶求偶鸣唱的积极偏爱次数显著高于非配偶求偶鸣唱(t=4.799,P<0.05;图 2),对配偶非求偶鸣唱的积极偏爱次数显著低于非配偶求偶鸣唱(t=-3.192,P<0.05;图 2)。

|
| 图 2 雌性灰文鸟对不同类型的鸣唱的响应 (平均值±标准差) Fig. 2 The female preference of different types of song (mean±SD) |
雌性灰文鸟对配偶求偶和非求偶鸣唱的响应会受到环境中非配偶鸣唱类型的影响。主要表现在:当一端播放配偶求偶鸣唱而另外一端分别播放非配偶求偶鸣唱或非配偶非求偶鸣唱时,雌性灰文鸟对配偶求偶鸣唱的积极偏爱次数在播放非配偶求偶鸣唱时要显著低于播放非配偶非求偶鸣唱(t=-2.218,P<0.05;图 3);同样,当一端播放配偶非求偶鸣唱,而另外一端分别播放非配偶求偶鸣唱或非配偶非求偶鸣唱时,雌性灰文鸟对配偶非求偶鸣唱的积极偏爱次数在播放非配偶求偶鸣唱时要显著低于播放非配偶非求偶鸣唱(t=-3.645,P<0.05;图 3)。上述说明相比而言非配偶的求偶鸣唱比非求偶鸣唱更加具有吸引力,对雌鸟配偶选择影响更大。

|
| 图 3 非配偶鸣唱类型对灰文鸟对配偶鸣唱响应的影响(平均值±标准差) Fig. 3 The effects of different song types of other males on the responses of females to the songs of their spouse (mean±SD) |
雌鸟配偶选择的积极性以每种实验播放组合中雌性对装置A、B两端积极偏爱的总次数进行评估。当A、B两端均播放求偶鸣唱(配偶求偶鸣唱+非配偶求偶鸣唱)时雌性对两端鸣唱的总偏爱次数显著高于只有一端播放求偶鸣唱(配偶求偶鸣唱+非配偶非求偶鸣唱,t=2.636,P<0.05;配偶非求偶鸣唱+非配偶求偶鸣唱,t=4.049,P<0.05;)时的总偏爱次数;只有一端播放配偶求偶鸣唱(配偶求偶鸣唱+非配偶非求偶鸣唱)时雌性对两端鸣唱的总偏爱次数显著高于只有一端播放非配偶求偶鸣唱(配偶非求偶鸣唱+非配偶求偶鸣唱)时的总偏爱次数(t=2.437,P<0.05;)。当A、B两端均播放非求偶鸣唱(配偶非求偶鸣唱+非配偶非求偶鸣唱)时雌性对两端鸣唱的总偏爱次数最低,显著低于其他条件下(P<0.05)。
2.2 实验二:雄鸟“听众效应”对雌-雌竞争的影响应用单因素分析(One-way ANOVA)对不同听众条件下的雌-雌竞争进行了对比。分析发现:雌鸟的攻击行为受到听众条件的显著影响(F=11.72,P<0.001;图 4)。在配偶雄性作为听众条件下雌鸟攻击行为最高,显著高于熟悉雄性为听众的条件下,并且两者都显著高于无听众条件下雌鸟的攻击行为(图 4)。

|
| 图 4 不同听众类别条件下雌鸟所表现出的攻击行为(平均值±标准差) Fig. 4 Agonistic behaviours expressed by females according to the presented audience (mean±SD) |
本研究中,在配偶鸣唱和非配偶鸣唱之间,雌性灰文鸟更偏爱自身配偶的鸣唱,而非其他雄鸟的鸣唱,说明灰文鸟能够根据鸣唱识别雄性个体。许多研究已证明鸟类对同种鸣声具有分辨能力,如大山雀[53],虎皮鹦鹉(Melopsittacus undulatus)[54]和斑胸草雀(Poephila guttata)[55]等。鸣声结构的复杂性和多样性既具有种属特征,也存在普遍的个体差异[56],这种个体差异是雌性灰文鸟根据鸣唱识别个体的关键。本实验中,灰文鸟对配偶鸣唱的偏爱显著较高,一方面是由于配偶关系的存在,另一方面可能是由于配偶鸣唱对雌鸟的作用时间更长,雌鸟对配偶鸣唱的感知和分辨更加灵敏。
在配偶的非求偶鸣唱和非配偶的求偶鸣唱之间,雌性更偏爱非配偶的求偶鸣唱,可能是由于求偶鸣唱能够反映出雄鸟较高的质量[24]。相对于非求偶鸣唱而言,求偶鸣唱对雌性配偶选择的影响更大:非配偶的求偶鸣唱比非求偶鸣唱更能减弱雌性对配偶鸣唱的偏爱(图 3),说明求偶鸣唱比非求偶鸣唱更能吸引雌鸟,对雌性配偶选择的影响更大。Forstmeier等对褐柳莺(Phylloscopus fuscatus)的研究中发现褐柳莺对鸣曲的偏爱可能与雄性鸣唱的音节类型和鸣唱方式有关[57]。雌鸟更加偏爱于那些具有性感音节、鸣唱难度大的鸣唱。由于雄鸟求偶鸣唱和非求偶鸣唱担负着不同的功能,而求偶鸣唱的音节结构复杂,具有更多的性感音节,鸣唱难度更大,能够传递求爱信号,因而更能吸引雌性灰文鸟,对雌性配偶选择的影响更大;而配偶的非求偶鸣唱的音节结构简单,具有更少的性感音节,鸣唱难度较低,对雌性灰文鸟鸣唱偏爱和配偶选择的影响较小。这一研究结果说明,当鸣唱作为雌鸟评估雄鸟质量的标准时,拥有结构复杂的性感音节和鸣唱难度大的求偶鸣唱,更有可能成为雌鸟配偶选择和婚外尝试的诱因之一。
本研究还发现求偶鸣唱能够刺激雌性灰文鸟配偶选择的积极性,性选择中雄性通过竞争争夺配偶,雌性对雄性进行评估选择配偶,因此在雌性主导的配偶选择中雌性成为积极的一方[8]。不积极可能对雌鸟有利,因为这样可以减少寻找潜在配偶所投入的资源和时间[58],因而在两端均播放非求偶鸣唱时雌鸟显得最不积极,因为这些鸣唱不值得雌鸟花费时间评估,而在播放求偶鸣唱时雌鸟变得积极,这样有利于雌鸟选择优质的配偶。尽管这意味着要投入更多的资源和时间,但总体而言这对雌鸟是有利的。两端均播放求偶鸣唱时雌鸟比仅在一端播放求偶鸣唱时更加积极,是因为这时可供选择的鸣唱更多,雌鸟需要对两种鸣唱均进行评估,因而更加积极。我们还发现,雌性个体在只播放配偶求偶鸣唱时比只播放非配偶求偶鸣唱时更加积极,这可能是由于求偶行为能够刺激雌性性欲[59],而求偶鸣唱是求偶行为的重要组成,所以在求偶鸣唱条件下雌性交配欲望更加强烈。而配偶的求偶鸣唱比非配偶的求偶鸣唱更能刺激雌鸟交配欲望,因而表现得更加积极。
很多情况下配偶选择由雌性主导。雌性能够观察雄-雄竞争对配偶及婚外潜在配偶进行评估,雄性会根据雌性听众的不同适当调节自身行为,这种现象即为“听众效应”[45]。目前对“听众效应”的研究主要集中在雌性“听众效应”对配偶选择的影响,如Price等对孔雀鱼(Poecilia reticulata)[60]和Tachon等对黄斑黑蟋蟀(Gryllus bimaculatus)[61]的研究均发现雄性在雌性“听众效应”下攻击行为增强。而在鸟类中也有这种现象,如Baltz等对虎皮鹦鹉研究中发现雌性“听众效应”下雄性会收敛婚外行为[62],而Davy Ung等对金丝雀的研究中也发现在雌性“听众效应”条件下雄性会收敛婚外行为,雄-雄竞争行为增强[45]。本研究发现雌-雌竞争也受到“听众效应”的影响,在有听众条件下的攻击行为明显高于无听众条件的攻击行为;而在配偶听众条件下,雌性的攻击明显高于熟悉雄性听众条件下的攻击行为。这一发现与Davy Ung等对雌性“听众效应”对雄性家养金丝雀雄-雄竞争影响[45]的结果类似。雌性在雄性“听众效应”影响下雌-雌竞争行为明显增强,可能与雄-雄竞争配偶类似,雄性也可以通过偷听雌-雌竞争对雌鸟品质进行评估选择配偶。自然界条件下,配偶争夺多是雄性个体之间竞争雌性个体,同样雌性个体也会相互竞争争夺配偶[63],如斑腹矶鹬(Actitis macularia)[64-65],海马(Hippocampus)[66]等,从而与多个雄性交配中获得直接或间接的利益[67-68]。配偶雄性“听众效应”下的雌-雌竞争行为明显高于熟悉雄性“听众效应”下的雌-雌竞争行为,原因可能是配偶做听众条件下,如果竞争失败将付出更大的的代价,她们将失去与更多雄鸟的交配机会。鸟类在野生环境下的雌-雌竞争还有待于进一步的研究。
4 结论鸣唱在灰文鸟配偶选择中发挥着重要作用,雌性灰文鸟更加偏爱配偶鸣唱,说明求偶鸣唱和非求偶鸣唱均具有个体识别功能。但相对于配偶的非求偶鸣唱雌鸟更加偏爱于非配偶的求偶鸣唱,求偶鸣唱比非求偶鸣唱更能吸引雌性灰文鸟,说明求偶鸣唱具有吸引雌性的作用,而且这有可能会成为婚外行为发生的诱因之一。可供选择的求偶鸣唱更多时雌性变得更加积极,而求偶鸣唱能够刺激雌性灰文鸟配偶选择的积极性,可能与求偶鸣唱能够刺激雌鸟性欲有关。雌性灰文鸟会由于雄性“听众效应”对雌-雌竞争行为做出适当调整,但是否是由于雄鸟会根据雌-雌竞争对雌性灰文鸟进性评估进而选择交配对象,还需进一步研究。
| [1] | 郑光美, 张正旺. 鸟类生态学的发展趋势. 动物学杂志 , 1989, 24 (4) : 43–45. |
| [2] | 尚玉昌. 行为生态学. 北京: 北京大学出版社, 1998: 20-29, 141-149, 240-270. |
| [3] | 尚玉昌. 动物行为研究的新进展 (六): 性选择和配偶选择. 自然杂志 , 2013, 35 (3) : 207–210. |
| [4] | Frame A M, Mills A F. Condition-dependent mate choice: A stochastic dynamic programming approach. Theoretical Population Biology , 2014, 96 : 1–7. DOI:10.1016/j.tpb.2014.06.001 |
| [5] | Atwell A, Wagner W E Jr. Female mate choice plasticity is affected by the interaction between male density and female age in a field cricket. Animal Behaviour , 2014, 98 : 177–183. DOI:10.1016/j.anbehav.2014.10.007 |
| [6] | 张建军, 张知彬. 动物的性选择. 生态学杂志 , 2003, 22 (4) : 60–64. |
| [7] | Kirkpatrick M. Sexual selection by female choice in polygynous animals. Annual Review of Ecology and Systematics , 1987, 18 : 43–70. DOI:10.1146/annurev.es.18.110187.000355 |
| [8] | Andersson M. Sexual Selection. Princeton: Princeton University Press, 1994. |
| [9] | Dunning J L, Pant S, Bass A, Coburn Z, Prather J F. Mate choice in adult female bengalese finches: females express consistent preferences for individual males and prefer female-directed song performances. PLoS One , 2014, 9 (2) : e89438. DOI:10.1371/journal.pone.0089438 |
| [10] | Catchpole C K, Slater P J B. Bird Song: Biological Themes and Variations. Cambridge: Cambridge University Press, 1995. |
| [11] | Catchpole C K, Dittami J, Leisler B. Differential responses to male song repertoires in female songbirds implanted with oestradiol. Nature , 1984, 312 (5994) : 563–564. DOI:10.1038/312563a0 |
| [12] | Searcy W A, Marler P. A test for responsiveness to song structure and programming in female sparrows. Science , 1981, 213 (4510) : 926–928. DOI:10.1126/science.213.4510.926 |
| [13] | Eens M, Pinxten R, Verheyen R F. Male song as a cue for mate choice in the European starling. Behaviour , 1991, 116 (3) : 210–238. DOI:10.1163/156853991X00049 |
| [14] | Wasserman F E, Cigliano J A. Song output and stimulation of the female in white-throated sparrows. Behavioral Ecology and Sociobiology , 1991, 29 (1) : 55–59. DOI:10.1007/BF00164295 |
| [15] | Hernandez A M, Phillmore L S, MacDougall-Shackleton S A. Effects of learning on song preferences and Zenk expression in female songbirds. Behavioural processes , 2008, 77 (2) : 278–284. DOI:10.1016/j.beproc.2007.11.001 |
| [16] | Marler P. Bird songs and mate selection//Lanyon W E, Tavolga W, eds. Animal Sounds and Communication. Washington: American Institute of Behavioral Science, 1960: 348-367. |
| [17] | Baker M C. Vocal dialect recognition and population genetic consequences. American Zoologist , 1982, 22 (3) : 561–569. DOI:10.1093/icb/22.3.561 |
| [18] | Lambrechts M M, Dhondt A A. Individual voice discrimination in birds. Current Ornithology , 1995, 12 : 115–139. |
| [19] | 王爱真, 雷富民, 贾志云. 雌性选择与雄鸟鸣唱的进化. 动物学研究 , 2003, 24 (4) : 305–310. |
| [20] | Ptacek M B. The role of mating preference in shaping interspecific divergence in mating signal s in vertebrates. Behavioural Processes , 2000, 51 (1/3) : 111–134. |
| [21] | 雷富民. 鸟类鸣声结构地理变异及其分类学意义. 动物分类学报 , 1999, 24 (2) : 232–240. |
| [22] | 雷富民, 王钢. 鸟类鸣声行为对其物种分化和新种形成影响. 动物分类学报 , 2002, 27 (3) : 641–648. |
| [23] | Vallet E, Beme I, Kreutzer M. Two-note syllables in canary songs elicit high levels of sexual display. Animal Behaviour , 1998, 55 (2) : 291–297. DOI:10.1006/anbe.1997.0631 |
| [24] | Zahavi A. Mate selection-a selection for a handicap. Journal of Theoretical Biology , 1975, 53 (1) : 205–214. DOI:10.1016/0022-5193(75)90111-3 |
| [25] | Kroodsma D E, Miller E H. Ecology and evolution of acoustic communication in birds. Comstock: Cornell University Press , 1996 : 409–425. |
| [26] | Mcgregor P K. Animal communication networks. Cambridge: Cambridge University Press, 2005. |
| [27] | Naguib M, Todt D. Effects of dyadic vocal interactions on other conspecific receivers in nightingales. Animal Behaviour , 1997, 54 (6) : 1535–1543. DOI:10.1006/anbe.1997.9997 |
| [28] | Naguib M, Fichtel C, Todt D. Nightingales respond more strongly to vocal leaders of simulated dyadic interactions. Proceedings of the Royal Society of London.Series B: Biological Sciences , 1999, 266 (1419) : 537–542. DOI:10.1098/rspb.1999.0669 |
| [29] | Oliveira R F, McGregor P K, Latruffe C. Know thine enemy: fighting fish gather information from observing conspecific interactions.. Proceedings of the Royal Society of London.Series B: Biological Sciences , 1998, 265 (1401) : 1045–1049. DOI:10.1098/rspb.1998.0397 |
| [30] | Peake T M, Terry A M R, McGregor P K, Dabelsteen T. Male great tits eavesdrop on simulated male-to-male vocal interactions. Proceedings of the Royal Society of London. Series B: Biological Sciences , 2001, 268 (1472) : 1183–1187. DOI:10.1098/rspb.2001.1648 |
| [31] | Amy M, Leboucher G. Male canaries can visually eavesdrop on conspecific food interactions. Animal Behaviour , 2007, 74 (1) : 57–62. DOI:10.1016/j.anbehav.2006.07.019 |
| [32] | Amy M, Leboucher G. Effects of eavesdropping on subsequent signalling behaviours in male canaries. Ethology , 2009, 115 (3) : 239–246. DOI:10.1111/eth.2009.115.issue-3 |
| [33] | Doutrelant C, McGregor P K. Eavesdropping and mate choice in female fighting fish. Behaviour , 2000, 137 (12) : 1655–1668. DOI:10.1163/156853900502763 |
| [34] | Ophir A G, Galef Jr B G. Female Japanese quail that ‘eavesdrop’ on fighting males prefer losers to winners. Animal Behaviour , 2003, 66 (2) : 399–407. DOI:10.1006/anbe.2003.2230 |
| [35] | Ophir A G, Persaud K N, Galef Jr B G. Avoidance of relatively aggressive male Japanese quail (Coturnix japonica) by sexually experienced conspecific females. Journal of Comparative Psychology , 2005, 119 (1) : 3–7. DOI:10.1037/0735-7036.119.1.3 |
| [36] | Leboucher G, Pallot K. Is he all he says he is? Intersexual eavesdropping in the domestic canary, Serinus canaria. Animal Behaviour , 2004, 68 (4) : 957–963. DOI:10.1016/j.anbehav.2003.12.011 |
| [37] | Amy M, Monbureau M, Durand C, Gomez D, Thery M, Leboucher G. Female canary mate preferences: differential use of information from two types of male-male interaction. Animal Behaviour , 2008, 76 (3) : 971–982. DOI:10.1016/j.anbehav.2008.03.023 |
| [38] | Otter K, McGregor P K, Terry A M R, Burford F R L, Peake T M, Dabelsteen T. Do female great tits (Parus major) assess males by eavesdropping? A field study using interactive song playback. Proceedings of the Royal Society of London. Series B: Biological Sciences , 1999, 266 (1426) : 1305–1309. DOI:10.1098/rspb.1999.0779 |
| [39] | Mennill D J, Ratcliffe L M, Boag P T. Female eavesdropping on male song contests in songbirds. Science , 2002, 296 (5569) : 873–873. DOI:10.1126/science.296.5569.873 |
| [40] | Doutrelant C, McGregor P K, Oliveira R F. The effect of an audience on intrasexual communication in male Siamese fighting fish, Betta splendens. Behavioral Ecology , 2001, 12 (3) : 283–286. DOI:10.1093/beheco/12.3.283 |
| [41] | Matos R J, Schlupp I. Performing in front of an audience: signallers and the social environment//McGregor P K, ed. Animal Communication Networks. Cambridge: Cambridge University Press, 2005: 63-83. |
| [42] | McGregor P K, Peake T M. Communication networks: social environments for receiving and signalling behaviour. Acta Ethologica , 2000, 2 (2) : 71–81. DOI:10.1007/s102110000015 |
| [43] | Bitetti M S D. Food-associated calls and audience effects in tufted capuchin monkeys, Cebus apella nigritus. Animal Behaviour , 2005, 69 (4) : 911–919. DOI:10.1016/j.anbehav.2004.05.021 |
| [44] | Plath M, Blum D, Schlupp I, Tiedemann R. Audience effect alters mating preferences in a livebearing fish, the Atlantic molly, Poecilia mexicana. Animal Behaviour , 2008, 75 (1) : 21–29. DOI:10.1016/j.anbehav.2007.05.013 |
| [45] | Ung D, Amy M, Leboucher G. Heaven it's my wife! Male canaries conceal extra-pair courtships but increase aggressions when their mate watches. PLoS One , 2011, 6 (8) : e22686. DOI:10.1371/journal.pone.0022686 |
| [46] | 陈云霜. 灰文鸟的饲养. 园林 , 1999, 6 : 41–41. |
| [47] | 李顺才, 张坤. 灰文鸟的人工养殖技术. 中国家禽 , 2004, 26 (10) : 26–27. |
| [48] | 徐磊. 灰文鸟的饲养与繁殖. 特种经济动植物 , 2004 (4) : 13–13. |
| [49] | Follett B K, Hinde R A, Steel E, Nicholls T J. The influence of photoperiod on nest building, ovarian development and luteinizing hormone secretion in canaries (Serinus canarius). Journal of Endocrinology , 1973, 59 (1) : 151–162. DOI:10.1677/joe.0.0590151 |
| [50] | Beguin N, Leboucher G, Bruckert L, Kreutzer M. Mate preferences in female canaries (Serinus canaria) within a breeding season. Acta Ethologica , 2006, 9 (2) : 65–70. DOI:10.1007/s10211-006-0017-3 |
| [51] | Collins S A, Hubbard C, Houtman A M. Female mate choice in the zebra finch-the effect of male beak colour and male song. Behavioral Ecology and Sociobiology , 1994, 35 (1) : 21–25. DOI:10.1007/BF00167055 |
| [52] | Rutstein, A N, Brazill-Boast J, Griffith S C. Evaluating mate choice in the zebra finch. Animal Behavior , 2007, 74 (5) : 1277–1284. DOI:10.1016/j.anbehav.2007.02.022 |
| [53] | Weary D M. Categorical perception of bird song: How do great tits (Parus major) perceive temporal variation in their song?. Journal of Comparative Psychology , 1989, 103 (4) : 320–325. DOI:10.1037/0735-7036.103.4.320 |
| [54] | Okanoya K, Dooling R J. Perception of distance calls by budgerigars (Melopsittacus undulatus) and zebra finches (Poephila guttata): Assessing species-specific advantages. Journal of Comparative Psychology , 1991, 105 (1) : 60–72. DOI:10.1037/0735-7036.105.1.60 |
| [55] | Cynx J. Conspecific song perception in zebra finches (Taeniopygia guttata). Journal of Comparative Psychology , 1993, 107 (4) : 395–402. DOI:10.1037/0735-7036.107.4.395 |
| [56] | Weeden J S, Falls J B. Differential responses of male ovenbirds to recorded songs of neighboring and more distant individuals. The Auk , 1959, 76 (3) : 343–351. DOI:10.2307/4081812 |
| [57] | Forstmeier W, Kempenaers B, Meyer A, Leisler B. A novel song parameter correlates with extra-pair paternity and reflects male longevity. Proceedings of the Royal Society of London. Series B: Biological Sciences , 2002, 269 (1499) : 1479–1485. DOI:10.1098/rspb.2002.2039 |
| [58] | Cotton S, Small J, Pomiankowski A. Sexual selection and condition-dependent mate preferences. Current Biology , 2006, 16 (17) : R755–R765. DOI:10.1016/j.cub.2006.08.022 |
| [59] | 邵明勤, 阿布力米提·阿布都卡迪尔, 高行宜, 孙铭娟. 鸟类行为研究进展. 干旱区研究 , 2002, 19 (2) : 75–80. |
| [60] | Price A C, Rodd F H. The effect of social environment on male-male competition in guppies (Poecilia reticulata). Ethology , 2006, 112 (1) : 22–32. DOI:10.1111/eth.2006.112.issue-1 |
| [61] | Tachon G, Murray A M, Gray D A, Cade W H. Agonistic displays and the benefits of fighting in the field cricket, Gryllus bimaculatus. Journal of Insect Behavior , 1999, 12 (4) : 533–543. DOI:10.1023/A:1020970908541 |
| [62] | Baltz A P, Clark A B. Extra-pair courtship behaviour of male budgerigars and the effect of an audience. Animal Behaviour , 1997, 53 (5) : 1017–1024. DOI:10.1006/anbe.1996.0354 |
| [63] | Clutton-Brock T. Sexual selection in females. Animal Behaviour , 2009, 77 (1) : 3–11. DOI:10.1016/j.anbehav.2008.08.026 |
| [64] | Oring L W, Colwell M A, Reed J M. Lifetime reproductive success in the spotted sandpiper (Actitis macularia): sex differences and variance components. Behavioral Ecology and Sociobiology , 1991, 28 (6) : 425–432. |
| [65] | Oring L W, Reed J M, Colwell M A, Lank D B, Maxson S J. Factors regulating annual mating success and reproductive success in spotted sandpipers (Actitis macularia). Behavioral Ecology and Sociobiology , 1991, 28 (6) : 433–442. |
| [66] | Kvarnemo C, Moore G I, Jones A G. Sexually selected females in the monogamous Western Australian seahorse. Proceedings of the Royal Society of London, Series B: Biological Sciences , 2007, 274 (1609) : 521–525. DOI:10.1098/rspb.2006.3753 |
| [67] | Tregenza T, Wedell N. Benets of multiple mates in the cricket Gryllus bimaculatus. Evolution , 1998, 52 (6) : 1726–1730. DOI:10.2307/2411345 |
| [68] | Fisher D O, Double M C, Blomberg S P, Jennions M D, Cockburn A. Post-mating sexual selection increases lifetime fitness of polyandrous females in the wild. Nature , 2006, 444 (7115) : 89–91. DOI:10.1038/nature05206 |