 2016, Vol. 36
2016, Vol. 36文章信息
- 韩瑞, 陈求稳, 王丽, 汤新武, 沈新强
- HAN Rui, CHEN Qiuwen, WANG Li, TANG Xinwu, SHEN Xinqiang.
- 基于生态通道模型的长江口水域生态系统结构与能量流动分析
- Analysis of the ecosystem structure and energy flow of the Yangtze River estuary and adjacent seas, based on the Ecopath model
- 生态学报[J]. 2016, 36(15): 4907-4918
- Acta Ecologica Sinica[J]. 2016, 36(15): 4907-4918
- http://dx.doi.org/10.5846/stxb201501150129
-
文章历史
- 收稿日期: 2015-01-15
- 网络出版日期: 2015-11-16





2. 南京水利科学院, 南京 225600;
3. 三峡大学, 宜昌 443002;
4. 中国水产科学研究院东海水产研究所, 上海 200091
2. Nanjing Hydraulic Research Institute, Nanjing 225600, China;
3. Three Gorges University, Yichang 443002, China;
4. East China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Shanghai 200091, China
长江口水域咸淡水交汇,环境条件复杂多变,理化和生物特征特殊。流域内水利工程众多,运行叠加效应在空间上远达长江口及东海[1-3],引起河口水域的水文、水化学和沉积环境变化,进而改变河口生物群落的结构、分布及生产力[1-2],使得长江口成为一个结构独特、功能多样的生态系统。三峡工程是长江流域最大的控制工程,自兴建便成为公众焦点。2003年6月10日三峡蓄水水位达135m高程(一期蓄水),2006年6月,蓄水位抬高至156m(二期蓄水),2009年9月,蓄水位至175m(正常蓄水位)。工程建成后,长江年径流总量略微增加[4],月均径流量受调控影响显著,蓄水期长江入海径流量减少约18%[5],一期蓄水后,长江入海泥沙通量仅为1968年前的37%[6],水下三角洲被侵蚀,-10m等深线后缩1—2km[7-8]。同时,长江口渔业捕捞生产也对生态系统产生了不可忽视的影响。本研究综合了长江口渔业捕捞的影响,从生态系统的整体角度出发,重点分析了三峡工程蓄水关键时期的生态系统状态,量化评估[9]三峡工程对长江口生态系统结构与能量流动的影响。
基于生态系统的分析方法和决策思想[10],在已有研究[11]的基础上,进一步调查了三峡工程蓄水后(2012年秋)主要物种的生物量、捕捞量等,分别构建了三峡工程关键时期(蓄水前—2000年秋、蓄水中,即156m蓄水位—2006年秋、蓄水后,即175m蓄水位—2012年秋)对应长江口水域生态通道模型,对比分析了不同时期生态系统的营养级构成、能量流动以及生态系统的总体特征,探讨长江口生态系统结构和功能的改变,为量化评价三峡工程对长江口生态系统的影响提供参考。
1 研究区域介绍与数据收集 1.1 研究区域概况长江口是极为重要的生态交错区,区内理化环境因子复杂多变,生物群落的生态类型多样,生态系统具有生产力水平高、环境梯度变化大等特点。同时,长江口是众多鱼类的育幼场和洄游性鱼类的洄游通道[12-13]。三峡工程蓄水后,长江冲淡水和最大混合区出现季节性萎缩,悬沙量显著减少[14],导致该区域的营养盐浓度和结构产生变异:溶解态无机氮、磷酸盐含量呈增长趋势,硅酸盐含量呈降低趋势,氮磷比、硅氮比、硅磷比均有不同程度的降低[15-16],浮游动物生物量增长而丰度降低[17],水母类比例持续升高[18]。总体而言,三峡工程对长江口产生的直接影响首先表现在化学组成和含沙量变化上,随后,通过生态阀门反馈或叠加,从而影响生物群落乃至生态系统[2]。
1.2 数据来源蓄水前(2000年秋)中(2006年秋)期的数据来源于中国水产科学研究院黄海水产研究所“北斗”号科学调查船在长江口及临近海域(122°—125°E, 29°—34°N)的调查,范围如图 1所示。蓄水后期(2012年秋)数据来源于中国水产科学研究院东海水产研究所,调查从长江口自西向东涵盖4个区域:河道、最大浑浊带、河口东部-10 m等深线、-20 m等深线,调查区域(121°—125°E, 30°—32°N)如图 1所示。本研究针对蓄水不同时期调查范围存在一定差异,在模型建立过程中选取重叠的区域,并参考了临近水域的研究结果[18-22]。同时,依据生态系统质量守恒原理及对应年份《中国渔业统计年鉴》[23-24]中上海地区海洋捕捞产量中甲壳类(虾、蟹)、头足类等渔获量对模型中对应功能组的渔获量进行对照。根据“简化食物网”[25]对2012年秋季的渔获物进行生物学测定和胃含物分析,估算生物量、生产量/生物量、消耗量/生物量、渔获量等参数,基本满足建模数据需求。

|
| 图 1 长江口生态调查调查区域 Fig. 1 The ecological surveys and sampling areas in Yangtze River estuary |
生态通道模型(Ecopath)在评估稳态生态系统的基础上[26],结合能量分析生态学[27]发展形成的基于营养动力学[28]的模型,可构造水生态系统结构、描述能量流动及定量分析生态系统特征参数[29]。Ecopath模型定义生态系统由一系列生态关联的功能组组成,功能组可以为浮游生物、某一种鱼类、某种鱼类某一年龄组或一类生态特性(如摄食)相似的鱼类等,所有功能组能够基本涵盖生态系统中能量流动的全部过程。
Ecopath模型有两组控制方程,第一组方程(公式1)主要描述生态系统各功能组的质量守恒,第二组方程(公式2)主要描述各功能组的能量平衡:
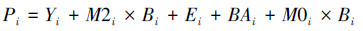 (1)
(1)
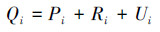 (2)
(2)
式中, Pi为功能组i的生产率;Yi为功能组i的捕获率;M2i为功能组i的瞬时捕食死亡率,可通过公式(3)计算;Bi为生物量;Ei为相应功能组的净迁移率;BAi为功能组i的生物量累积率;M0i为功能组i的其它死亡率,可通过公式(4)计算;Qi为功能组i的消耗量;Ri为功能组i的呼吸量;Ui为功能组i未消化的食物量。
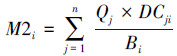 (3)
(3)
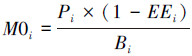 (4)
(4)
式中, EEi为功能组i的生态营养转换率,指生产量在生态系统中的利用比例;DCji为被捕食者j占捕食者i的食物组成比例,一般根据生物的胃含物分析获得。
公式(1)可进一步表达为:
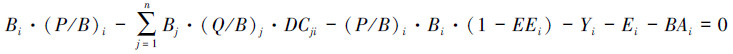 (5)
(5)
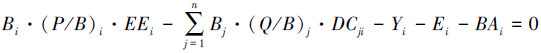 (6)
(6)
式中, (P/B)i为功能组i的生产量与生物量比值,通常可用瞬时总死亡率代替[30];(Q/B)i为功能组i的消耗量与生物量的比值,可参考鱼类尾鳍外形比的多元回归模型计算[31]。
2.2 功能组划分Ecopath模型中功能组的划分主要根据研究需求以及相应的生态学和生物分类学理论,通常需要符合以下原则:①将生态位重叠度高的种群合并,如将具有相同或类似的食物构成、个体大小等特征的生物种群划分为一组;②至少包括1个有机碎屑功能组;③保证功能组的完整性,如优势种。
根据以上原则,本研究综合考虑研究目的、水域现状及“简化食物网”的研究策略[25],且为将不同时期生态系统模型进行比较,参照了长江口生物调查[32]和已建立模型[11]的功能组划分,将2012年秋季调查结果划分为17个功能组,基本涵盖生态系统能量流动的全过程。17个功能组分别为:鱼食性鱼类、广食性鱼类、虾食性鱼类、浮游动物食性鱼类、虾或蟹食性鱼类、其它中上层鱼类、其它底层鱼类、其它底栖鱼类、虾、蟹、头足类、大型底栖动物、小型底栖动物、水母类、浮游植物、浮游动物、碎屑。不同时期主要鱼类功能组及对应物种见表 1。
| 功能组Functional group | 2000年秋 The autumn of 2000 |
2006年秋 The autumn of 2006 |
2012年秋 The autumn of 2012 |
| 鱼食性鱼类Piscivorous fishes | 小黄鱼、竹荚鱼、鲐 | 小黄鱼、竹荚鱼 | 半滑舌鳎、小黄鱼 |
| 广食性鱼类Generalist predators | 叫姑鱼、白姑鱼 | 黄鲫 | 花鲢、叫姑鱼 |
| 虾食性鱼类Shrimp predators | 矛尾虾虎鱼 | 矛尾虾虎鱼 | 矛尾虾虎鱼 |
| 浮游动物食性鱼类Zooplanktivores | 发光鲷、七星底灯鱼 | 刺鲳、鳀 | 凤鲚、斑鰶、刀鲚 |
| 虾/蟹食性鱼类Shrimp/crab predators | 多棘腔吻鳕 | 虻鲉 | 孔鳐 |
模型要求输入生物量(B)、生产量/生物量(P/B)、消耗量/生物量(Q/B)和生态营养转换效率EE这4组基本参数中的任意3组,以及食物组成矩阵DC和渔获量Y。
(1)生物量B
模型中每个功能组的生物量均为研究区域内的平均值(t/km2)。浮游植物的生物量通过长江口叶绿素含量平均值转化而得。浮游动物、底栖动物等功能组的生物量由调查直接获得[33]。本研究中水母和碎屑的生物量数据缺乏且较难估算,在此根据相邻水域的生态通道模型[34]进行计算,这也是生态通道模型建模中允许的,即对缺乏参数的功能组可参考邻近海域生态通道模型中相同或相似功能组参数。鱼类和头足类的生物量数据通过采样调查获得,转化公式如下:
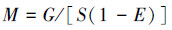 (7)
(7)
式中, M为生物量,G为调查生物量,S为网口的扫海面积,E为逃逸率。
(2)生产量/生物量(P/B)
P/B为生产量与平均生物量之间的比值,即生物量周转率,标准单位为1/a[35],可根据调查具体情况进行调整,本研究为对应年份秋季。由于鱼类的P/B与总死亡率Z之间存在P/B=Z的关系[30],所以可通过鱼类总死亡率Z而得到P/B值,可通过下式估算[36]:
 (8)
(8)
式中, tmax为调查功能群中的最大年龄。
(3)消耗量/生物量(Q/B)
Q/B为某种生物摄食量与生物量的比值,标准单位为1/year,可根据调查具体情况进行调整,本研究为对应年份秋季。模型中各鱼类功能组中主要根据鱼类种类的生长方程等资料获得。
(4)生态营养转换效率EE
EE指生物在生态系统中用于被捕食和输出的部分,取值在0—1范围内。在长江口生态通道模型中,水母的EE值是直接输入的,其他功能组的EE值通过模型计算获得。
(5)食物组成矩阵DC
食物组成数据来自长江口历史调查[32, 37-38]、胃含物分析及fishbase(www.fishbase.org)的相关信息。
2.4 模型调试与敏感性分析生态通道模型是稳态模型,模型内部各功能组都必须同时达到物质和能量的双重平衡。模型的调试过程即在客观规律和可靠经验的基础上,反复调整修正输入参数,最小化不合理因素,从而在模型达到最终平衡状态的同时能够最真切地反映生态系统实际状况。在模型输入参数中,生态营养转换效率是一个较难获得的参数,通常设为未知,通过模型计算获得。因此,调试首先考虑各功能组都要保证0 < EE < 1,使能量在生态系统中流动保持平衡,从而获得系统其它生态学参数的合理值。此外,0.1 < P/Q < 0.3,同时各功能组的呼吸量必须大于0。鉴于在所有的输入参数中,有关功能组食物组成信息准确度相比较其它参数而言不高,长江口生态通道模型调试主要从食物组成着手,对各功能组的食物构成做出调整,模型的敏感度在±20%范围内。调试平衡后参数设置如表 2。
| 功能组Function group | 2000年秋The autumn of 2000 | 2006年秋The autumn of 2006 | 2012年秋The autumn of 2012 | ||||||||||||||||
| 编号No | 名称Name | TL | B | P/B | Q/B | EE | Y | TL | B | P/B | Q/B | EE | Y | TL | B | P/B | Q/B | EE | Y |
| 1 | 鱼食性鱼类Piscivorous fishes | 3.802 | 0.586 | 0.758 | 4.400 | 0.997 | 0.404 | 4.058 | 0.140 | 0.800 | 4.500 | 0.994 | 0.100 | 4.019 | 0.222 | 0.800 | 4.500 | 0.999 | 0.176 |
| 2 | 广食性鱼类Generalist predators | 3.634 | 0.0176 | 0.974 | 7.300 | 0.992 | 0.017 | 3.850 | 0.021 | 0.850 | 4.400 | 0.998 | 0.012 | 3.847 | 0.015 | 0.910 | 7.000 | 0.989 | 0.012 |
| 3 | 虾食性鱼类Shrimp predators | 3.463 | 0.0243 | 0.860 | 5.500 | 0.866 | 0.006 | 3.776 | 0.105 | 0.850 | 4.500 | 0.879 | 0.035 | 3.776 | 0.180 | 0.860 | 5.000 | 0.987 | 0.052 |
| 4 | 浮游动物食性鱼类Zooplanktivores | 3.181 | 0.925 | 2.100 | 8.000 | 0.907 | 0.012 | 3.516 | 0.276 | 0.700 | 5.000 | 0.962 | 0.120 | 3.513 | 0.130 | 1.000 | 6.000 | 0.994 | 0.029 |
| 5 | 虾/蟹食性鱼类Shrimp/crab predators | 3.627 | 0.210 | 1.300 | 4.500 | 0.939 | 0.221 | 3.846 | 0.066 | 1.200 | 6.000 | 0.990 | 0.062 | 3.621 | 0.156 | 0.870 | 6.000 | 0.995 | 0.127 |
| 6 | 其它中上层鱼类Other pelagic fishes | 2.699 | 0.721 | 1.400 | 4.700 | 0.916 | 0.139 | 2.974 | 0.691 | 0.808 | 4.500 | 0.997 | 0.150 | 2.970 | 0.314 | 0.930 | 4.500 | 0.982 | 0.009 |
| 7 | 其它底层鱼类Other demersal fishes | 3.153 | 0.192 | 0.800 | 5.000 | 0.944 | 0.015 | 3.316 | 0.208 | 0.879 | 4.950 | 0.986 | 0.020 | 3.274 | 0.303 | 0.900 | 5.000 | 0.965 | 0.026 |
| 8 | 其它底栖鱼类Other benthic fishes | 3.252 | 0.0626 | 0.950 | 4.800 | 0.835 | 0.028 | 3.152 | 0.010 | 0.958 | 4.933 | 0.961 | 0.040 | 3.152 | 0.136 | 0.900 | 5.000 | 0.998 | 0.006 |
| 9 | 虾Shrimps | 2.237 | 0.264 | 5.400 | 28.00 | 0.954 | 0.013 | 2.701 | 0.252 | 5.600 | 28.90 | 0.975 | 0.150 | 2.701 | 0.266 | 4.090 | 28.00 | 0.996 | 0.033 |
| 10 | 蟹Crabs | 2.730 | 0.128 | 3.000 | 12.00 | 0.838 | 0.041 | 2.884 | 0.465 | 3.500 | 12.00 | 0.929 | 0.110 | 2.883 | 0.386 | 4.500 | 15.00 | 0.978 | 0.041 |
| 11 | 头足类Cephalopod | 3.390 | 0.170 | 3.000 | 10.00 | 0.976 | 0.040 | 3.252 | 0.382 | 3.100 | 8.000 | 0.949 | 0.160 | 3.232 | 0.300 | 3.100 | 11.97 | 0.800 | 0.048 |
| 12 | 大型底栖动物Macrobenthos | 2.257 | 8.790 | 2.000 | 8.600 | 0.305 | / | 2.224 | 3.800 | 2.000 | 8.600 | 0.452 | / | 2.224 | 3.149 | 2.000 | 8.600 | 0.661 | / |
| 13 | 小型底栖动物Meiobenthos | 2.000 | 4.723 | 9.280 | 33.00 | 0.114 | / | 2.045 | 3.373 | 9.280 | 33.00 | 0.450 | / | 2.045 | 1.392 | 9.280 | 33.00 | 0.851 | / |
| 14 | 水母类Jellyfish | 2.731 | 0.765 | 5.000 | 20.00 | 0.320 | 0.090 | 2.328 | 2.026 | 5.000 | 20.00 | 0.350 | 0.020 | 2.328 | 1.785 | 5.000 | 20.00 | 0.350 | 0.110 |
| 15 | 浮游动物Zooplankton | 2.029 | 3.935 | 25.00 | 180.0 | 0.536 | / | 2.053 | 4.013 | 25.00 | 180.0 | 0.535 | / | 2.053 | 5.000 | 25.00 | 180.0 | 0.484 | / |
| 16 | 浮游植物Phytoplankton | 1.000 | 18.98 | 180.0 | - | 0.188 | / | 1.000 | 5.604 | 200.0 | - | 0.444 | / | 1.000 | 7.000 | 200.0 | - | 0.439 | / |
| 17 | 碎屑Detritus | 1.000 | 15.00 | - | - | 0.072 | / | 1.000 | 18.00 | - | - | 0.401 | / | 1.000 | 18.00 | - | - | 0.314 | / |
| ①TL:营养级Trophic Level,B:生物量biomass(t/km2),P/B:生产量/生物量Production/Biomass,Q/B:消耗量/生物量Consumption/Biomass,EE:生态营养转换效率Ecotrophic efficiency,Y: 渔获量(t/km2);②黑体数据为生态通道模型估算结果Bold parameters were estimated by Ecopath models;③-相应值不存在means the corresponding value did not exist,/无统计数据means no statistical data | |||||||||||||||||||
长江口水域生态通道模型的部分计算结果见表 2,可以看出:①3个时期生态系统模型中,各功能组所处营养级符合生态学规律,即第一营养级是自养营养级,第二至第五营养级是异养营养级,植物性浮游动物位于第二营养级(2级),低级肉食性鱼类为第三级(3.0—3.5),中级肉食性鱼类为第四级(3.6—4.0),高级肉食性鱼类为第五级(4.0—4.5)[33]。且各功能组所处营养级分布近似,如3个时期,鱼食性鱼类均处于对应时期生态系统营养级上层。在构建长江口水域模型时,暂未考虑哺乳动物对生态系统的影响;②蓄水前中后期对应生态系统模型中,虾食性、浮游动物食性、虾/蟹食性鱼类、虾、蟹等功能组的生态营养转换效率有升高趋势,主要可能由于长江口水域渔业资源过度捕捞造成,如定置网具大量杀伤幼鱼、深水张网等无选择作业不限制投产网数和作业时间等[39]。但头足类的生态营养转换效率逐渐降低,主要由于头足类生物量在蓄水中后期均有所增加,而渔获量在蓄水中期显著增加,后期降低至蓄水前水平。
生态系统总能流量、生物量和渔获量在各营养级的分布结果(表 3)表明:①不同时期生态系统的生物量和能量流动量均呈金字塔型分布,即低营养级的生物量、能量流动量大,越到顶级越小,基本符合能流量和生物量的分布规律;②蓄水前能流量普遍高于蓄水中后期,蓄水中期(2006)能量流动普遍低于蓄水后;③不同时期生态系统的生物量呈现逐渐减少的趋势;④渔获量营养级结构未发生明显变化,主要渔获量分布在第III、IV营养级,渔获量随蓄水的进行逐渐减少。
| 营养级 Trophic level |
总能流量/ (t km-2 a-1) Throughput |
生物量/ (t km-2 a-1) Biomass |
渔获量/ (t km-2 a-1) Catch |
||||||
| 2000 | 2006 | 2012 | 2000 | 2006 | 2012 | 2000 | 2006 | 2012 | |
| Ⅰ | 6486 | 1121 | 2466 | 33.98 | 23.60 | 25.00 | - | - | - |
| Ⅱ | 913.7 | 498.0 | 950.0 | 16.28 | 12.41 | 10.55 | 0.134 | 0.174 | 0.104 |
| Ⅲ | 41.20 | 13.91 | 30.34 | 4.369 | 2.676 | 2.416 | 0.454 | 0.487 | 0.329 |
| Ⅳ | 4.345 | 1.469 | 4.161 | 0.765 | 0.728 | 0.667 | 0.381 | 0.272 | 0.235 |
| Ⅴ | 0.477 | 0.187 | 0.486 | 0.090 | 0.090 | 0.0913 | 0.053 | 0.041 | 0.047 |
长江口生态系统能量流动和转换通过牧食食物链和碎屑食物链共同实现,不同时期生态系统能量流动主要发生在7个营养级间,且主要发生在低营养级。从表 4及图 2看出:蓄水中后期营养级的能量流分布极为相似,碎屑和浮游植物占据生态系统营养级底层,且变化较小;在较高层营养级别的能流分布在蓄水不同时期变化较大,主要集中在Ⅳ、Ⅴ级,说明生态系统食物网结构与蓄水前相比,在营养级中的分布上升,而食物网结构的改变影响了整个生态系统的能量流动分布状况。
| 营养级 Trophic level |
2000年秋The autumn of 2000 | 2006年秋The autumn of 2006 | 2012年秋The autumn of 2012 | |||
| 初级生产者/% Primary producer |
碎屑/% Detritus |
初级生产者/% Primary producer |
碎屑/% Detritus |
初级生产者/% Primary producer |
碎屑/% Detritus |
|
| Ⅱ | 4.7 | 4.6 | 2.8 | 5.2 | 2.0 | 5.4 |
| Ⅲ | 10.1 | 16.2 | 11.9 | 16.7 | 16.6 | 16.6 |
| Ⅳ | 19.9 | 19.6 | 18.2 | 17.5 | 17.3 | 17.3 |
| Ⅴ | 19.3 | 19.5 | 18.1 | 18.9 | 17.4 | 17.8 |
| Ⅵ | 19.2 | 19.4 | 19.3 | 17.8 | 17.5 | 17.2 |
| Ⅶ | 19.0 | - | 18.0 | 19.8 | 17.5 | 17.6 |
| 转换率Transfer efficiency | 9.8 | 11.4 | 8.5 | 11.5 | 7.5 | 11.6 |
| 总转换率 Total transfer efficiency |
10.2 | 10.0 | 9.4 | |||

|
| 图 2 2000年秋、2006年秋和2012年秋长江口生态系统能量流动结构图 Fig. 2 Lindeman Spines in the Yangtze River estuary ecosystem in autumns of 2000, 2006 and 2012 |
蓄水前,牧食食物链在系统能量流动中占优势,能量通过牧食食物链传递56%,通过碎屑食物链传递44%,平均总转换率为10.2%;蓄水中,牧食食物链在系统能量流动中具有优势,传递56%能量,通过碎屑食物链传递44%,平均总转换率为10.0%,较蓄水前略降低;蓄水后,牧食食物链在系统能量流动中的优势增强,能量通过牧食食物链传递59%,通过碎屑食物链传递41%,平均总转换率略有下降,为9.4%。3个时期能量分布主要集中在Ⅰ—Ⅱ级,占全部能量的90%以上,营养流随营养层次的升高而减少,越高层次的营养级能量流动分布越少,在Ⅳ—Ⅴ级的流动甚少。
3.3 混合营养效应及关键功能组分析生态通道模型中混合营养影响可以定量评价所有功能组间直接或间接的营养影响[40],分析各功能组的生物量增加对其它功能组的影响[41]。不同时期长江口生态系统中各功能组对自身的影响均为负向影响(同种生物对同类资源的种内竞争),被捕食者对捕食者存在正向影响,作为生态系统初级生产力的碎屑和浮游植物对各功能组的影响趋于正向。
关键功能组是指生态系统食物网中生物量相对较少但对系统结构具有重大影响的功能组,主要基于混合营养影响分析获得。图 3表明不同时期长江口生态系统的关键功能组分别为浮游动物食性鱼类(蓄水前)、鱼食性鱼类(蓄水中后期),长江口生态系统中高营养级鱼类通过食物资源生态位分化作用于低营养级功能组,如浮游动物、低营养级鱼类等,从而影响了生态系统的发展。蓄水前,浮游动物食性鱼类为最关键功能组,鱼食性鱼类、虾、头足类、浮游动物均处于较重要的位置;蓄水中后期,鱼食性鱼类为最关键功能组,但蓄水中期处于较关键功能组的虾、蟹在后期关键度降低。研究长江口生态系统的上下行效应对控制长江口赤潮爆发有重要意义,宋洪军等依托本调查,研究了长江口浮游植物上下行控制作用,得出营养盐对浮游植物具有上行效应,浮游动物下行抑制作用不明显[42],但其他研究[43]表明部分区域(122°E、31°N附近)高等生物的下行摄食调节机制仍有一定作用。

|
| 图 3 2000年秋、2006年秋和2012年秋长江口生态系统关键功能组 Fig. 3 Keystones in the Yangtze River estuary ecosystem in autumns of 2000, 2006 and 2012 |
河口生态系统受人类高强度活动干扰,结构复杂、生产力较高[44],生态通道模型可在系统生态学基础上提出具体的量化特征参数。从表 5可看出:①生态系统最重要的指标是总初级生产量/总呼吸量(TPP/TR)[45]。在系统发育早期,生产量会超过呼吸量,因此TPP/TR>1;成熟系统中,TPP/TR≈1。由此判断,蓄水前中后期长江口生态系统均处于不成熟阶段;②系统总流量(Total throughput)是表征系统规模的指标,是总摄食量、总输出量、总呼吸量和流入有机碎屑量的总和,长江口生态系统的总流量在蓄水中期显著下降,后期略有上升,但当中流向有机碎屑的流量逐渐下降,约有40.85%(前)、30.11%(中)、30.47%(后)的物质未被生物摄食、吸收,而转化为有机碎屑,蓄水中后期系统规模较蓄水前明显减少,能量流向碎屑的比例持续降低,碎屑食物链的重要性下降,牧食食物链的重要性上升;③系统的总生物量主要由系统中可利用的能流所维持,在成熟的系统中,总生物量/总流量(TB/T)的数值一般较大,不同时期长江口生态系统的TB/T值均很低,同样反映出了系统的不成熟;④系统连接指数(CI)和系统杂食指数(SOI)反映了系统内复杂程度,越成熟越复杂。长江口生态系统的CI和SOI反应了生态系统内部结构一直较为简单,生物多样性水平不高,系统相对不成熟。⑤循环指数(FCI)指系统中通过碎屑路径循环流量与总流量的比值,平均路径长度(MPL)指的是每个循环流经食物链的平均长度。在成熟的生态系统里,物质再循环的比例通常较高且营养流所经过的食物链较长,而在长江口生态系统在蓄水前中后期均体现出了不成熟的特点,蓄水中后期生态系统在系统流量规模上明显小于蓄水前,但食物网复杂性及系统成熟性方面均优于蓄水前;⑥长江口渔业结构变化不明显,仍维持在较高营养级水平,但蓄水中后期总渔获量明显下降,蓄水后期渔获量比蓄水前缩减了29.92%。
| 名称Name | 参数Parameters | 单位Units | 2000年秋 The autumn of 2000 |
2000年秋 The autumn of 2000 |
2012年秋 The autumn of 2012 |
| 系统总能流量Total throughput | TST | t/km2 | 7517.423 | 2935.707 | 3498.801 |
| 总生物量Total biomass | TB | t/km2 | 40.491 | 21.515 | 20.733 |
| 渔获物平均营养级Mean trophic level of catch | - | - | 3.406 | 3.240 | 3.369 |
| 总渔获量Total catch | t/km2 | 1.026 | 0.979 | 0.719 | |
| 总初级生产量Total primary production | TPP | t/km2 | 3415.770 | 1120.50 | 1400.00 |
| 总初级生产量/总呼吸量Total primary production/ total respiration | TPP/TR | - | 5.551 | 1.899 | 2.095 |
| 总生物量/总能流量Total biomass/ Total throughput | TB/T | - | 0.005 | 0.007 | 0.006 |
| 循环指数Finn Cycling Index | FCI | % | 1.63 | 7.26 | 5.99 |
| 平均能量流动路径Finn′s mean path length | MPL | - | 2.169 | 2.619 | 2.500 |
| 连接指数Connectance Index | CI | - | 0.359 | 0.371 | 0.371 |
| 系统杂食指数System Omnivory Index | SOI | - | 0.179 | 0.205 | 0.196 |
本研究将蓄水前后不同时期计算结果与长江流域和附近海域的其它生态通道模型计算结果进行对比(表 6),从时间(1960年—2012年)和空间(长江流域、东海、黄海)上,对长江口生态系统的演替过程进行进一步分析。
| 生态系统Ecosystem | 年份Year | TPP/TR | SOI | FCI/% | MPL | CI |
| 长江口The Yangtze River estuary | 2000 | 5.551 | 0.179 | 1.63 | 2.169 | 0.359 |
| 2006 | 1.899 | 0.205 | 7.26 | 2.619 | 0.371 | |
| 2012 | 2.095 | 0.196 | 5.99 | 2.50 | 0.371 | |
| 东海南部[34] The Southern East China Sea | 1999—2002 | 3.060 | 0.213 | 4.100 | 2.398 | 0.330 |
| 东海[18] The East China Sea | 1997—2000 | 3.38 | 0.2 | 0.18 | - | 0.19 |
| 太湖[22] The Taihu Lake | 1960 | 23.735 | - | 6.86 | 2.464 | 0.186 |
| 2002 | 5.457 | - | 15.49 | 3.428 | 0.215 | |
| 黄海[20] The Yellow Sea | 2000—2001 | 1.42 | 0.21 | 9.83 | - | 0.36 |
| -相应值在原文中未列出 | ||||||
分析表明:①长江口及附近海域生态系统的TPP/TR值均大于1,生态系统长期处于发展阶段,尚未出现南海北部湾生态系统的逆行演替现象[46];②SOI值变幅较小(0.17—0.32),且随时间有减少趋势,生态位宽度渐窄;③FCI值虽变幅较大(0.18—15.5),但仍呈增大趋势,表示系统流量利用循环效率和系统的成熟度、稳定性有所上升,生态系统抵抗外力干扰的能力有所增强;④MPL变幅为2.16—3.43、CI变幅为0.18—0.38,生态系统中营养流经食物链较短,系统内部联系复杂程度较低,生态系统处于不成熟状态。
4 结论长江口处于长江与东海海陆交互作用区域,来自陆源的物质和长江水动力的变化对河口生态系统的发育和维持有极显著的影响,自20世纪50年代起,在长江口水域已有众多调查,但调查区域重叠较少,时间连续性较弱(仅1985—1986年1次/月),为长江口水域的长期研究带了一定的困难。本研究在已有研究及相关调查的基础上,通过建立三峡工程蓄水前中后不同时期长江口生态系统的生态通道模型,计算生态系统重要的状态参数,量化长江口生态系统结构、功能及能量流动等,得出如下结论:
(1)三峡工程蓄水前中后各功能组的营养级分布相近,但蓄水后功能组生态营养转换率普遍提高,这可能是由于长江口面临的捕捞压力逐年增加,较高营养层次转换成终极产品的转换率被迫提高;
(2)长江口渔获物结构在不同时期变化不明显,但渔获物平均营养级降低,渔获量也有所减少,渔业资源有向小型化、低值化发展的趋势。近年来,由于长江流域经济的迅速发展,大量污染物流至河口,不断加重河口污染,导致富营养化日趋严重,这可能是渔业资源变化的原因之一[2];
(3)三峡工程蓄水后,长江入海径流量减少约18%[5],长江冲淡水面积缩减[47],陆源营养物质和悬浮颗粒的输入减少,海水透明度升高,促进了藻类等浮游生物的生长,使得生态系统的能量传递更依赖牧食食物链,造成大部分剩余能量在系统中沉积,增大系统内源性污染的风险;
(4)长江口生态系统在蓄水前中后均处于不成熟阶段,蓄水后系统总能流量、总生产量、初级生产量及流向碎屑的能量均有所降低,但系统的净效率和再循环率升高,能量在系统中流动的路径增多、营养交互关系复杂化,对外界扰动的适应性增强,生态系统总体处于发育阶段。
致谢: 国家海洋局第一海洋研究所和中国水产科学研究院东海水产研究所给予支持,浙江省杭州工业大学王颖老师和山东省渔业资源与生态环境重点实验室林群老师给予帮助,特此致谢。| [1] | 刘瑞玉, 罗秉征. 三峡工程对长江口生态和渔业的影响[J]. 水土保持通报 , 1987, 7 (4) : 37–40. |
| [2] | 线薇薇, 刘瑞玉, 罗秉征. 三峡水库蓄水前长江口生态与环境[J]. 长江流域资源与环境 , 2004, 13 (2) : 119–124. |
| [3] | 翟世奎, 孟伟, 于志刚. 三峡工程一期蓄水后的长江口海域环境 , 2008 . |
| [4] | Yang Z, Wang H, Saito Y, Milliman J D, Xu K, Qiao S, Shi G. Dam impacts on the Changjiang (Yangtze) River sediment discharge to the sea: The past 55 years and after the Three Gorges Dam[J]. Water Resources Research , 2006, 42 : W04407. DOI:10.1029/2005WR003970 |
| [5] | Wang X L, Wang B D, Zhang C S, Shi X Y, Zhu C J, Xie L P, Han X R, Xin Y, Wang J G. Nutrient composition and distributions in coastal waters impacted by the Changjiang plume[J]. Acta Oceanologica Sinica , 2008, 27 (5) : 111–125. |
| [6] | Yang S L, Milliman J D, Li P, Xu K. 50'000 dams later: Erosion of the Yangtze River and its delta[J]. Global and Planetary Change , 2011, 75 (1/2) : 14–20. |
| [7] | Yang S L, Belkin I M, Belkina A I, Zhao Q Y, Zhu J, Ding P X. Delta response to decline in sediment supply from the Yangtze River: evidence of the recent four decades and expectations for the next half-century[J]. Estuarine, Coastal and Shelf Science , 2003, 57 (4) : 689–699. DOI:10.1016/S0272-7714(02)00409-2 |
| [8] | Wang Z H, Li L Q, Chen D C, Xu K Q, Wei T Y, Gao J H, Zhao Y W, Chen Z Y, Masabate W. Plume front and suspended sediment dispersal off the Yangtze (Changjiang) River mouth, China during non-flood season[J]. Estuarine, Coastal and Shelf Science , 2007, 71 (1/2) : 60–67. |
| [9] | 宋兵, 陈立侨, ChenYong. Ecopath with Ecosim在水生生态系统研究中的应用[J]. 海洋科学 , 2007, 31 (1) : 83–86. |
| [10] | 米玮洁, 胡菊香, 赵先富. Ecopath模型在水生态系统评价与管理中的应用[J]. 水生态学杂志 , 2012, 33 (1) : 127–130. |
| [11] | 林群, 金显仕, 郭学武, 张波. 基于Ecopath模型的长江口及毗邻水域生态系统结构和能量流动研究[J]. 水生态学杂志 , 2009, 2 (2) : 28–36. |
| [12] | 李建生, 李圣法, 程家骅. 长江口渔场拖网渔业资源利用的结构分析[J]. 海洋渔业 , 2004, 26 (1) : 24–28. |
| [13] | Blaber S J M, Cyrus D P, Albaret J J, Ching C V, Day J W, Elliott M, Fonseca M S, Hoss D E, Orensanz J, Potter I C, Silvert W. Effects of fishing on the structure and functioning of estuarine and nearshore ecosystems[J]. ICES Journal of Marine Science , 2000, 57 (3) : 590–602. DOI:10.1006/jmsc.2000.0723 |
| [14] | 李从先, 杨守业, 范代读, 赵娟. 三峡大坝建成后长江输沙量的减少及其对长江三角洲的影响[J]. 第四纪研究 , 2004, 24 (5) : 495–500. |
| [15] | 王保栋, 战闰, 藏家业. 长江口及其邻近海域营养盐的分布特征和输送途径[J]. 海洋学报 , 2002, 24 (1) : 53–58. |
| [16] | Chai C, Yu Z M, Shen Z L, Song X X, Cao X H, Yao Y. Nutrient characteristics in the Yangtze River Estuary and the adjacent East China Sea before and after impoundment of the Three Gorges Dam[J]. Science of the Total Environment , 2009, 407 (16) : 4687–4695. DOI:10.1016/j.scitotenv.2009.05.011 |
| [17] | 刘光兴, 陈洪举, 朱延忠, 齐衍萍. 三峡工程一期蓄水后长江口及其邻近水域浮游动物的群落结构[J]. 中国海洋大学学报 , 2007, 37 (5) : 789–794. |
| [18] | Jiang H, Cheng H Q, Xu H G, Arreguín-Sánchez F, Zetina-Rejón M J, Del Monte Luna P, William J F L. Trophic controls of jellyfish blooms and links with fisheries in the East China Sea[J]. Ecological Modelling , 2008, 212 (3/4) : 492–503. |
| [19] | Guo C B, Ye S W, Lek S, Liu J S, Zhang T L, Yuan J, Li Z J. The need for improved fishery management in a shallow macrophytic lake in the Yangtze River basin: evidence from the food web structure and ecosystem analysis[J]. Ecological Modelling , 2013, 267 : 138–147. DOI:10.1016/j.ecolmodel.2013.07.013 |
| [20] | Lin Q, Jin X S, Zhang B. Trophic interactions, ecosystem structure and function in the southern Yellow Sea[J]. Chinese Journal of Oceanology and Limnology , 2013, 31 (1) : 46–58. DOI:10.1007/s00343-013-2013-6 |
| [21] | 李睿, 韩震, 程和琴, 江红. 基于Ecopath模型的东海区生物资源能量流动规律的初步研究[J]. 资源科学 , 2010, 32 (4) : 600–605. |
| [22] | 李云凯, 宋兵, 陈勇, 禹娜, 陈立侨. 太湖生态系统发育的Ecopath with Ecosim动态模拟[J]. 中国水产科学 , 2009, 16 (2) : 257–265. |
| [23] | 农业部渔业局. 中国渔业统计年鉴 , 2000 . |
| [24] | 农业部渔业局. 中国渔业统计年鉴 , 2006 . |
| [25] | 唐启升. 海洋食物网与高营养层次营养动力学研究策略[J]. 海洋水产研究 , 1999, 20 (2) : 1–11. |
| [26] | Polovina J J. An overview of the Ecopath model[J]. Fishbyte , 1984, 2 : 5–7. |
| [27] | Ulanowicz R E. Growth and Development: Ecosystem Phenomenology , 1986 : 203. |
| [28] | Christensen V, Pauly D. Ecopath II-a software for balancing steady-state ecosystem models and calculating network characteristics[J]. Ecological Modelling , 1992, 61 (3/4) : 169–185. |
| [29] | 仝龄. Ecopath-一种生态系统能量平衡评估模式[J]. 海洋水产研究 , 1999, 20 (2) : 103–107. |
| [30] | Allen K R. Relation between production and biomass[J]. Journal of the Fisheries Research Board of Canada , 1971, 28 (10) : 1573–1581. DOI:10.1139/f71-236 |
| [31] | Palomares M L D, Pauly D. A multiple regression model for predicting the food consumption of marine fish populations[J]. Marine and Freshwater Research , 1989, 40 (3) : 259–273. DOI:10.1071/MF9890259 |
| [32] | 张波, 唐启升, 金显仕. 东海高营养层次鱼类功能群及其主要种类[J]. 中国水产科学 , 2007, 14 (6) : 939–949. |
| [33] | 沈新强, 陈求稳.重大水利工程影响下长江口环境与生态安全:国家重点基础研究发展计划(973计划)课题结题验收报告.南京. 2014. |
| [34] | 李云凯, 禹娜, 陈立侨, ChenYong, 冯德祥. 东海南部海区生态系统结构与功能的模型分析[J]. 渔业科学进展 , 2010, 31 (2) : 30–39. |
| [35] | 宋兵.太湖渔业和环境的生态系统模型研究[D].上海:华东师范大学, 2004. |
| [36] | Christensen V, Walters C J, Pauly D. Ecopath with Ecosim: A User's Guide , 2005 : 39. |
| [37] | 罗秉征, 韦晟, 窦硕增. 长江口鱼类食物网与营养结构的研究[J]. 海洋科学集刊 , 1997, 38 : 143–153. |
| [38] | 张波, 金显仕, 唐启升. 长江口及邻近海域高营养层次生物群落功能群及其变化[J]. 应用生态学报 , 2009, 20 (2) : 344–351. |
| [39] | 陈渊泉. 长江河口区渔业资源特点、渔业现状及其合理利用的研究[J]. 中国水产科学 , 1999, 6 (5) : 48–51. |
| [40] | Ulanowicz R E, Puccia C J. Mixed trophic impacts in ecosystems[J]. Coenoses , 1990, 5 : 7–16. |
| [41] | Libralato S, Christensen V, Pauly D. A method for identifying keystone species in food web models[J]. Ecological Modelling , 2006, 195 (3/4) : 153–171. |
| [42] | 宋洪军, 张学雷, 王保栋, 孙霞, 王晓, 辛明. 长江口及邻近海域浮游植物现存量的上下行控制分析[J]. 海洋学报 , 2014, 36 (8) : 91–100. |
| [43] | 王勇.中国若干典型海域作用于浮游植物上下行效应的研究[D].青岛:中国科学院, 2001. |
| [44] | 陆健健. 河口生态学 , 2003 : 198. |
| [45] | 陈作志, 邱永松, 贾晓平, 黄梓荣, 王跃中. 基于Ecopath模型的北部湾生态系统结构和功能[J]. 中国水产科学 , 2008, 15 (3) : 460–468. |
| [46] | 陈作志, 邱永松, 贾晓平. 北部湾生态通道模型的构建[J]. 应用生态学报 , 2006, 17 (6) : 1107–1111. |
| [47] | 江红, 程和琴, 徐海根, Arreguín-SánchezF, LeQuesne W. 大型水母爆发对东海生态系统中上层能量平衡的影响[J]. 海洋环境科学 , 2010, 29 (1) : 91–94. |