 2016, Vol. 36
2016, Vol. 36文章信息
- 吴文广, 张继红, 王巍, 李加琦, 房景辉, 刘毅, 高振锟, 张义涛, 陈洁
- WU Wenguang, ZHANG Jihong, WANG Wei, LI Jiaqi, FANG Jinghui, LIU Yi, GAO Zhenkun, ZHANG Yitao, CHEN Jie.
- 桑沟湾春季叶绿素a浓度分布及其影响因素
- Distribution of chlorophyll-a concentration and its control factors in spring in Sungo Bay
- 生态学报[J]. 2016, 36(15): 4855-4863
- Acta Ecologica Sinica[J]. 2016, 36(15): 4855-4863
- http://dx.doi.org/10.5846/stxb201501130102
-
文章历史
- 收稿日期: 2015-01-13
- 网络出版日期: 2015-11-16

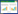




浮游植物作为海洋初级生产力的主要贡献者,是海洋生态系统的能量流动和物质流动的关键环节,它是海水养殖活动的重要支撑,是滤食性贝类的主要食物之一[1-2]。叶绿素a浓度是表征海洋中浮游植物现存生物量和光合作用有机碳同化能力的重要指标,是海域肥瘠程度和评价海域生态环境的重要依据[3-5]。有关养殖海域叶绿素a浓度分布及其变化的研究已有报道[6-9],叶绿素a浓度的高低在一定程度上能反映海区海水质量状况,同时也是评价水体理化性质动态变化的综合反映指标之一。桑沟湾是中国最为著名的天然养殖良港之一,海产品产量高、品质佳,其环境特点、养殖品种、养殖规模和养殖模式等均具有代表性。养殖的主要种类包括栉孔扇贝、长牡蛎、虾夷扇贝、海带、海参、龙须菜等,养殖方式以筏式养殖为主,网箱养殖和底播养殖为辅。因此该海域是研究养殖活动与叶绿素a相互关系的理想海域。自20世纪80年代以来,已有专家学者对桑沟湾的浮游植物、营养盐和海水pCO2[10-14]等进行了深入的研究,对桑沟湾主要养殖区表底层叶绿素a及其定点连续监测的研究报道较少。
叶绿素a浓度的分布变化受水温、养殖活动、营养盐、昼夜节律和潮汐等因素的影响,海水营养盐是海洋浮游植物生长和繁殖的关键因子,是海洋初级生产力和食物链的基础。N、P、Si是海洋浮游植物必不可少的营养盐,由于不同海区的状况不同,以上几项营养盐都有可能成为浮游植物生长的限制因子[15]。在海水营养盐充足的情况下,叶绿素a浓度随温度升高而增大,温度是叶绿素a浓度增长的限制因子[2]。不同的海水养殖模式、养殖品种和养殖水域等均能引起叶绿素a浓度的改变,但不同海域的叶绿素a浓度的变化及其影响因素不尽相同,尤其养殖海域的特定地理位置及独特的养殖模式,均会对叶绿素a浓度产生影响。桑沟湾是山东省海水养殖业的重点海湾,近年来逐渐形成了由湾内向湾外依次排列的贝类养殖区、海带和贝类混养区、海带养殖区的多元养殖模式,其多元的养殖模式就必然会对该海域叶绿素a的分布产生影响。研究特定养殖海域叶绿素a的分布及其影响因素有助于掌握不同养殖模式、养殖品种和养殖水域等环境下浮游植物生物量和水质状况,从而为指导生产提供数据支持。
2014年5月在桑沟湾主要养殖区进行表底层海水叶绿素a浓度的调查,并对其4个不同养殖区进行24h定点连续观测,分析研究叶绿素a浓度垂直分布及昼夜变化。并探讨叶绿素a浓度与营养盐(待发表数据)的关系,有助于掌握该海域不同养殖区对叶绿素a的分布特征及其变化的影响机制,为创建新型健康养殖模式及养殖容量评估提供科学依据。
1 材料与方法 1.1 调查站位本次调查于2014年5月在桑沟湾主要养殖区设置两条航线,4个连续监测站点(图 1)。两条调查船按照两条航线同时进行调查,共计14个站位。两条航线的航行线路分别为:航线1,18-15-10-6-3-1-21;航线2,19-14-11-5-4-22。航线调查时间为9:00—11:00。4个24h连续监测站点分别为:A贝类区,B海带区,C网箱区,D海草区(图 1)。桑沟湾三面环陆,湾口向东,水深0—18 m,湾内平均水深7—8 m,为典型的浅水半封闭海湾,各站位间的水深不一,底层深度变化幅度较大。

|
| 图 1 桑沟湾春季主要养殖区叶绿素a调查站位设置 Fig. 1 The Survey stations for chlorophyll-a in Sanggou Bay in spring |
样品采集参照国家《海洋监测规范》(GB17378.7—2007)进行。每个站位分别采集表层、底层水样,昼夜连续站水样采集白天每2h取样1次,晚间每3h取样1次。所有水样经0.45μm玻璃纤维滤膜过滤后滴加2滴饱和碳酸镁溶液,过滤体积为500mL,每个水样3个平行,滤膜于-20℃冷冻保存并带回实验室分析。样品的测定采用分光光度法[6]。
1.3 数据处理实验数据采用Microsoft Excel 2010和SPSS17.0统计分析软件进行统计学分析,ANOVA单因子方差分析检验组间差异,Pearson相关分析检验叶绿素a与营养盐归回关系显著性。
2 结果 2.1 航线调查桑沟湾航线1表、底层叶绿素a浓度如图 2所示。从图 2中可以看出航线1叶绿素a浓度范围在0.14—1.40μg/L,均值为0.68μg/L,除1#站位叶绿素a浓度底层高于表层外,其余站位表层叶绿素a浓度均高于底层,即外海区底层叶绿素a高于表层,贝类区、混养区和藻类区表层叶绿素a高于底层(P < 0.05)。航线1叶绿素a浓度分布趋势为:贝类区>混养区>海带区>外海区(P < 0.05)。

|
| 图 2 桑沟湾春季航线1表、底层叶绿素a浓度 Fig. 2 The chlorophyll-a concentration of surface and bottom layer for lane 1 in Sanggou Bay in spring |
桑沟湾航线2表、底层叶绿素a浓度如图 3所示。从图 3中可以看出航线2叶绿素a浓度范围在0.11—0.92μg/L,均值为0.73μg/L,表层叶绿素a浓度均高于底层,即贝类区、混养区及藻类区表层叶绿素a高于底层,外海区则相反(P < 0.05)。航线2叶绿素a浓度分布为贝类区>混养区和外海区>海带区(P < 0.05)。

|
| 图 3 桑沟湾春季航线2叶绿素a表、底层分布特征 Fig. 3 The chlorophyll-a concentration of surface and bottom layer for lane 2 in Sanggou Bay in spring |
桑沟湾主要养殖区叶绿素a浓度24h定点连续监测见图 4。从图中可以看出,网箱区叶绿素a浓度变化范围在0.92—2.65μg/L,除18:00外,网箱区叶绿素a浓度底层均高于表层(P < 0.05);海草区叶绿素a浓度变化范围在0.14—0.95μg/L,9:00—20:00即白天海草区叶绿素a浓度表层高于底层,23:00—5:00即夜间时则底层高于表层,7:00—9:00则变为表层高于底层(P < 0.05),其中海草区在23:00—5:00时间段,底层高于表层。网箱区叶绿素a浓度总体上高于海草区。

|
| 图 4 桑沟湾春季主要养殖区叶绿素a浓度24h定点连续监测 Fig. 4 The continuous observation of chlorophyll-a for 24 h from the four culture zones in Sanggou Bay in spring |
贝类区叶绿素a浓度变化范围在0.12—1.84μg/L,表层浓度均高于底层,且白天高于夜间(P < 0.05),其中00:00—04:00时间段,底层高于表层;海带区叶绿素a浓度变化范围在0.24—1.40μg/L,海带区叶绿素a浓度上午表层高于底层,下午底层高于表层,夜间先是表层高于底层,然后底层高于表层(P < 0.05),表底层叶绿素a浓度呈现出升降交替的现象。
桑沟湾春季不同养殖区连续站表底层叶绿素a浓度均值比较见表 1。从表 1中可以看出,桑沟湾4个连续站叶绿素a浓度趋势为:网箱区>贝类区>海带区和海草区(P < 0.05),除网箱区外,叶绿素a浓度均为表层略高于底层。
| 连续监测区 The continuous observation zones |
海草区 Sea grasses zone |
贝类区 Shellfish zone |
海带区 Kelp zone |
网箱区 Cage zone |
| 表层Surface layer | 0.60±0.25c | 1.31±0.45b | 0.70±0.18c | 1.53±0.48a |
| 底层Bottom layer | 0.55±0.29c | 1.16±0.32b | 0.68±0.26c | 1.87±0.36a |
| 表中不同处理之间带有不同字母的数据表示相互之间差异显著(P < 0.05) | ||||
桑沟湾夏季叶绿素a浓度与营养盐、温度以及盐度的关系显示出夏季主要养殖区叶绿素a浓度与硅酸盐、温度呈较显著的正相关关系(图 5),与盐度、氨氮、硝酸盐、亚硝酸等无明显相关性。航线表层叶绿素a浓度(C1)分别与硅酸盐浓度(S1)、温度(T)均呈较显著的正相关关系(图 5,航线表层),网箱区底层叶绿素a浓度(C2)与硅酸盐浓度(S2)呈较显著的正相关关系(图 5,网箱区底层),海带区表层叶绿素a浓度(C3)与硅酸盐浓度(S3)呈较显著的正相关关系(图 5,海带区表层)。表 2为桑沟湾春季不同养殖区营养盐平均浓度及氮磷、硅氮比数据(数据来源:张义涛, 陈洁待发表数据),从表 2中看出航线调查及四个海区氮磷比(N/P)范围在15—57.33,硅氮比(Si/N)范围在0.11—0.32。

|
| 图 5 桑沟湾春季连续监测区叶绿素a浓度与硅酸盐、温度的关系 Fig. 5 The relationship between chlorophyll-a concentration and silicate, temperature from the continuous observation zones in Sanggou Bay in spring |
| 调查站位 Survey sites |
总氮 Total nitrogen/ (μmol/L) |
溶解无机氮 DIN/ (μmol/L) |
磷酸盐 PO43--P/ (μmol/L) |
硅酸盐 SiO32--Si/ (μmol/L) |
氮磷比 Ratio N/P |
硅氮比 Ratio Si/N |
|
| 航线Lane | S | 278.47±148.21ab | 12.84±1.12a | 0.41±0.12a | 1.66±0.15b | 31.32 | 0.13 |
| B | 242.03±114.99ab | 11.80±1.03a | 0.50±0.14a | 1.29±0.11b | 23.60 | 0.11 | |
| 网箱区 | S | 119.79±42.25b | 6.41±1.25c | 0.18±0.14b | 1.08±0.48c | 35.61 | 0.17 |
| Cage zone | B | 148.40±47.52b | 5.34±0.87c | 0.18±0.1b | 0.93±0.41c | 29.67 | 0.17 |
| 海草区 | S | 335.47±79.34a | 8.60±1.64b | 0.15±0.13b | 1.35±0.27b | 57.33 | 0.16 |
| Sea grasses zone | B | 341.05±97.93a | 8.23±1.41b | 0.19±0.13b | 1.34±0.18b | 43.32 | 0.16 |
| 贝类区 | S | 119.54±75.24b | 8.33±2.13b | 0.47±0.23a | 2.70±0.69a | 17.72 | 0.32 |
| Shellfish zone | B | 142.67±84.13b | 7.64±2.22bc | 0.40±0.18a | 2.43±0.55a | 19.10 | 0.32 |
| 海带区 | S | 26.71±11.18c | 7.35±2.08bc | 0.49±0.17a | 1.69±0.40b | 15.00 | 0.23 |
| Kelp zone | B | 33.28±18.14c | 7.96±2.16b | 0.44±0.23a | 1.79±0.45b | 18.09 | 0.22 |
| DIN:溶解无机氮Dissolved inorganic nitrogen,S:为表层Surface layer,B:为底层Bottom layer; 表中不同处理之间带有不同字母的数据表示相互之间差异显著(P < 0.05) | |||||||
桑沟湾春季主要养殖区表、底层叶绿素a浓度与潮汐作用的关系见图 6,从图 6中可以看出海草区表层叶绿素a浓度低潮时大于高潮(P < 0.05),而其他3个连续监测区叶绿素a浓度变化不明显;海带区底层叶绿素a浓度高潮时大于低潮(P < 0.05),而其他3个连续监测区叶绿素a浓度变化不明显。

|
| 图 6 桑沟湾春季主要养殖区表、底层叶绿素a浓度与潮汐作用的关系 Fig. 6 The relationship between chlorophyll-a concentration and tidal effect in the continuous observation zones in Sanggou Bay in spring |
桑沟湾春季叶绿素a浓度的整体趋势是从湾内向湾外逐渐递减的,与郝林华等研究结果相吻合[2]。桑沟湾春季牡蛎、扇贝等高密度的养殖,其摄食压力并未明显降低该区域叶绿素a的浓度,而混养区和海带区的海带处于高速生长阶段,吸收大量的营养盐,限制了该区域浮游植物对营养盐的吸收,使该区叶绿素a浓度降低,这说明桑沟湾春季营养盐的限制作用较贝类的摄食压力强,即,上行控制>下行控制。此外,在大面走航时,早出晚归,从而造成取样时间上的差异,可能会给监测带来误差,误差范围因不同养殖区而异,所以在对桑沟湾未来监测时需要加强长期、连续数据的获取。
本次桑沟湾航线调查结果表明叶绿素a浓度平均值为0.70μg/L,低于历史同期叶绿素a浓度均值(表 3),并且与其他养殖海湾相比,桑沟湾海域春季叶绿素a浓度均值也处于较低水平,但是,桑沟湾春季表层叶绿素a浓度 < 1 ug/L,浮游植物已经成为滤食性贝类生长的限制因子[15]。近年来专家学者对桑沟湾养殖品种结构、养殖布局规划不断改善和优化,明显改善了桑沟湾的养殖环境,从而导致桑沟湾春季叶绿素a浓度低于历史同期水平,并与其他养殖海湾相比也处于较低水平。
对桑沟湾主要养殖区进行24h定点连续监测,掌握不同养殖环境下叶绿素a浓度的昼夜变化规律,从而分析不同养殖活动对该养殖地点的海水水质的影响状况。海草区叶绿素a浓度白天表层高于底层,而夜间则相反;网箱区叶绿素a浓度底层均高于表层;贝类区叶绿素a浓度表层均高于底层;海带区叶绿素a浓度表底层叶绿素a浓度呈现出升降交替的规律。四个连续监测站点的叶绿素a浓度昼夜变化规律各不相同,说明不同养殖活动影响叶绿素a浓度的升降。海草区叶绿素a浓度昼夜的改变可能是受到潮汐作用以及浮游植物光合作用周期的共同影响[16-17]。网箱养殖底部沉积物中富含无机氮、无机磷和硫化物,使微生物活动增强,加速沉积物中氮、磷等营养盐的释放,使水体碳、氮、磷负荷升高[18-20],桑沟湾春季网箱区内大量养殖鲆鲽类,以及投饵使用冻杂鱼和肉食性饲料,鲆鲽类等均属于底栖鱼类[21-22],在网箱中大都处于底层,所以高密度的网箱鱼类的生理代谢活动产生的代谢废物,再加上残饵、粪便等均沉降到底层,为该养殖区底层浮游植物提供了充足的营养盐,使得网箱区叶绿素a浓度底层高于表层。滤食性贝类的摄食行为主要是通过选择性摄食周围海水中以浮游植物为主的悬浮有机物颗粒方式来进行的[23]。桑沟湾浮游植物摄食者主要有养殖贝类和浮游动物,而贝类的摄食可显著改变浮游植物生物量的分布[24]。而本研究中贝类养殖区的表层叶绿素a浓度白天高于夜间,说明叶绿素a浓度受滤食性贝类摄食活动和浮游植物光合作用周期的共同影响,即高密度贝类养殖区夜间的摄食活动能有效降低叶绿素a浓度。另外,水体混浊度也是影响浮游植物的重要因素[20],尤其是底层的叶绿素a浓度变化,所以在今后的研究中应增加对桑沟湾海域水体混浊度数据的监测。
桑沟湾四个主要养殖区叶绿素a浓度整体趋势:网箱区>贝类区>海带区和海草区。大型海藻养殖可以净化养殖废水、控制水域富营养化、调控水域生态平衡[25],而桑沟湾海草区也有较高密度的野生大叶藻[26-27],海草床可以加速悬浮颗粒物的沉降, 增加水体透明度, 净化水质, 加速滨海生态系统中的营养循环等等[28-29],所以桑沟湾春季海草区和海带区的海草和海带生长可能与浮游植物形成竞争,导致这两个海区的叶绿素a浓度显著低于网箱区和贝类区,这也说明了海草区和海带区在桑沟湾养殖系统中起着净化水质、调控水域生态平衡的重要作用。
3.2 叶绿素a的影响因素营养盐对于浮游植物生长的影响是迅速的、灵敏的[30-31]。桑沟湾航线表层、海带区表层以及网箱区底层叶绿素a浓度均与硅酸盐浓度呈较显著的正相关关系。Redfield定律认为藻类健康生长及生理平衡所需的氮磷原子比率为16∶1,硅氮原子比率为1[32],当氮磷比超过16∶1,磷被认为是限制性因素,硅氮比低于1时,硅被认为是限制性因素。从表 2中看出航线调查及四个海区氮磷比值均大于16,硅氮比则小于1,但网箱区和海草区浮游植物存在磷限制程度要强于贝类区和海带区,四个不同养殖区浮游植物均存在硅限制,且活性硅酸盐的浓度也低于浮游植物生长的阈值(2μmol/L)[33],这说明磷元素和硅元素是影响桑沟湾春季浮游植物生长繁殖的限制性因素。从图 7和图 8中可以看出,2014年桑沟湾春季溶解无机氮低于国家一类水质标准,与历史同期相比处于中等水平[13, 16, 34],溶解无机磷则处于国家一类水质标准和二类水质标准,高于历史同期水平,但活性硅酸盐与历史同期相比则处于较低水平,这也从一定程度上反映出硅酸盐是影响桑沟湾春季浮游植物生长的限制性因素。李超伦等研究表明桑沟湾浮游植物种类主要由硅藻类(74种)和甲藻类(11种)组成,硅藻是绝对优势种(春季平均为63. 0×104个/m2)[10],而硅酸盐是硅藻的必需营养元素,除了作为细胞壁结构成分外,还参与光合色素合成、蛋白质合成、DNA合成和细胞分裂等多种代谢和生长过程[35]。孙珊、曲克明等研究表明春季硅浓度低于浮游植物生长的最低阈值,也是一个潜在的限制因素[12, 36]。另外还有研究表明,由于Si结合于硅藻壳中很难再生,并且在水体中的再循环速率比N和P慢得多[37]。本研究结果与前者的研究结论相吻合,即桑沟湾春季海区硅酸盐和温度成为该海域浮游植物主要生长限制因子。

|
| 图 7 2014年桑沟湾春季溶解无机氮、磷酸盐与历史同期数据比较 Fig. 7 The mean comparison of DIN and phosphate with historical data in spring 2014 of Sanggou Bay |

|
| 图 8 2014年桑沟湾春季活性硅酸盐与历史同期比较 Fig. 8 The mean comparison of silicate with historical data in spring 2014 of Sanggou Bay |
桑沟湾春季航线调查显示(图 9),春季湾内水温比湾口回升快,从湾底到湾口,水温逐渐降低,湾外21#站位水温高于1#站位,而盐度变化较小。航线表层叶绿素a浓度与温度呈较显著的正相关关系(图 5),这与郝华林、张继红等研究结果相一致[1-2]。硅藻喜低温,最适合的温度通常低于18℃[38-39],而桑沟湾春季水温在11—18℃,是硅藻生长的理想温度,能促进其大量繁殖,形成明显的优势种。

|
| 图 9 桑沟湾春季航线温度、盐度数据 Fig. 9 The temperature and Salt of lane 1 and 2 in spring 2014 of Sanggou Bay |
潮汐作用是影响沿岸海域水交换的主要因素,同时也是影响叶绿素a浓度变化的因素之一,但在不同海域所表现的叶绿素a变化特征也不尽相同。彭安国等研究厦门湾叶绿素a随潮汐的变化表明该海区表底层叶绿素a浓度均为高潮>低潮[40]。海带区表层叶绿素a浓度高潮时大于低潮,可能是由于桑沟湾大面积海带和贝类养殖等筏式养殖的影响,阻碍了海水流速,降低了桑沟湾内的水交换能力,而海带区处于桑沟湾湾口附近,海水交换能力较强,从而导致了海带区叶绿素a浓度高潮时大于低潮,这与彭安国等研究相一致。海草区表层叶绿素a浓度低潮时大于高潮,可能是海水涨潮时,大量密集的海草床具有缓冲减弱水流并且加速悬浮颗粒物的沉降、增加水体透明度以及净化水质等功能,从而加速浮游植物等颗粒物的沉降,导致表层叶绿素a浓度低潮时大于高潮。
本文的研究不足之处只涉及到一个季节,不能得出桑沟湾主要养殖区叶绿素a浓度分布特征的季节动态变化,今后有待于加强对其季节动态变化以及初级生产力估算的研究,以便为多元养殖模式下养殖水域固碳潜力的计量及估算提供数据支持。
| [1] | 张继红, 方建光, 蒋增杰, 王巍, 王诗欢, 孙松. 獐子岛养殖水域叶绿素含量时空分布特征及初级生产力季节变化[J]. 海洋水产研究 , 2008, 29 (4) : 22–29. |
| [2] | 郝林华, 孙丕喜, 郝建民, 杜蓓蓓, 张新军, 徐玉珊, 毕进红. 桑沟湾海域叶绿素a的时空分布特征及其影响因素研究[J]. 生态环境学报 , 2012, 21 (2) : 338–345. |
| [3] | 刘子琳, 陈建芳, 刘艳岚, 高生泉, 李宏亮, 张海生. 2008年夏季白令海粒度分级叶绿素a和初级生产力[J]. 海洋学报 , 2011, 33 (3) : 148–157. |
| [4] | Sara G, Mazzola A. The carrying capacity for Mediterranean bivalve suspension feeders:evidence from analysis of food availability and hydrodynamics and their integration into a local mode1[J]. Ecological Modelling , 2004, 179 : 281–296. DOI:10.1016/j.ecolmodel.2004.03.005 |
| [5] | Jose L R, Smaal A C, Scholten H. A growth model of the cockle (Cerastoderma edule L.) tested in the Oosterschelde estuary(The Netherlands)[J]. Journal of Sea Research , 2005, 54 : 276–298. DOI:10.1016/j.seares.2005.06.001 |
| [6] | 李斌, 谷伟丽, 靳洋, 马元庆, 白艳艳, 孙珊, 王文杰, 刘小静. 烟台四十里湾叶绿素a和初级生产力的分布特征[J]. 渔业科学进展 , 2012, 33 (2) : 19–23. |
| [7] | 李建军, 毕远溥. 小窑湾贝类养殖对叶绿素a及初级生产力的影响[J]. 水产科学 , 2003, 22 (3) : 7–25. |
| [8] | 孙晓霞, 孙松, 张永山, 张芳. 胶州湾叶绿素a及初级生产力的长期变化[J]. 海洋与湖沼 , 2011, 42 (5) : 654–661. |
| [9] | 薛克, 于清深, 滕利平. 大窑湾牡蛎养殖区叶绿素a分布和初级生产力估算[J]. 水产科学 , 2002, 21 (4) : 4–13. |
| [10] | 李超伦, 张永山, 孙松, 吴玉霖, 方建光, 张继红. 桑沟湾浮游植物种类组成、数量分布及其季节变化[J]. 渔业科学进展 , 2010, 31 (4) : 1–8. |
| [11] | 慕建东, 董玮, 陈碧鹃, 王巍, 方建光, 唐学玺. 桑沟湾浮游植物生态特征[J]. 渔业科学进展 , 2009, 30 (3) : 91–96. |
| [12] | 孙珊, 刘素美, 任景玲, 张继红, 蒋增杰. 桑沟湾养殖海域营养盐和沉积物-水界面扩散通量研究[J]. 海洋学报(中文版) , 2010, 32 (6) : 108–117. |
| [13] | 张继红, 王巍, 韩婷婷, 刘顶海, 方建光, 蒋增杰, 刘新杰, 张新军, 连岩. 桑沟湾春季营养盐分布特征及赤潮暴发诱因[J]. 水产学报 , 2012, 36 (1) : 132–139. |
| [14] | 张继红, 吴文广, 任黎华, 韩婷婷, 王巍, 方建光. 桑沟湾表层水pCO2的季节变化及影响因素分析[J]. 渔业科学进展 , 2013, 34 (1) : 57–64. |
| [15] | 邹芳睿.渤海湾分级叶绿素对营养盐浓度变化的响应.天津:天津大学, 2009, 3-10. |
| [16] | 宋云利, 崔毅, 孙耀, 方建光, 孙慧玲, 杨琴芳, 匡世焕. 桑沟湾养殖海域营养状况及其影响因素分析[J]. 海洋水产研究 , 1996, 17 (2) : 41–51. |
| [17] | 张学雷, 朱明远, 李瑞香, 王宗灵. 贝类养殖环境的多参数同步连续监测[J]. 海洋科学进展 , 2004, 22 (3) : 340–345. |
| [18] | 蒋增杰, 崔毅, 陈碧鹃. 唐岛湾网箱养殖区沉积物一水界面溶解无机氮的扩散通量[J]. 环境科学 , 2007, 28 (5) : 1001–1005. |
| [19] | Kalantzi L, Karakassis L. Benthic impacts of fish farming: meta-analysis of com unity and geochemical data[J]. Marine Pollution Bulletin , 2006, 52 (5) : 484–493. DOI:10.1016/j.marpolbul.2005.09.034 |
| [20] | 粟丽, 朱长波, 张汉华, 李卓佳, 余招龙. 对虾池塘网箱养殖罗非鱼期间水体悬浮颗粒物的动态及对罗非鱼生长和存活的影[J]. 中国水产科学 , 2012, 19 (2) : 256–264. |
| [21] | 唐贤明, 刘天密, 隋曌, 李向民. 漠斑牙鲆养殖技术之三海南漠斑牙鲆养殖技术[J]. 中国水产 , 2007, 375 (2) : 55–56. |
| [22] | 方镇熔. 大西洋牙鲆养殖模式与技术[J]. 养殖与饲料 , 2009, 2 : 14–16. |
| [23] | 蒋增杰, 方建光, 门强, 王巍. 桑沟湾贝类筏式养殖与环境相互作用研究[J]. 南方水产 , 2006, 2 (1) : 23–29. |
| [24] | Nunes J P, Ferreira J G, Gazeau F, Lencart-Silva J, Zhang X L, Zhu M Y, Fang J G. A model for sustainable management of shellfish polyculture in coastal bays[J]. Aquaculture , 2003, 219 (1/4) : 257–277. |
| [25] | 毛玉泽, 颜婷茹, 朱玲, 蒋增杰, 王巍, 方建光. 桑沟湾不同养殖区水体微生物群落结构特征[J]. 中国水产科学 , 2013, 20 (4) : 824–83. |
| [26] | 高亚平, 方建光, 唐望, 张继红, 任黎华, 杜美荣. 桑沟湾大叶藻海草床生态系统碳汇扩增力的估算[J]. 渔业科学进展 , 2013, 34 (1) : 17–21. |
| [27] | 刘炳舰, 周毅, 刘旭佳, 刘鹏, 张晓梅. 桑沟湾楮岛近岸海域大叶藻生态学特征的基础研究[J]. 海洋科学 , 2013, 1 : 42–48. |
| [28] | Hemminga M A, Harrison P G, Lent F van. The balance of nutrient losses and gains in seagrass meadows[J]. Marine Ecology Progress Series , 1991, 71 : 85–96. DOI:10.3354/meps071085 |
| [29] | 高亚平, 方建光, 张继红, 李峰, 毛玉泽, 杜美荣. 桑沟湾大叶藻附着生物的季节变化[J]. 渔业科学进展 , 2010, 31 (4) : 59–64. |
| [30] | 鲁北伟, 王荣, 王文琪. 春季东海不同水域的表层叶绿素含量[J]. 海洋科学 , 1997 (5) : 53–55. |
| [31] | Uye S. Replacement of large copepods by small ones with eutrophication of embayments: cause and consequence[J]. Ecology and Morphology of Copepods , 1994, 292-293 (1) : 513–519. |
| [32] | Redfield A C, Ketchum B H, Richard F A. The influence of organism on the composition of seawater[J]. The Sea , 1963, 2 : 26–77. |
| [33] | Fisher Thomas R, Peele Emily R, Ammerman James W, Harding Lawrence W Jr. Nutrient limitation of phytoplankton in Chesapeake Bay[J]. Marine Ecology Progress Series , 1992, 82 : 51–63. DOI:10.3354/meps082051 |
| [34] | 张继红, 蒋增杰, 王巍, 邹建, 薛素艳, 方建光, 连岩, 张新军, 刘新杰, 周颖敏. 桑沟湾营养盐时空分布及营养盐限制分析[J]. 渔业科学进展 , 2010, 31 (4) : 16–25. |
| [35] | 梁君荣, 陈丹丹, 高亚辉, 程金凤. 海洋硅藻硅质细胞壁结构的形成机理研究概述[J]. 海洋学报(中文版) , 2010, 32 (5) : 1–8. |
| [36] | 曲克明, 宋云利, 徐勇, 孙耀, 方建光. 桑沟湾养殖水域春季和夏季营养限制的现场实验[J]. 海洋环境科学 , 2008, 27 (2) : 124–128. |
| [37] | Conley Daniel J, Schelske Claire L, Stoermer Eugene F. Modification of the biogeochemical cycle of silica with eutrophication[J]. Marine Ecology Progress Series , 1993, 101 : 179–192. DOI:10.3354/meps101179 |
| [38] | Wasmund Norbert, Tuimala Jarno, Suikkanen Sanna, Vandepitte Leen, Kraberg Alexandra. Long-term trends in phytoplankton composition in the western and central BalticSea[J]. Journal of Marine Systems , 2011, 87 (2) : 145–159. DOI:10.1016/j.jmarsys.2011.03.010 |
| [39] | 周然, 彭士涛, 覃雪波, 石洪华, 丁德文. 渤海湾浮游植物与环境因子关系的多元分析[J]. 环境科学 , 2013, 34 (3) : 864–873. |
| [40] | 彭安国, 黄奕普, 陈敏, 刘广山, 邱雨生. 厦门湾水体中不同粒级颗粒物、Chl-a和234Th随潮汐的变化及其海洋学意义[J]. 台湾海峡 , 2004, 23 (4) : 403–409. |