 2016, Vol. 36
2016, Vol. 36文章信息
- 问荣荣, 马玲, 刘哲强, 焦玥, 顾伟, 满子源, 刘雪英
- WEN Rongrong, MA Ling, LIU Zheqiang, JIAO Yue, GU Wei, MAN Ziyuan, LIU Xueying.
- 小兴安岭森林恢复期不同植被区域蛾类组成及其多样性
- Diversity and species composition of moths in different forests types at different stages of restoration on Xiaoxing'an Mountain, Northeast China
- 生态学报[J]. 2016, 36(15): 4816-4823
- Acta Ecologica Sinica[J]. 2016, 36(15): 4816-4823
- http://dx.doi.org/10.5846/stxb201501140116
-
文章历史
- 收稿日期: 2015-01-14
- 网络出版日期: 2015-11-16





森林生态系统是陆地生态系统中面积最大、最重要的自然生态系统,昆虫作为森林生态系统的一个重要组成部分,其种群数量变化和行为变动与森林生态系统密切相关,选择昆虫作为森林生态系统评价的指示性生物有着极大的应用潜力[1]。鳞翅目是昆虫纲中第二大目,是农林害虫中种类最多的一个目,蛾类约占鳞翅目昆虫的90%[2]而且食性大多为植食性,与生境中植物群落有着密切的联系,同时也是食物网中的重要环节。
小兴安岭林区是我国重点林区之一,也是中国天然林生态系统的核心区域之一。黑龙江凉水国家级自然保护区坐落于小兴安岭东南端,达里带岭支脉的东坡。区内森林类型多样,既有处于演替顶级状态的原始红松林,又有处于不同演替阶段的次生林以及红松、落叶松、云杉等树种的人工林,几乎囊括了小兴安岭山脉的所有森林类型。凉水国家级自然保护区是小兴安岭林区开发前的真实缩影,具有很高的科学研究价值。众多学者对凉水自然保护区植被以及土壤等的研究已日渐成熟,但对昆虫的研究还刚刚起步[3-6],还未曾有学者对蛾类群落结构进行调查。
本研究根据植被类型的不同,选取了小兴安岭凉水自然保护区内处于森林恢复期的不同植被区域与未受人类活动干扰的原始红松林共4个区域进行采样,调查分析各区域蛾类群落结构动态,分析不同恢复区域与原始林区域之间蛾类群落的差异。旨在为森林恢复区的保护和资源分配提供昆虫方面的基础数据,并为其环境质量监测提供依据,进而为小兴安岭地区生物多样性保护、森林生态可持续发展规划提供理论基础。
1 研究区域和研究方法 1.1 样地选取根据小兴安岭地区凉水自然保护区的原始林和不同植被类型恢复区域,选择了4个典型样地进行研究。分别为天然针阔混交林恢复区域(Ⅰ)、人工次生林恢复区域(Ⅱ)、天然阔叶林恢复区域(Ⅲ)、原始红松林区域(Ⅳ)。各植被恢复区域均为封山育林状态下恢复50a的森林类型,其中,样地(Ⅰ)为经择伐的天然林,树种组成为6云3白1冷,优势树种平均年龄130a,平均胸径28cm;样地(Ⅱ)为人工林,树种组成5云3冷2白,优势树种平均年龄58a,平均胸径23cm;样地(Ⅲ)为皆伐后天然恢复的阔叶混交林,树种组成3白2椴1红1色1云1水1枫,优势树种平均年龄55a,平均胸径29cm。原始林实验样地(Ⅳ),树种组成为7红1冷1椴1榆,优势树种平均年龄220年,平均胸径38cm。各样地土壤类型均为暗棕壤。
1.2 研究方法调查采样时间为2014年6月1日至2014年9月15日,各样地均每20d调查1次,遇不良天气顺延。调查方法为灯诱法,灯诱时间为19:30—23:30,诱集灯为450W高压汞灯,幕布采用1.5×1.5m2白色幕布。在各样地边缘随机选取样地,用支架将诱虫灯挂于幕布前15cm处,利用蛾类的趋光性原理于样地边缘诱集昆虫,并在诱集灯下放铺放白纸以收集落在地面的昆虫。将诱集到的昆虫用装有乙酸乙酯毒瓶毒杀,带回室内,分类制作标本后鉴定其种类并记录每种昆虫数量。
大部分蛾类标本(包括所有优势种)均鉴定到种,部分难于鉴定的标本鉴定到科。所有标本保存在东北林业大学凉水自然保护区昆虫标本室。
1.3 数据处理实验数据采用Excel 2010和SPSS19.0等统计分析软件处理。
多样性分析主要采用以下指数:个体总数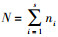
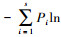
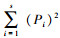
种-多度分析采用X轴为倍程刻度(log3对数级数),Y轴为普通算术刻度的坐标系,以蛾类昆虫个体数为横坐标,蛾类种类数为纵坐标,描绘种-多度曲线图。
相似性分析采用Jaccard系数I=a/(a+b+c)(a为两个样方共有种数,b为存在于样方j但不存在于样方k的种数,c为存在于样方k但不存在于样方j的种数)。群落的排序采用极点排序法,通过排序将各样点昆虫群落作为点,并且用各群落属性作为坐标轴,在二维空间中按照各昆虫群落的相似关系将其排列。
2 结果与分析 2.1 不同植被区域蛾类群落组成实验共采集到蛾类昆虫标本56079号,隶属于28科598种(表 1)。其中夜蛾科(226种)、尺蛾科(121种)为小兴安岭地区蛾类优势类群,分别占总种数的37.8%和20.2%。螟蛾科(54种)、舟蛾科(45种)、卷蛾科(40种)的种类也较多。从各类群个体数量上来看,夜蛾科(20646头)和螟蛾科(14410头)数量最多,分别占总个体数的36.8%和25.7%,其次为尺蛾科(5851头)和灯蛾科(3721头)。巢蛾科虽然只有3种,但是由于区域Ⅱ7月中旬稠李巢蛾和苹果巢蛾的大量发生,总数量在所有科中居第5位。
| 种类 Species |
Ⅰ | Ⅱ | Ⅲ | Ⅳ | 合计Total | |||||||
| S | N | S | N | S | N | S | N | S | R | N | R | |
| 波纹蛾科 Thyatiridae | 6 | 100 | 8 | 233 | 7 | 681 | 6 | 206 | 8 | 1.3 | 1220 | 2.2 |
| 蚕蛾科 Bombycidae | 1 | 12 | 1 | 32 | 1 | 28 | 1 | 18 | 1 | 0.2 | 90 | 0.2 |
| 草蛾科 Ethmiidae | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0.2 | 1 | 0.0 |
| 巢蛾科 Yponomeutidae | 3 | 147 | 3 | 2856 | 3 | 175 | 2 | 46 | 3 | 0.5 | 3224 | 5.7 |
| 尺蛾科 Geometridae | 83 | 1065 | 93 | 1512 | 85 | 1523 | 99 | 1751 | 121 | 20.2 | 5851 | 10.4 |
| 刺蛾科 Limacodidae | 3 | 12 | 4 | 39 | 5 | 39 | 3 | 27 | 5 | 0.8 | 117 | 0.2 |
| 大蚕蛾科 Saturniidae | 4 | 40 | 5 | 66 | 4 | 86 | 3 | 43 | 5 | 0.8 | 235 | 0.4 |
| 灯蛾科 Arctiidae | 14 | 1432 | 16 | 721 | 18 | 988 | 13 | 580 | 18 | 3.0 | 3721 | 6.6 |
| 毒蛾科 Lymantriidae | 15 | 621 | 15 | 428 | 14 | 680 | 15 | 294 | 18 | 3.0 | 2023 | 3.6 |
| 钩蛾科 Drepanidae | 4 | 323 | 4 | 206 | 5 | 176 | 5 | 70 | 5 | 0.8 | 775 | 1.4 |
| 冠翅蛾科 Ypsolophidae | 0 | 0 | 0 | 0 | 2 | 4 | 1 | 3 | 2 | 0.3 | 7 | 0.0 |
| 虎蛾科 Agaristidae | 0 | 0 | 1 | 8 | 1 | 6 | 1 | 1 | 1 | 0.2 | 15 | 0.0 |
| 举肢蛾科 Stathmopodidae | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 0.2 | 1 | 0.0 |
| 卷蛾科 Tortricidae | 20 | 176 | 28 | 303 | 18 | 180 | 26 | 380 | 40 | 6.7 | 1039 | 1.9 |
| 枯叶蛾科 Lasiocampidae | 9 | 62 | 8 | 67 | 8 | 99 | 9 | 47 | 10 | 1.7 | 275 | 0.5 |
| 瘤蛾科 Nolidae | 1 | 1 | 2 | 3 | 2 | 9 | 3 | 20 | 4 | 0.7 | 33 | 0.1 |
| 箩纹蛾科 Brahmaeidae | 1 | 4 | 1 | 7 | 1 | 15 | 1 | 2 | 1 | 0.2 | 28 | 0.0 |
| 麦蛾科 Gelechiidae | 1 | 1 | 1 | 11 | 0 | 0 | 2 | 7 | 3 | 0.5 | 19 | 0.0 |
| 螟蛾科 Pyralidae | 32 | 4888 | 40 | 4049 | 38 | 4105 | 34 | 1368 | 54 | 9.0 | 14410 | 25.7 |
| 木蠹蛾科 Cossidae | 0 | 0 | 1 | 1 | 2 | 7 | 1 | 1 | 2 | 0.3 | 9 | 0.0 |
| 鞘蛾科 Coleophoridae | 1 | 13 | 1 | 25 | 1 | 5 | 1 | 16 | 1 | 0.2 | 59 | 0.1 |
| 蓑蛾科 Psychidae | 1 | 2 | 0 | 0 | 1 | 2 | 0 | 0 | 1 | 0.2 | 4 | 0.0 |
| 天蛾科 Sphingidae | 16 | 106 | 18 | 114 | 16 | 208 | 12 | 103 | 19 | 3.2 | 531 | 0.9 |
| 网蛾科 Thyrididae | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0.2 | 1 | 0.0 |
| 夜蛾科 Noctuidae | 165 | 5929 | 165 | 6080 | 170 | 4870 | 124 | 3767 | 226 | 37.8 | 20646 | 36.8 |
| 羽蛾科 Pterophoridae | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 0.2 | 4 | 0.0 |
| 长角蛾科 Adelidae | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0.2 | 1 | 0.0 |
| 舟蛾科 Notodontidae | 32 | 473 | 34 | 560 | 35 | 543 | 27 | 164 | 45 | 7.5 | 1740 | 3.1 |
| 合计 Total | 412 | 15047 | 451 | 17323 | 438 | 14430 | 393 | 8919 | 598 | 100 | 56079 | 100 |
| S:种数Species;N:个体数Individuals;R:相对多度Relative abundance | ||||||||||||
从科级阶元来看,各区域中蛾类科级分布差异不显著。区域Ⅰ有20个科,区域Ⅱ和区域Ⅲ均有23个科。区域Ⅳ有25个科,在所有区域中居第一位,且草蛾科、网蛾科、长角蛾科等科只在区域Ⅳ中出现,他们可能是该生境的专属科。综合不同植被区域蛾类的物种数和个体数来看,各区域均以夜蛾科为主,尺蛾科次之,各区域中各科分布差异不显著。
在区域Ⅰ中,各优势科的相对多度排序为夜蛾科(39.40)>螟蛾科(32.48)>灯蛾科(9.52)>尺蛾科(7.08);区域Ⅱ中为夜蛾科(35.09)>螟蛾科(23.37)>巢蛾科(16.49)>尺蛾科(8.73);区域Ⅲ中为夜蛾科(33.75)>螟蛾科(28.45)>尺蛾科(10.55)>灯蛾科(6.85);区域Ⅳ中为夜蛾科(42.25)>尺蛾科(19.63)>螟蛾科(15.34)>灯蛾科(6.50)。夜蛾科在各个区域中所占比例均最高,且所占比例相近,受植被类型影响较小。灯蛾科、毒蛾科和舟蛾科等科在不同区域中相对多度分布也比较均匀。尺蛾科在原始林区域所占比例最高,相对多度比其他林型高约10%。螟蛾科在3个植被恢复区域(区域Ⅰ—Ⅲ)中所占比例均位居第2位,在原始林中个体数量所占比例明显小于其他区域;区域Ⅱ中巢蛾科个体数所占比例为16.49%,明显高于其他区域。螟蛾科和巢蛾科的在各区域中个体数量分布受优势种个体数量的影响比较大,原始林区域相比各恢复区域植被类型丰富,昆虫群落内部网更为复杂,能够有效抑制蛾类优势种的爆发,控制其个体数量。
2.2 不同植被区域蛾类群落的优势种类实验调查发现,豆卷叶野螟、头橙荷苔蛾、一色兜夜蛾、阴亥夜蛾、稠李巢蛾个体数均大于2000头,高于其他种,为小兴安岭地区优势种。但不同区域优势种略有差异,各优势种优势度见表 2。豆卷叶野螟在各森林恢复期实验区域Ⅰ、Ⅱ、Ⅲ中优势度指数均为最高,而原始林实验区域Ⅳ中优势度指数最高的是阴亥夜蛾。稠李巢蛾仅在区域Ⅱ中为优势种,且优势度指数较高,在其他区域数量较少。区域Ⅳ相对区域Ⅰ、Ⅱ、Ⅲ优势种较少,且各优势种优势度指数之间相对更接近。
| 种名 Species | 优势度指数 Dominance index | ||||
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | 合计 Total | |
| 豆卷叶野螟 Pleuroptya ruralis (Scopoli) | 0.2441 | 0.1460 | 0.2023 | 0.0586 | 0.1740 |
| 头橙荷苔蛾Ghoria gigantea (Oberthür) | 0.0732 | 0.0266 | 0.0368 | 0.0420 | 0.0447 |
| 阴亥夜蛾 Hydrillodes funeralis (Warren) | 0.0452 | 0.0378 | 0.0153 | 0.0974 | 0.0435 |
| 一色兜夜蛾Cosmia unicolora (Staudinger) | 0.0419 | 0.0636 | 0.0090 | 0.0697 | 0.0445 |
| 稠李巢蛾Yponomeuta evonymellus (Linnaeus) | 0.0049 | 0.0876 | 0.0064 | 0.0028 | 0.0303 |
| 苹果巢蛾 Yponomeuta padella (Linnaeus) | 0.0041 | 0.0721 | 0.0053 | 0.0022 | 0.0250 |
| 仑狭翅夜蛾Hermonassa arenosa (Butler) | 0.0152 | 0.0315 | 0.0548 | 0.0122 | 0.0298 |
在群落调查中,往往得到许多个体数很少的罕见种和几种个体数很多的富集种,而种-多度关系可以反映群落中各物种分享可利用资源的方式,比较各物种的相对重要性[7]。对蛾类种-多度关系处理,经拟合[8]分析,在随机生态位假说、生态位优先占领假说和对数正态分布假说3个模型中,4个区域均更接近于对数正态分布假说,在这样的群落里,十分富集的物种和很稀有的物种都较少,而处于中间状态的物种较多。
按Preston[9]方法分别对4区域蛾类多度进行对数正态分布模型拟合[10],结果见图 1。经拟合得出4区域种-多度曲线公式为:Ⅰ,S(R)=105e-[0.36(R-1)]2,R2=0.9958;Ⅱ,S(R)=117e-[0.44(R-2)]2,R2=0.9958;Ⅲ,S(R)=100e-[0.38(R-2)]2,R2=0.9240;Ⅳ,S(R)=119e-[0.51(R-2)]2,R2=0.9421,其中S(R)为第R倍程的理论种数,R为倍程数,e为是自然对数函数的底,R2为拟合优度。

|
| 图 1 不同植被区域蛾类群落种-多度曲线 Fig. 1 The curve of species abundance of the moths in different areas in Liangshui Reserve |
按公式ST=S0SQRT(π)/α[11]估计小兴安岭蛾类总种类数为687种。
2.4 不同植被区域蛾类群落多样性指数分析从表 3可以看出,各森林恢复区域蛾类个体总数均明显高于原始林区域,人工次生林Ⅱ个体总数最多,其次为天然针阔混交林Ⅰ和天然阔叶林Ⅲ,这可能与原始林区域在长期的自然演替的过程中形成了稳定的昆虫群落,群落中天敌昆虫和植食性昆虫比例适当,各昆虫之间相互制约,能够很好的抑制优势种的爆发,因此也制约了蛾类优势种的个体数量。
| 区域 Areas |
个体总数 Individual number |
多样性指数 Diversity index |
均匀度指数 Evenness index |
优势集中性指数 Dominant concentration index |
| Ⅰ | 15407 | 3.9191 | 0.6506 | 0.0780 |
| Ⅱ | 17323 | 4.1386 | 0.6769 | 0.0497 |
| Ⅲ | 14430 | 4.3966 | 0.7229 | 0.0520 |
| Ⅳ | 8919 | 4.6177 | 0.7730 | 0.0264 |
各区域多样性指数均较高,多样性指数和均匀度指数大小次序为原始红松林Ⅳ>天然阔叶林Ⅲ>人工次生林Ⅱ>天然针阔混交林Ⅰ;优势集中性指数与多样性指数大小次序大致相反,天然针阔混交林Ⅰ>天然阔叶林Ⅲ>人工次生林Ⅱ>原始红松林Ⅳ。总体上说原始红松林Ⅳ中蛾类多样性水平最高,并且蛾类各种类之间数量分布相对最为均衡;天然针阔混交林Ⅰ蛾类优势种明显,其多样性在几个区域中相对较差。植物群落对昆虫的多样性有着重要的影响,蛾类昆虫多数是植食性的,显示了与植物更高的相关性[12]。几个植被区域中,原始林区域在长期的自然演替过程中形成了更加稳定多样的植被环境,天然阔叶林区域林分类型多样有白桦、椴树、红松、云杉等多种主要组成树种,因此两个林型多样性各指数位居前两位。
昆虫群落均匀度、优势集中性和个体数都是决定多样性的重要参数,本实验中多样性指数与均匀度指数不一致(r=0.173—0.471,P>0.05),与优势集中性指数显著负相关(r=-0.7—-0.974,P<0.05)或相关性不显著(原始林区域Ⅳ,r=-0.759,P>0.05),与物种数不一致(r=0.397—0.669,P>0.05)或一致(原始林区域Ⅳ,r=0.821,P<0.05)。
2.5 不同植被区域蛾类群落的相似性分析各区域间蛾类群落的相似性系数见表 4,从表中可以看出,各区域相似性系数分布在0.5470—0.6764之间,SI均大于0.5。根据Jaccard[13]的相似性系数原理,相似系数SI的大小反映两地区间物种相似程度,各区域之间的相似性水平较高,处于中等相似水平。由此可见各区域蛾类类群相近,这主要是因为凉水自然保护区多年的封山育林,使各恢复区域都得到了良好的保护,环境条件良好,均接近未受人为干扰的原始林红松林区域。
| 区域 Areas | Ⅰ | Ⅱ | Ⅲ | Ⅳ |
| Ⅰ | - | 0.6764 | 0.6621 | 0.5470 |
| Ⅱ | - | 0.6667 | 0.6034 | |
| Ⅲ | - | 0.5739 | ||
| Ⅳ | - |
采用极点排序法计算得到各群落的相关数值,对各区域蛾类群落进行排序。从图 2的排序结果可以看出,3个恢复区域之间蛾类群落较为靠近,而与原始红松林区域距离较大。其中,天然阔叶林恢复区域与原始红松林区域之间蛾类群落差异最大,人工次生林恢复区域与原始林区域差异最小。

|
| 图 2 不同植被区域蛾类群落极点排序 Fig. 2 Polar ordination of month communities |
通过灯诱法对凉水自然保护区蛾类系统调查发现,小兴安岭地区蛾类种类繁多,数量丰富。凉水自然保护区已知蛾类28科598种,其中夜蛾科、尺蛾科为小兴安岭地区蛾类优势类群,其次为螟蛾科、舟蛾科、卷蛾科。豆卷叶野螟、头橙荷苔蛾、一色兜夜蛾、阴亥夜蛾、稠李巢蛾等为小兴安岭地区优势种。原始林区域优势度最高的种与区域Ⅰ、Ⅱ、Ⅲ不同,且与其他区域相比各优势种优势度指数之间相对更为接近,无某种蛾类大量发生。这与原始林中自然环境良好,植被类型丰富,天敌昆虫与植食性昆虫比例均衡密切相关,而森林生态系统组成多样性和结构复杂性是实现森林生态系统对病虫害自我调控功能的基础条件[14-15]。小兴安岭地区和长白山地区森林植被相近,均具有阔叶红松林的原始林区域和处于不同恢复期的次生林和人工林区域,而森林植被类型是决定蛾类物种和数量的主要因素[16],转化性高的植食性昆虫的多样性受生境内植物组成的影响[17]。小兴安岭地区蛾类优势类群与众多学者[18-21]对长白山阔叶红松林蛾类组成研究结果相近,在长白山采集到的蛾类27科中,除织蛾科外,在本实验中均有获得,并且均以夜蛾科、尺蛾科、螟蛾科、舟蛾科、灯蛾科等为优势科。长白山地区蛾类优势种,如尘尺蛾 Hypomecis punctinalis conferenda (Butler)、枞灰尺蛾 Deileptenia ribeata (Clerck)、短扇舟蛾 Clostera albosigma curtuloides Erschoff、阿泊波纹蛾 Tethea ampliata (Butler)、灰歹夜蛾 Diarsia canescens (Butler)、黄绿组夜蛾 Anaplectoides virens (Butler) 等在本实验中均有获得,且个体数目均大于100。但长白山地区以尺蛾科个体数和物种数量最为丰富,而本调查发现小兴安岭地区蛾类以夜蛾科最为占优;两地蛾类昆虫的优势种不同,本调查中所采集到的优势种中仅有仑狭翅夜蛾 Hermonassa arenosa (Butler)在长白山蛾类优势种报道中有提及。虽然小兴安岭地区和长白山地区的主要森林类型相近,但是两地森林主要树种组成比例以及林下植被的不同和各蛾类类群对两地环境条件的长期适应造成了两地蛾类组成有一定的差异。
对四个区域进行种-多度分析均表明,小兴安岭地区蛾类种-多度关系比较接近对数正态分布假说,符合这种分布的群落多属于环境条件较好、物种丰富而分布又比较均匀的群落[7]。尤平等[22]对天津湿地蛾类种-多度进行分析,发现七里海和鸭淀符合对数正态分布假说,环境质量好;而团泊洼、北大港和独流减河接近接生态位优先占领假说,生态环境较差。杨向东等[23]研究冀西北坝上地区蛾类接近与对数正态分布,也表明该地蛾类群落环境良好。四个区域昆虫群落对数正态分布模型拟合优度均较高,这说明凉水自然保护区经过多年的封山育林和合理的保护措施,原始林区域和森林恢复区域均有着优越的环境条件,适合多种物种共存,昆虫群落结构稳定。
对四个区域多样性分析发现,各森林恢复区域蛾类个体总数均明显高于原始林区域,这可能是由于原始林区域植被类型丰富,昆虫群落内部网络关系复杂,可以有效抑制蛾类优势种的大量发生。原始林区域多样性指数相对较高,优势集中性指数相对较低同样支持这一论述。天然针阔混交林区域多样性、均匀度较其他区域均较差,而且优势种更加明显,因此需要加强对其环境的保护和蛾类优势种的监测,防止虫害的发生。原始林区域Ⅳ物种多样性与蛾类物种数和个体数均保持一致,但各植被恢复区域相关性不明显,因为多样性的高低除了要考虑物种数外,还要注意均匀度的影响[24]。均匀度对多样性的影响,不同学者研究结构不一致。贺达汉等[25]认为荒漠草原昆虫群落均匀度和多样性指数一致,说明群落结构稳定;侯沁文[26]等研究历山保护区蛾类多样性与均匀度不一致,但环境条件优越。本研究中,蛾类多样性与均匀度指数不一致,但与Simpson优势度集中性指数高度一致,说明优势种的数量对群落多样性指数也有一定的影响。
昆虫群落的极点排序结果可以直观地反映区域间环境条件关系[27]。从不同区域蛾类群落的极点排序,可以直观地看出原始红松林和其他区域距离较大,说明小兴安岭地区各植被恢复区域与原始林均存在着环境差异,但各区域之间的相似性系数均大于0.5,各区域之间蛾类群落结构差异不大,群落稳定性较高。天然阔叶林恢复区域与原始红松林区域之间距离最大,说明二者之间蛾类群落差异最大,这可能是因为原始红松林林分类型以针叶林为主,蛾类寄主植物与由阔叶树组成的天然阔叶林区域差异较大。尤平等[22]研究也发现蛾类群落和植被类型有一定的相关性。
综合以上结果分析,种类众多、数量丰富的昆虫是森林生态系统的中物种多样性的重要组成部分,把蛾类可以作为指示物种来监测环境变化是可行的。总体来说,凉水自然保护区,经过多年合理的保护措施,各区域昆虫多样性水平较高,昆虫群落结构稳定,有着优越的环境条件,多年的森林恢复措施效果显著。但各森林恢复区域的昆虫群落结构和自然环境与原始林之间依然存在着差异,森林生态系统受到破坏之后再恢复需要很长的时期。与原始林相比,各恢复区域蛾类优势种类多且数量偏高,因此需要对豆卷叶野螟、一色兜夜蛾等蛾类寄主植物进行监控,防止虫害发生。天然针阔混交林区域和人工次生林区域相对与其他区域,植被组成略显单一,蛾类优势种更为明显,多样性较低,通过采取合理措施,调整林分结构、丰富林下植被,增加天敌昆虫调节昆虫间捕食和竞争关系,进而控制蛾类优势种数量。
| [1] | Akutsu K, Khen C V, Toda M J. Assessment of higher insect taxa as bioindicators for different logging-disturbance regimes in lowland tropical rain forest in Sabah, Malaysia[J]. Ecological Research , 2007, 22 (4) : 542–550. DOI:10.1007/s11284-006-0052-6 |
| [2] | 郑乐怡, 归鸿. 昆虫分类. 南京: 南京师范大学出版社, 1999805–806. |
| [3] | 顾伟, 马玲, 刘哲强, 焦玥, 王利东, 张琛. 凉水自然保护区不同林型地表甲虫物种多样性[J]. 东北林业大学学报 , 2014 (1) : 131–135. |
| [4] | 殷秀琴, 吴东辉, 韩晓梅. 小兴安岭森林土壤动物群落多样性的研究[J]. 地理科学 , 2003, 23 (3) : 316–322. |
| [5] | 安静超, 殷秀琴. 小兴安岭凉水自然保护区土壤昆虫群落特征及多样性研究[J]. 东北师大学报: 自然科学版 , 2010, 42 (3) : 139–144. |
| [6] | 刘哲强, 马玲, 焦玥, 张静, 曹传旺, 顾伟, 满子源, 张曼胤. 小兴安岭森林恢复期典型人工林与原始林昆虫群落结构动态[J]. 应用生态学报 , 2015, 26 (2) : 555–562. |
| [7] | 赵志模, 郭依泉. 群落生态学原理与方法. 重庆: 科学技术文献出版社重庆分社, 1990. |
| [8] | 刘梦雪, 刘佳佳, 杜晓光, 郑小刚. 亚高寒草甸不同生境植物群落物种多度分布格局的拟合[J]. 生态学报 , 2010, 30 (24) : 6935–6942. |
| [9] | Preston F W. The commonnes, and rarity, of species[J]. Ecology , 1948, 29 : 254–283. DOI:10.2307/1930989 |
| [10] | Guo X G, Wu D, Meng X Y, Dong W G, Qian T J. Species abundance distribution of ectoparasitic gamasid mites on Rattus flavipectus[J]. Journal of Pathogen Biology , 2006, 1 (1) : 15–19. |
| [11] | May R M. Patterns of species abundance and diversity//Cody M L, Diamond J M, eds. Ecology and Evolution of Communities. Cambridge: Harvard University Press, 1975: 81-120. |
| [12] | 朱慧, 彭媛媛, 王德利. 植物对昆虫多样性的影响[J]. 生态学杂志 , 2008, 27 (12) : 2215–2221. |
| [13] | Jaccard P. Distribution de la flore alpine: dans le bassin des dranses et dans quelques régions voisines[J]. Bulletin de la Société Vaudoise des Sciences Naturelles , 1901, 37 (140) : 241–272. |
| [14] | 曾凡勇, 孙志强. 森林生态系统中植食性昆虫与寄主的互作机制、假说与证据[J]. 生态学报 , 2014, 34 (5) : 1061–1071. |
| [15] | 梁军, 朱彦鹏, 孙志强, 张星耀. 森林生态系统组成和结构与病虫害防治[J]. 中国森林病虫 , 2012, 31 (5) : 7–12. |
| [16] | Usher M B, Keiller S W J. The macrolepidoptera of farm woodlands: determinants of diversity and community structure[J]. Biodiversity & Conservation , 1998, 7 (6) : 725–748. |
| [17] | Lepš J, Novotný V, Basset Y. Habitat and successional status of plants in relation to the communities of their leaf-chewing herbivores in Papua New Guinea[J]. Journal of Ecology , 2001, 89 (2) : 186–199. DOI:10.1046/j.1365-2745.2001.00540.x |
| [18] | 贾玉珍, 张春雨, 赵秀海, 孟庆繁, 夏富才. 长白山红松阔叶林不同恢复阶段蛾类组成和多样性研究[J]. 应用与环境生物学报 , 2008, 14 (5) : 630–634. |
| [19] | 孟庆繁, 高文韬, 刘生冬. 长白山北坡针叶林带蛾类多样性[J]. 东北林业大学学报 , 2008, 36 (3) : 60–62. |
| [20] | 王珍, 姬兰柱, 张悦, 易雪梅. 长白山三种林型对蛾类群落结构和多样性的影响[J]. 生态学杂志 , 2012, 31 (5) : 1214–1220. |
| [21] | 刘生冬, 孟庆繁, 高文韬. 长白山自然保护区北坡蛾类的多样性[J]. 东北林业大学学报 , 2007, 35 (10) : 51–53. |
| [22] | 尤平, 李后魂. 天津湿地蛾类丰富度和多样性及其环境评价[J]. 生态学报 , 2006, 26 (3) : 629–637. |
| [23] | 杨向东, 董建臻, 李瑞军, 张立峰, 黄亚群. 冀西北坝上地区灯下蛾类群落结构特征[J]. 生态学报 , 2010, 30 (15) : 4234–4240. |
| [24] | 马玲, 顾伟, 丁新华, 骆有庆, 韩争伟, 吴思亮. 扎龙湿地昆虫群落结构及动态[J]. 生态学报 , 2011, 31 (5) : 1371–1377. |
| [25] | 贺答汉, 田畴, 任国栋, 郝峰茂, 马世瑜. 荒漠草原昆虫的群落结构及其演替规律初探[J]. 中国草地 , 1988, 11 (6) : 24–28. |
| [26] | 侯沁文, 白海艳, 铁军, 边高鹏, 史宝忠. 山西历山国家级自然保护区不同演化阶段林型蛾类多样性[J]. 应用昆虫学报 , 2014, 51 (5) : 1310–1319. |
| [27] | 韩争伟, 马玲, 曹传旺, 张静, 王步勇. 太湖湿地昆虫群落结构及多样性[J]. 生态学报 , 2013, 33 (14) : 4387–4397. |