 2016, Vol. 36
2016, Vol. 36文章信息
- 郑云普, 党承华, 郝立华, 程东娟, 徐明
- ZHENG Yunpu, DANG Chenghua, HAO Lihua, CHENG Dongjuan, XU Ming.
- 华北平原玉米叶片光合及呼吸过程对实验增温的适应性
- Photosynthetic and respiratory acclimation of maize leaves to experimental warming in the North China Plain
- 生态学报[J]. 2016, 36(16): 5236-5246
- Acta Ecologica Sinica[J]. 2016, 36(16): 5236-5246
- http://dx.doi.org/10.5846/stxb201501190154
-
文章历史
- 收稿日期: 2015-01-19
- 网络出版日期: 2015-11-30

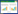




2. 中国科学院地理科学与资源研究所 生态系统网络观测与模拟重点实验室, 北京 100101
2. Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographical Sciences and Natural Resources, Chinese Academy of Sciences, Beijing 100101, China
自从工业革命以来,大气温室气体浓度的增加已经导致全球地面平均温度上升约0.74 ℃,到21世纪末全球地表温度还将会上升1.1—6.4 ℃[1]。气候变暖通过多种方式来间接或直接地影响植物的生理生化过程[2-11],并进一步导致生物量积累及其分配格局的变化[12-14]。相关研究结果表明,全球变暖不仅能够通过增加土壤氮的矿化速率及其有效性[15-19]、降低土壤湿度[20-21]以及延长生长季[21-22]对植物产生间接影响,还可以通过改变植物的光合与呼吸过程直接改变植物的生长和发育进程[10, 12, 23-27]。
近年来,相关的研究结果显示,植物的光合特性能够对其生长环境的温度产生一定的适应性,即光合反应过程的最适温度会随着植物生存环境温度的改变而发生移动[28-30]。然而,目前关于引起植物光合作用最适温度变化的机理还并不是十分的清楚。一些研究结果表明,光系统Ⅱ的电子传递决定着光合作用最适温度的适应性[31-32];例如,近年来的研究利用叶绿素荧光技术发现,光系统Ⅱ的热稳定性在不同的生长温度下发生了变化[33]。另有研究的结果还表明,植物光合作用最大反应速率最适温度的移动还同1,5-二磷酸核酮糖(Ribulose-1,5-Bisphosphate,RuBP)的羧化、再生和活性有着非常密切的关系[34-38]。同时,在不同的生长温度下光合作用的最大电子传递速率(The Maximum Capacity of Electron Transport Mediated RuBP Regeneration,Jmax)的最适温度也会发生移动[34, 39-40]。因此,这种Jmax对生长温度的响应也可能是引起RuBP再生限制净光合的温度曲线移动的原因之一。另外,以往的研究还发现,RuBP再生过程中所需酶的耐热性在不同的生长温度下发生了变化。Badger等[31]发现夹竹桃叶片卡尔文循环中不同酶的热稳定性在不同的生长温度下发生了变化。此外,RuBP羧化和再生之间的平衡也可能会影响植物光合的温度敏感性[37, 41]。由于RuBP羧化限制的A和RuBP再生限制的A的温度敏感性不同[34, 42],则不同的生长温度下受RuBP羧化限制的A和受RuBP再生限制的A的温度敏感性也会发生相应的变化。当植物从较低的生长温度转移到较高的生长温度下时,RuBP羧化限制的A和RuBP再生限制的A会同时增加,但是两者的比率会降低,此时A就变为受RuBP羧化和再生的双重限制。例如,Wullschleger[43]对109个物种的最大羧化反应速率( Vcmax)和Jmax进行整合分析的结果显示,尽管生长环境和物种间存在差异,但是Jmax和Vcmax两者的比率为定值。然而,Hikosaka等[34]的研究却发现不同生长温度下栓皮栎(Quercus myrsinaefolia)的Jmax/Vcmax发生了改变。植物叶片呼吸过程对温度的响应可以表现在短期和长期两个不同的时间尺度上[44]。以往的研究普遍认为呼吸速率与温度之间是指数函数的关系,并且一般认为呼吸的温度敏感性为一个常数Q10= 2.0[44-45]。然而,植物呼吸对温度的响应非常敏感,其温度敏感性Q10并不是一个常数并且会随着温度的改变而发生变化[46-48]。从另一方面来讲,长期的增温也会导致植物的呼吸过程对温度产生一定的适应性[47],则Q10也不是一个固定不变的值。尽管如此,目前许多关于碳平衡的模型将Q10 作为一个定值来描述呼吸对温度的短期响应,并且忽视其对温度的适应性。从机理上来说,通常影响植物呼吸温度敏感性Q10最主要的两个因素是温度和底物供应[44],其中温度主要是通过影响呼吸相关的酶,从而对呼吸的温度敏感性产生影响[45]。然而,植物的呼吸过程在比较合适的温度下几乎不会受到酶活性的限制,这主要是由于此时无论是可溶性酶还是与膜结合的酶都具有较高的Vmax[46];这种情况下限制呼吸过程的因素就成为底物的供应状况[45-47]。底物的供应能力也会随着温度的改变而变化,从而影响到呼吸的温度敏感性[48-50]。
华北平原是我国最重要的粮食产区之一,该区域主要以玉米和小麦轮作系统为主,每年全国大约50%的小麦和30%的玉米均产自于该粮食产区[51]。然而,值得关注的是,许多气候模型已经证实我国的气候正在逐渐呈现变暖的趋势,特别是自从20世纪70年代以后我国北部地区 (如华北平原) 的气候呈现出年平均气温升高的特征[51-54]。这种区域甚至全球范围内的气候变暖势必改变农作物的蒸散和水分利用效率[55],并进一步影响其生长速率和粮食产量[56]。此外,近年来的许多研究结果还表明,全球变暖也会通过改变农作物的生长季而对其物候期产生深远的影响。例如,Tao等[57]的研究表明,全球气候变暖导致作物产量形成的重要生长阶段(从开花到成熟)呈现延长的趋势。Hou等[58]在华北平原的增温实验研究发现,气候变暖将会缩短小麦的生长季,但并不改变其生育期,最终导致小麦产量减少。同样,以往的研究结果已经发现气候变暖导致全球许多地区玉米产量的减少[59]。由此看来,气候变暖对农作物产量的影响将会直接关系到未来区域/全球粮食供应的安全问题。然而,目前全球气候变暖对作物产量产生影响的机理还不清楚。此外,值得注意的是叶片光合和呼吸等关键生理过程在一定程度上决定着粮食的产量;同时,农作物也可能会对光合和呼吸两大生理过程的最适温度做出调整,来应对气候变暖给其带来的不利影响,即在一定程度上适应气候变暖。因此,深入探讨作物叶片的光合和呼吸过程对未来气候变暖的适应能力将有助于准确估算未来气候变暖对华北平原粮食产量的影响以及全面理解全球粮食供应面临的风险。本研究利用野外原位的红外线增温实验,探讨华北平原典型农田生态系统玉米叶片的光合和呼吸过程对实验增温的适应性机理,为预测未来气候变暖对该区域的粮食产量产生的影响和全球粮食安全供应问题提供数据支撑和理论依据。
1 材料与方法 1.1 研究区概况本研究在中国科学院禹城综合试验站((36°57′N,116°38′E,海拔 23.4 m)完成。该试验站位于山东省禹城市,属大陆性季风气候暖温带地区。年平均气温 13.1 ℃,1月份平均温度-3 ℃,7月份平均温度26.9 ℃;年平均降水量610 mm,降水季节分配不均匀,3—5月平均降水量75.7 mm,占年平均降水量的12.4%;6—8月降水量平均为419.7 mm,占年平均降水量的68.8%。实验站土壤质地以粉砂和轻壤为主;pH值较高,为7.9—8.0;土壤有机质含量较低,为0.6%—1.0%;全N含量为0.05%—0.065%。从土壤自然条件看,该站在华北平原具有典型代表性。当地种植制度以冬小麦-夏玉米轮作为主。
1.2 野外增温实验设计在2009年9月份,在试验区随机选取6个2 m × 3 m 的样方用以布置增温实验。随机选取其中的3个样方作为增温组,另3个为对照组。3个增温样方从2009年11月18日开始利用悬挂于样方正上方距离地表 2.25 m高度的MSR-2420型红外发射器(165×1.5 cm,Kalglo Electronics Inc.,Bethlehem,PA) 进行连续的增温处理。每个对照样方内同样在距离地面2.25 m高度处悬挂一个相同大小和形状的“伪”红外增温装置模拟红外发射器对下方土壤产生阴影的效果。增温样方和对照样方相距约5 m,避免红外增温装置对对照样方产生影响。利用PT100热电偶系统(Unism Technologies Incorporated,Beijing,China)自动记录每小时2.25 m高度处的空气温度和5 cm 深处的土壤温度。另外,在本研究中还利用红外温度计 (FLUKE 574,Fluke Inc.,USA) 测量玉米冠层叶片表面的温度。在整个玉米生长季从2011年6月24日到2011年10月7日,相对于对照样地,增温样地平均的空气温度、土壤温度、冠层温度分别增加了(1.42±0.18)/(1.77±0.24) ℃(白天/夜晚)、(1.68±0.9)/(2.04±0.16) ℃(白天/夜晚)、(2.08±0.72) ℃(白天)。同时,本实验还利用FDS100土壤湿度传感器(Unism Technologies Incorporated,Beijing,China)监测了样方内0—10 cm深度土壤湿度的变化状况。在整个监测时期,对照样地10 cm 深处土壤湿度为26%,而增温样地10 cm 深处土壤湿度为25%[58]。
1.3 实验材料优良玉米品种 (Zea may L. cv. Zhengdan 958) 的种子播种之前先在黑暗、干燥的环境下4 ℃冷藏2 d。随后,2011年6月24日将玉米种子分别播种到增温和对照样地,增温和对照样地的玉米苗均于7月1日出苗。为了避免干旱胁迫,利用地下水在整个生长季从2011年6月24日到2011年10月7日对各个样地的玉米苗灌溉。随机从增温和对照样地中选取1个样地作为取样的样地。由于玉米穗位叶是决定玉米产量最重要的叶片,因此在玉米播种60 d后(即2011年8月24日)在所选的采样样地各随机采集5个完全伸展的穗位叶进行各个指标的测量和观察。
1.4 实验方法 1.4.1 叶片An/Ci曲线的测定本研究利用Li-6400便携式光合测定系统 (LI-COR Inc. Lincoln,Nebraska,USA)来测定玉米叶片的An/Ci曲线。同Li-6400便携式光合测定系统配套使用的2 cm × 3 cm 标准气室能独立控制光合光量子通量密度 (PPFD)、CO2浓度、叶片温度及气室湿度。由于Li-6400便携式光合测定系统自身对叶片温度的调整范围非常有限,通过增加一些具有内部水槽的金属块来加热或冷却叶室。这些水槽通过塑料管与2个既能加热又能冷却的容器相连接。该容器加热或冷却的功能通过添加热水或冰水来实现,最终达到改变叶室温度的目的。在华北平原夏季玉米的生长季,经过改造后的Li-6400便携式光合测定系统能够使叶片温度的调控范围扩大到10— 40 ℃。
分别从增温和对照样地随机的选择5个完全伸展开的穗位叶进行不同叶片温度下典型An/Ci曲线的测定。分别测定以5 ℃为间隔从10 ℃升高到40 ℃叶片温度下的An/Ci曲线。为了利用自然环境的温度变化,从7:00 am开始测定低温下的An/Ci曲线(叶片温度10 ℃),然后随着环境温度的逐渐升高,再测定稍高温度下的An/Ci曲线(叶片温度15 ℃、20 ℃和25 ℃);当下午空气温度最高的时候测定高温下的An/Ci曲线(叶片温度30 ℃、35 ℃和40 ℃)。每次在测定An/Ci曲线之前先使叶片处于饱和的光合光量子通量密度(1500 μmol photons m-2 s-1)和大气CO2浓度(380 μL/L)下至少10min,以保证气室内叶片气体交换过程达到平衡。为了避免测定温度的大幅改变导致Li-6400便携式光合测定系统气室内水汽凝结,从而影响测定数据和结果的准确性,在整个An/Ci曲线测定过程中始终控制气内的相对湿度低于70%。待An/Ci曲线的测定开始后,Li-6400便携式光合测定系统气室内的CO2浓度首先逐渐从380 μL/L降低为50 μL/L,然后再由380 μL/L增加到1200 μL/L,每条An/Ci曲线均由9个数据点组成,气室内具体的的CO2浓度变化为5、100、200、300、380、600、800、1000、1200 μL/L。
1.4.2 叶片暗呼吸速率的测定当An/Ci曲线测定结束后关闭叶室内的LED光源至少5 min,然后进行叶片暗呼吸的测定。叶片暗呼吸测定的过程中保持LED光源处于关闭状态,其它所有的测定条件与An/Ci曲线测定时一致,整个测定过程中保证气室内的CO2浓度固定为380 μL/L不变。叶片暗呼吸的测定过程中以30 s间隔采集5个数据值。将这5个数值的平均值作为该叶片在特定温度下的呼吸值。
1.4.3 参数拟合和计算根据Farquhar光合作用模型(FvCB),光合作用受到Rubisco羧化过程(Ac)影响时的计算方程为
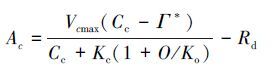 (1)
(1)
式中,Vcmax为最大羧化反应速率,Cc是Rubisco羧化点的CO2分压,Г*代表了CO2的补偿点,Kc和Ko分别是Rubisco酶对于CO2和O2的Michaelis-Menten常数,O为叶绿体处的氧气浓度,Rd指白天的呼吸速率[41]。 当光合作用受到RuBP再生限制时,光合作用的方程为:
 (2)
(2)
式中,Jmax表示最大电子传递速率。净光合速率的值由Ac和Aj中较小的值决定。
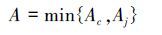 (3)
(3)
本研究中利用Sharkey等[60]的Microsoft Excel软件对测定的所有An/ Ci曲线拟合出参数Vcmax和Jmax。另外,叶片暗呼吸的Q10用方程Q10 = e10b来计算[48]。
1.4.4 叶片组织碳水化合物及碳氮元素测定将用于气体交换参数测定的植物叶片样品在75 ℃下烘干至恒重,然后利用球形研磨仪研磨成粉末。根据Wong[61]和Hoch等[62]的方法提取和确定葡萄糖、蔗糖、果糖及淀粉的含量。总非结构性碳含量(Non-structural Carbohydrates,NSC)为可溶性糖(葡萄糖、蔗糖及果糖)和淀粉的总和。另外,利用元素分析仪(Vario Max CN; Elemnetar Corp.,Germany) 测定叶片内总碳、总氮的含量,并进一步计算组织中的碳氮比率。所有化学成分含量的表达均基于样品干重,每个指标的测量均有5个重复。
1.5 数据统计分析本研究中关于增温对玉米叶片生理参数影响的数据利用多因素方差分析的方法(ANOVA)比较处理间的显著性差异。本研究中所有的统计分析均利用SPSS 13.0 (Chicago,IL) 统计软件完成,所有作图利用Sigmaplot来实现。
2 结果与分析 2.1 增温对光合及呼吸作用的影响无论是对照还是增温处理的玉米叶片净光合-叶温曲线(An-Tleaf Curve)均呈现出典型的“钟形”分布 (图 1)。研究结果表明,增温导致玉米叶片的净光合速率显著升高 (P<0.001)。增温还导致玉米叶片An-Tleaf曲线向高温端发生了移动。增温使基于叶面积净光合速率的最适温度从28.97 ℃增加到30.53 ℃ (图 1)。另外,增温显著降低了玉米叶片的暗呼吸速率Rd(P<0.001,图 1),同时降低了Rd 的温度敏感性Q10,增温使玉米基于叶面积暗呼吸的Q10由1.53降低为1.44 (P<0.05,图 1)。

|
| 图 1 玉米叶片的净光合速率(An)及暗呼吸速率(Rd)对增温的响应 Fig. 1 Responses of net photosynthesis rates (An) and dark respiration rates (Rd) to experimental warming in maize leaves |
同净光合速率对温度的响应相似,对照和增温叶片Jmax对温度的响应曲线也均呈现出典型的“钟形” (图 2)。然而,增温叶片的Jmax显著高于对照(P<0.001),尤其是在较高的叶片温度下(图 2)。本研究结果还表明,增温不仅使Jmax-Tleaf曲线在纵向上发生移动,而且还导致Jmax- Tleaf曲线在水平方向上产生移动。我们的研究发现增温使Jmax的最适温度由31.84 ℃升高为33.29 ℃(P<0.05,图 2)。另外,本研究的结果还显示,对照和增温处理叶片的Vcmax值非常相似,表明增温对低温下的Vcmax几乎没有影响。统计的结果表明,增温确实没有对Vcmax产生影响(P=0.259)。另外,增温也没有对Vcmax的温度敏感性产生显著的影响 (P>0.05,图 2)。

|
| 图 2 最大羧化反应速率(Vcmax)和最大电子传递速率(Jmax) 对增温响应 Fig. 2 Responses of Vcmax and Jmax to experimental warming in maize |
由于净光合作用速率为总光合速率与呼吸速率的差值,因此植物呼吸过程和光合过程之间的平衡关系是影响净光合作用温度响应及其最适温度的重要特征。本研究的结果发现,对照和增温处理的玉米叶片暗呼吸速率与总光合速率的比值(Rd/Ag) 随着温度的上升均呈现出增加的趋势,但是增温和对照组的差异并不显著(P=0.181,图 3)。另外,本研究的结果还显示Jmax/ Vcmax比率同测量温度之间呈现出线性相关关系(对照r2 =0.89;增温r2 =0.81)(图 3)。同时,统计结果表明实验增温并没有显著影响Jmax/Vcmax的比率 (P=0.104,图 3)。

|
| 图 3 增温对玉米叶片Rd/Ag 和 Jmax/Vcmax比值的影响 Fig. 3 Warming effects on Rd/Ag ratio Jmax/Vcmax ratio of maize leaves |
本研究的结果显示,对照和增温叶片的气孔导度(Gs)均随着叶片温度的逐渐增加呈现出先增加后降低的趋势(图 4)。对照和增温叶片的Gs在叶片温度分别达到约20 ℃和25 ℃之前,Gs呈现增加的趋势;但当叶片温度超过20 ℃和25 ℃后Gs却逐渐降低。更有趣的是增温降低了叶片在低温时(10—20 ℃)的Gs,但却增加了叶片在高温时(25—40 ℃)的Gs。然而,统计结果却表明,增温并没有显著增加玉米叶片的Gs(P>0.05)。另外,与Gs不同的是对照和增温处理下玉米叶片的蒸腾速率(Tr)均随着叶片温度的升高而逐渐增加 (图 4)。在叶片温度低于大约25 ℃时对照和增温叶片的Tr差别不大;但当叶片温度高于约25 ℃时,增温处理叶片的Tr明显高于对照叶片。同时。本研究的统计结果表明增温显著增加了玉米叶片的Tr (P<0.01)。另外,对照和增温叶片的水分利用效率(WUE)对温度的响应均呈现出先增加再降低的趋势 (图 4)。当叶片温度升高到约30 ℃时,对照和增温叶片的WUE均达到最大值。增温使叶片最大的WUE增加约30%。此外,本研究结果还显示当叶片温度低于约25 ℃时增温增加叶片的WUE,但当叶片温度高于约25 ℃时增温却降低叶片的WUE (图 4)。然而,统计结果表明增温并没有显著增加叶片的WUE (P>0.05)。

|
| 图 4 玉米叶片气孔导度(Gs)、蒸腾速率(Tr)和水分利用效率(WUE)对实验增温的响应 Fig. 4 Responses of stomatal conductance (Gs),transpiration rates (Tr),and water use efficiency (WUE) to experimental warming in maize leaves |
本研究的结果表明,尽管增温并没有显著改变玉米叶片中淀粉的含量,但是却增加可溶性糖(主要为葡萄糖、蔗糖及果糖)的含量,从而显著提高叶片总非结构性碳的含量(NSC)。相对于对照温度,增温分别提高玉米叶片中葡萄糖、果糖、蔗糖的含量25%、53% 和83%,从而使NSC含量增加约44%(P<0.05,表 1)。然而,尽管增温没有对叶片中淀粉的浓度产生影响,但由于叶片可溶性糖含量的显著增加,导致叶片NSC含量增加约30%(表 1)。同时,实验增温显著提高叶片的C/N比 (P<0.05),但并没有明显影响到基于干重的叶片C、N元素的含量。具体来讲,对照环境下玉米叶片中的碳氮元素含量低于增温环境下的叶片,但增温并没有显著增加单位干重叶片C、N元素的含量(表 1)。更为有趣的是尽管增温没有显著改变叶片中C、N的浓度,但却明显增加了叶片C/N比约10%(表 1)。
| 对照 Control | 增温 Warming | 增量 Increasing/% | P | |
| 碳水化合物含量 Non-structural carbohydrates/(mg/g干重) | ||||
| 葡萄糖 Glucose | 16.2±2.7 | 20.2±1.8 | 24.9 | 0.048 |
| 果糖 Fructose | 11.4±1.8 | 17.4±0.9 | 52.8 | 0.003 |
| 蔗糖 Sucrose | 5.1±1.1 | 9.4±1.9 | 83.0 | 0.009 |
| 可溶性糖 Soluble sugars | 32.7±4.6 | 47.0±1.4 | 43.7 | 0.001 |
| 淀粉 Starch | 37.5±4.3 | 45.0±3. 6 | 20.0 | 0.080 |
| 非结构性碳 NSC | 69.9±5.2 | 91.3±4.5 | 30.8 | 0.006 |
| 组织碳/氮比率 Tissue C/N ratio (mg/g湿重) | ||||
| 总碳 Total carbon | 436.0±3 | 439.0±3 | 0.8 | 0.179 |
| 总氮 Total nitrogen | 28.5±1.6 | 26.4±0.7 | -7.4 | 0.058 |
| 碳氮比 C/N ratio | 15.4±0.7 | 16.6±0.3 | 7.8 | 0.028 |
许多基于实验室增温或野外增温的实验研究探讨了不同物种光合作用对温度的响应及适应性机理[25, 38, 40]。本研究的结果显示,野外实验增温使华北平原玉米光合作用的最适温度从27.50 ℃到29.06 ℃增加了1.56 ℃(图 1)。然而,值得注意的是不同物种的光合作用对于增温的响应存在着很大的差异,尤其是C3和C4利用不同光合途径的物种之间的差异会更大。玉米是一种典型的C4植物,而C4植物可能对增温比其他的C3植物更加敏感,这可能主要是由于不同物种对温度响应的差异造成的。本研究中玉米叶片的净光合速率的最适温度增加了1.56 ℃,表明叶片在增温条件下对高温环境产生了适应(图 1)。根据解释光合作用适应性机制提出的5个假说,通过对光合作用二氧化碳羧化系统、电子传递系统、呼吸作用与光合作用平衡关系以及二氧化碳扩散过程等几个方面分析光合作用产生适应性的机制。在本研究中,增温没有对Vcmax的温度敏感性产生显著影响(图 2),表明 Rubisco酶羧化过程温度响应曲线的改变不是导致光合作用温度适应性的原因。然而,增温条件下Jmax的最适温度却向高温一端发生了移动(图 2),表明电子传递系统对高温的适应性可能是导致光合作用最适温度改变的原因之一。
以往的研究认为,Rubisco羧化和RuBP再生两个过程之间的平衡关系也可能会影响到光合作用的最适温度[37, 41, 43]。这种推断主要是基于光合系统中氮素利用效率最大化理论[38]。已有的研究已经发现,Rubisco羧化和RuBP再生两个光合系统的酶系统并不相同,并且这两个系统的温度敏感性也存在较大的差异[42]。通常来讲,氮素利用效率的最大化要求光合作用主要受到这两个过程的共同限制,假如只是受到其中一个系统的限制,那么就会造成N素在另一个系统之中的多余累积,就会导致氮素从这个系统转移到另一个系统之中[42]。当植物所处的生长温度发生改变时,由于这两个系统的温度敏感性不同,会有一个系统的速率增加的快,而另一个系统的速率增加的相对较慢,这就导致N素在两个系统中重新分配[42]。然而,近年来关于增温对Jmax/Vcmax比例影响的研究结果还尚无定论[37]。本研究的结果显示,增温对玉米叶片Jmax/Vcmax比值的影响不显著(图 3,P>0.05),表明增温并没有使玉米叶片Rubisco羧化和RuBP再生两个光合系统中N素的分配比例发生变化。因此,玉米叶片光合作用最适温度的移动并不是由于Jmax/Vcmax比值的改变而引起的。
从光合作用温度响应曲线来看,净光合速率在高温下的降低是由于呼吸作用在高温下非线性增加引起的[43],这也可能是决定光合作用最适温度的一个重要因素。总光合作用与呼吸作用之间的比例关系决定了净光合作用的最适温度[41-43]。呼吸作用的温度适应性常表现为低温下叶片暗呼吸速率的增长和在高温下暗呼吸速率的降低[44]。此外,关于光合和呼吸速率还存在一种平衡态观点,认为适应性的产生使得在一定的培养温度范围内,植物的光合速率和呼吸速率都维持在一个稳定的值[41]。FvCB光合作用模型理论认为,这种现象通常用呼吸速率与光合速率之间的比值(Rd/Ag)来表征[41]。本研究的结果显示,增温并没有显著影响玉米叶片呼吸速率与光合速率之间的比值Rd/Ag(图 3,P>0.05),表明玉米叶片光合作用最适温度的移动也不是由于Rd/Ag比值的改变而引起的。
通常,植物叶片的暗呼吸(Rd)与叶片温度之间呈现为典型的指数关系[45]。然而,植物生长温度的增加能够降低相同叶片温度下的呼吸速率,从而使呼吸的温度敏感性(Q10)降低[44]。以往的许多研究结果发现,植物叶片Rd的温度敏感性Q10随着植物生长温度的增加而降低[41-44]。本研究的结果也同样发现玉米生长温度的增加使叶片Rd的温度敏感性Q10显著降低(P<0.05),再次证实了叶片暗呼吸与植物生长温度之间的负相关关系。近年来的研究普遍认为,植物呼吸的温度敏感性主要由基质的有效性和最大的酶活性而决定[44]。然而,在本研究中增温增加了玉米叶片中非结构性碳水化合物(NSC)的含量 (尤其是可溶性糖的含量) (表 1)。因此,本研究中叶片Rd温度敏感性的降低可能是由于较高的生长温度使玉米叶片呼吸过程中关键酶活性的降低而引起的。另外,以往的大量研究结果表明,植物叶片含氮量同叶片呼吸存在正相关的关系[23]。我们的研究也发现增温使玉米叶片的组织含氮量降低约10% (表 1),由此分析认为,组织含氮量的降低也可能是导致玉米叶片Rd温度敏感性下降的原因之一。
4 结论本文利用华北平原典型农田生态系统的原位实验增温平台,研究了玉米叶片光合及呼吸过程对实验增温的适应能力,并深入探讨了其产生适应性的潜在原因和机理,得到如下4点结论:(1) 实验增温提高了玉米叶片的净光合反应速率(An)及其最适温度;(2) 实验增温导致最大电子传递速率(Jmax)及其最适温度的升高,表明电子传递系统对高温的适应性是导致光合作用最适温度改变的原因;(3) 实验增温降低了玉米叶片的暗呼吸速率(Rd)及其Q10值,但没有对Rd/ Ag和Jmax/Vcmax产生影响。(4) 玉米在叶片尺度上调控叶片光合及呼吸过程最适温度来适应增温环境的能力很有限,未来气候变暖仍可能会对华北平原玉米的生长发育过程以及粮食的品质和产量造成一定的影响。
| [1] | IPCC. Technical summary//Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt K B, Tignor M, Miller H L, eds. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge UK: Cambridge University Press, 2007. |
| [2] | Niu S L, Wan S Q. Warming changes plant competitive hierarchy in a temperate steppe in northern China. Journal of Plant Ecology , 2008, 1 (2) : 103–110. DOI:10.1093/jpe/rtn003 |
| [3] | Zhao C Z, Liu Q. Growth and physiological responses of Picea asperata seedlings to elevated temperature and to nitrogen fertilization. Acta Physiologiae Plantarum , 2009, 31 (1) : 163–173. DOI:10.1007/s11738-008-0217-8 |
| [4] | Klein J A, Harte J, Zhao X Q. Experimental warming causes large and rapid species loss, dampened by simulated grazing, on the Tibetan Plateau. Ecology Letters , 2004, 7 (12) : 1170–1179. DOI:10.1111/ele.2004.7.issue-12 |
| [5] | Walker M D, Wahren C H, Hollister R D, Henry G H R, Ahlquist L E, Alatalo J M, Bret-Harte M S, Calef M P, Callaghan T V, Carroll A B, Epstein H E, Jónsdóttir I S, Klein J A, Magnússon B, Molau U, Oberbauer S F, Rewa S P, Robinson C H, Shaver G R, Suding K N, Thompson C C, Tolvanen A, Totland Ø, Lee Turner P, Tweedie C E, Webber P J, Wookey P A. Plant community responses to experimental warming across the tundra biome. Proceedings of the National Academy of Sciences of the United States of America , 2006, 103 (5) : 1342–1346. DOI:10.1073/pnas.0503198103 |
| [6] | Franco A M A, Hill J K, Kitschke C, Collingham Y C, Roy D B, Fox R, Huntley B, Thomas C D. Impacts of climate warming and habitat loss on extinctions at species' low-latitude range boundaries. Global Change Biology , 2006, 12 (8) : 1545–1553. DOI:10.1111/gcb.2006.12.issue-8 |
| [7] | Biasi C, Meyer H, Rusalimova O, Hämmerle R, Kaiser C, Baranyi C, Daims H, Lashchinsky N, Barsukov P, Richter A. Initial effects of experimental warming on carbon exchange rates, plant growth and microbial dynamics of a lichen-rich dwarf shrub tundra in Siberia. Plant and Soil , 2008, 307 (1/2) : 191–205. |
| [8] | Jägerbrand A K, Alatalo J M, Chrimes D, Molau U. Plant community responses to 5 years of simulated climate change in meadow and heath ecosystems at a subarctic-alpine site. Oecologia , 2009, 161 (3) : 601–610. DOI:10.1007/s00442-009-1392-z |
| [9] | Kudo G, Suzuki S. Warming effects on growth, production, and vegetation structure of alpine shrubs: a five-year experiment in northern Japan. Oecologia , 2003, 135 (2) : 280–287. DOI:10.1007/s00442-003-1179-6 |
| [10] | Llorens L, Peñuelas J, Estiarte M, Bruna P. Contrasting growth changes in two dominant species of a Mediterranean shrubland submitted to experimental drought and warming. Annals of Botany , 2004, 94 (6) : 843–853. DOI:10.1093/aob/mch211 |
| [11] | Yin H J, Liu Q, Lai T. Warming effects on growth and physiology in the seedlings of the two conifers Picea asperata and Abies faxoniana under two contrasting light conditions. Ecological Research , 2008, 23 (2) : 459–469. DOI:10.1007/s11284-007-0404-x |
| [12] | Prieto P, Peñuelas J, Llusià J, Asensio D, Estiarte M. Effects of experimental warming and drought on biomass accumulation in a Mediterranean shrubland. Plant Ecology , 2009, 205 (2) : 179–191. DOI:10.1007/s11258-009-9608-1 |
| [13] | Lin D L, Xia J Y, Wan S Q. Climate warming and biomass accumulation of terrestrial plants: a meta-analysis. New Phytologist , 2010, 188 (1) : 187–198. DOI:10.1111/j.1469-8137.2010.03347.x |
| [14] | Wang J C, Duan B L, Zhang Y B. Effects of experimental warming on growth, biomass allocation, and needle chemistry of Abies faxoniana in even-aged monospecific stands. Plant Ecology , 2012, 213 (1) : 47–55. DOI:10.1007/s11258-011-0005-1 |
| [15] | Rustad L, Campbell J, Marion G, Norby R, Mitchell M, Hartley A, Cornelissen J, Gurevitch J. A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia , 2001, 126 (4) : 543–562. DOI:10.1007/s004420000544 |
| [16] | Peñuelas J, Gordon C, Llorens L, Nielsen T, Tietema A, Beier C, Bruna P, Emmett B, Estiarte M, Gorissen A. Nonintrusive field experiments show different plant responses to warming and drought among sites, seasons, and species in a north-south European gradient. Ecosystems , 2004, 7 (6) : 598–612. |
| [17] | Sardans J, Peñuelas J, Prieto P, Estiarte M. Drought and warming induced changes in P and K concentration and accumulation in plant biomass and soil in a Mediterranean shrubland. Plant and Soil , 2008, 306 (1/2) : 261–271. |
| [18] | Sardans J, Peñuelas J, Estiarte M, Prieto P. Warming and drought alter C and N concentration, allocation and accumulation in a Mediterranean shrubland. Global Change Biology , 2008, 14 (10) : 2304–2316. DOI:10.1111/gcb.2008.14.issue-10 |
| [19] | Wan S Q, Hui D F, Wallace L, Luo Y Q. Direct and indirect effects of experimental warming on ecosystem carbon processes in a tallgrass prairie. Global Biogeochemical Cycles , 2005, 19 (2) : GB2014. DOI:10.1029/2004GB002315 |
| [20] | Holsten A, Vetter T, Vohland K, Krysanova V. Impact of climate change on soil moisture dynamics in Brandenburg with a focus on nature conservation areas. Ecological Modeling , 2009, 220 (7) : 2076–2087. |
| [21] | Menzel A, Fabian P. Growing season extended in Europe. Nature , 1999, 397 (6721) : 659–659. DOI:10.1038/17709 |
| [22] | Cleland E E, Chiariello N R, Loarie S R, Mooney H A, Field C B. Diverse responses of phenology to global changes in a grassland ecosystem. Proceedings of the National Academy of Sciences of the United States of America , 2006, 103 (37) : 13740–13744. DOI:10.1073/pnas.0600815103 |
| [23] | Tjoelker M G, Reich P B, Oleksyn J. Changes in leaf nitrogen and carbohydrates underlie temperature and CO2 acclimation of dark respiration in five boreal tree species. Plant, Cell & Environment , 1999, 22 (7) : 767–778. |
| [24] | Zha T S, Ryyppö A, Wang K K, Kellomäki S. Effects of elevated carbon dioxide concentration and temperature on needle growth, respiration and carbohydrate status in field-grown Scots pines during the needle expansion period. Tree Physiology , 2001, 21 (17) : 1279–1287. DOI:10.1093/treephys/21.17.1279 |
| [25] | Niu S L, Li Z X, Xia J Y, Wu M Y, Wan S Q. Climatic warming changes plant photosynthesis and its temperature dependence in a temperate steppe of northern China. Environmental and Experimental Botany , 2008, 63 (1/3) : 91–101. |
| [26] | Han C, Liu Q, Yang Y. Short-term effects of experimental warming and enhanced ultraviolet-B radiation on photosynthesis and antioxidant defense of Picea asperata seedlings. Plant Growth Regulation , 2009, 58 (2) : 153–162. DOI:10.1007/s10725-009-9363-2 |
| [27] | Albert K R, Ro-Poulsen H, Mikkelsen T N, Michelsen A, van der Linden L, Beier C. Interactive effects of elevated CO2, warming, and drought on photosynthesis of Deschampsia flexuosa in a temperate heath ecosystem. Journal of Experimental Botany , 2011, 62 (12) : 4253–4266. DOI:10.1093/jxb/err133 |
| [28] | Billings W D, Godfrey P J, Chabot B F, Bourque D P. Metabolic acclimation to temperature in arctic and alpine ecotypes of Oxyria digyna. Arctic and Alpine Research , 1971, 3 (4) : 277–289. DOI:10.2307/1550044 |
| [29] | Smith E M, Hadley E B. Photosynthetic and respiratory acclimation to temperature in Ledum groenlandicum populations. Arctic and Alpine Research , 1974, 6 (1) : 13–27. DOI:10.2307/1550366 |
| [30] | Battaglia M, Beadle C, Loughhead S. Photosynthetic temperature responses of Eucalyptus globulus and Eucalyptus nitens. Tree Physiology , 1996, 16 (1/2) : 81–89. |
| [31] | Badger M R, Björkman O, Armond P A. An analysis of photosynthetic response and adaptation to temperature in higher plants: temperature acclimation in the desert evergreen Nerium oleander L. Plant, Cell & Environment , 1982, 5 (1) : 85–99. |
| [32] | Epron D. Effects of drought on photosynthesis and on the thermotolerance of photosystem Ⅱ in seedlings of cedar (Cedrus atlantica and C. libani). Journal of Experimental Botany , 1997, 48 (5) : 1835–1841. |
| [33] | Berry J, Björkman O. Photosynthetic response and adaptation to temperature in higher plants. Annual Review of Plant Physiology , 1980, 31 : 491–543. DOI:10.1146/annurev.pp.31.060180.002423 |
| [34] | Hikosaka K, Murakamia A, Hirose T. Balancing carboxylation and regeneration of ribulose-1, 5-bisphosphate in leaf photosynthesis: temperature acclimation of an evergreen tree, Quercus myrsinaefolia. Plant, Cell & Environment , 1999, 22 (7) : 841–849. |
| [35] | Crafts-Brandner S J, Law R D. Effect of heat stress on the inhibition and recovery of the ribulose-1, 5-bisphosphate carboxylase/oxygenase activation state. Planta , 2000, 212 (1) : 67–74. DOI:10.1007/s004250000364 |
| [36] | Crafts-Brandner S J, Salvucci M E. Rubisco activase constrains the photosynthetic potential of leaves at high temperature and CO2. Proceedings of the National Academy of Sciences of the United States of America , 2000, 97 (24) : 13430–13435. DOI:10.1073/pnas.230451497 |
| [37] | Onoda Y, Hikosaka K, Hirose T. Seasonal change in the balance between capacities of RuBP carboxylation and RuBP regeneration affects CO2 response of photosynthesis in Polygonum cuspidatum. Journal of Experimental Botany , 2005, 56 (412) : 755–763. DOI:10.1093/jxb/eri052 |
| [38] | Yamori W, Noguchi K, Terashima I. Temperature acclimation of photosynthesis in spinach leaves: analyses of photosynthetic components and temperature dependencies of photosynthetic partial reactions. Plant, Cell & Environment , 2005, 28 (4) : 536–547. |
| [39] | Mitchell R A C, Barber J. Adaptation of photosynthetic electron-transport rate to growth temperature in pea. Planta , 1986, 169 (3) : 429–436. DOI:10.1007/BF00392141 |
| [40] | Yamasaki T, Yamakawa T, Yamane Y, Koike H, Satoh K, Katoh S. Temperature acclimation of photosynthesis and related changes in photosystem Ⅱ electron transport in winter wheat. Plant Physiology , 2002, 128 (3) : 1087–1097. DOI:10.1104/pp.010919 |
| [41] | Farquhar G D, von Caemmerer S. Modelling of photosynthetic response to environment//Lange O L, Nobel P S, Osmond C B, Ziegler H, eds. Encyclopedia of Plant Physiology Ⅱ. Berlin, Heidelberg: Springer, 1982: 549-587. |
| [42] | Kirschbaum M U F, Farquhar G D. Temperature dependence of whole-leaf photosynthesis in Eucalyptus pauciflora Sieb. ex Spreng. Australian Journal of Plant Physiology , 1984, 11 (6) : 519–538. DOI:10.1071/PP9840519 |
| [43] | Wullschleger S D. Biochemical limitations to carbon assimilation in C3 plants-a retrospective analysis of the A/Ci curves from 109 species. Journal of Experimental Botany , 1993, 44 (5) : 907–920. DOI:10.1093/jxb/44.5.907 |
| [44] | Atkin O K, Tjoelker M G. Thermal acclimation and the dynamic response of plant respiration to temperature. Trends in Plant Science , 2003, 8 (7) : 343–351. DOI:10.1016/S1360-1385(03)00136-5 |
| [45] | Armstrong A F, Logan D C, Atkin O K. On the developmental dependence of leaf respiration: responses to short- and long-term changes in growth temperature. American Journal of Botany , 2006, 93 (11) : 1633–1639. DOI:10.3732/ajb.93.11.1633 |
| [46] | Azcón-Bieto J, Osmond C B. Relationship between photosynthesis and respiration: the effect of carbohydrate status on the rate of CO2 production by respiration in darkened and illuminated wheat leaves. Plant Physiology , 1983, 71 (3) : 574–581. DOI:10.1104/pp.71.3.574 |
| [47] | Bolstad P V, Reich P, Lee T. Rapid temperature acclimation of leaf respiration rates in Quercus alba and Quercus rubra. Tree Physiology , 2003, 23 (14) : 969–976. DOI:10.1093/treephys/23.14.969 |
| [48] | Gershenson A, Bader N E, Cheng W X. Effects of substrate availability on the temperature sensitivity of soil organic matter decomposition. Global Change Biology , 2009, 15 (1) : 176–183. DOI:10.1111/gcb.2009.15.issue-1 |
| [49] | Covey-Crump E M, Attwood R G, Atkin O K. Regulation of root respiration in two species of Plantago that differ in relative growth rate: the effect of short- and long-term changes in temperature. Plant, Cell & Environment , 2002, 25 (11) : 1501–1513. |
| [50] | Davidson E A, Janssens I A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature , 2006, 440 (7081) : 165–173. DOI:10.1038/nature04514 |
| [51] | Tao F L, Yokozawa M, Xu Y L, Hayashi Y, Zhang Z. Climate changes and trends in phenology and yields of field crops in China, 1981–2000. Agricultural and Forest Meteorology , 2006, 138 (1/4) : 82–92. |
| [52] | Lin E D. Agricultural vulnerability and adaptation to global warming in China. Water, Air, and Soil Pollution , 1996, 92 (1/2) : 63–73. |
| [53] | Smit B, Cai Y L. Climate change and agriculture in China. Global Environment Change , 1996, 6 (3) : 205–214. DOI:10.1016/0959-3780(96)00018-0 |
| [54] | Chen L X, Zhu W Q, Wang W, Zhou X J, Li W L. Studies on climate change in China in recent 45 years. Acta Meteorologica Sinica , 1998, 12 (1) : 1–17. |
| [55] | Mo X G, Liu S X, Lin Z H, Guo R P. Regional crop yield, water consumption and water use efficiency and their responses to climate change in the North China Plain. Agriculture, Ecosystem & Environment , 2009, 134 (1/2) : 67–78. |
| [56] | Liu S X, Mo X G, Lin Z H, Xu Y Q, Ji J J, Wen G, Richey J. Crop yield responses to climate change in the Huang-Huai-Hai Plain of China. Agriculture Water Management , 2010, 97 (8) : 1195–1209. DOI:10.1016/j.agwat.2010.03.001 |
| [57] | Tao F L, Zhang S, Zhang Z. Spatiotemporal changes of wheat phenology in China under the effects of temperature, day length and cultivar thermal characteristics. European Journal of Agronomy , 2012, 43 : 201–212. DOI:10.1016/j.eja.2012.07.005 |
| [58] | Hou R X, Ouyang Z, Li Y S, Wilson G V, Li H X. Is the change of winter wheat yield under warming caused by shortened reproductive period. Ecology and Evolution , 2012, 2 (12) : 2999–3008. DOI:10.1002/ece3.403 |
| [59] | Lobell D B, Field C B. Global scale climate-crop yield relationships and the impacts of recent warming. Environment Research Letter , 2007, 2 : 014002. DOI:10.1088/1748-9326/2/1/014002 |
| [60] | Sharkey T D, Bernacchi C J, Farquhar G D, Singsaas E L. Fitting photosynthetic carbon dioxide response curves for C3 leaves. Plant, Cell & Environment , 2007, 30 (9) : 1035–1040. |
| [61] | Wong S C. Elevated atmospheric partial pressure of CO2 and plant-growth. Ⅱ: Non-structural carbohydrate content in cotton plants and its effect on growth parameters. Photosynthesis Research , 1990, 23 (2) : 171–180. DOI:10.1007/BF00035008 |
| [62] | Hoch G, Richter A, Körner C. Non-structural carbon compounds in temperate forest trees. Plant, Cell & Environment , 2003, 26 (7) : 1067–1081. |