 2016, Vol. 36
2016, Vol. 36文章信息
- 卢增斌, 欧阳芳, 张永生, 关秀敏, 门兴元
- LU Zengbin, OUYANG Fang, ZHANG Yongsheng, GUAN Xiumin, MEN Xingyuan.
- 华北平原地区景观格局对麦田害螨种群数量的影响
- Impacts of landscape patterns on populations of the wheat mites, Petrobia latens (Müller) and Penthaleus major (Duges), in the North China Plain
- 生态学报[J]. 2016, 36(14): 4447-4455
- Acta Ecologica Sinica[J]. 2016, 36(14): 4447-4455
- http://dx.doi.org/10.5846/stxb201412112457
-
文章历史
- 收稿日期: 2014-12-11
- 网络出版日期: 2015-12-08





2. 中国科学院动物研究所, 农业虫害鼠害综合治理研究国家重点实验室, 北京 100101;
3. 湖南农业大学植物保护学院, 长沙 410128;
4. 山东省农业厅植物保护总站, 济南 250100
2. State Key Laboratory of Integrated Management of Pest Insects and Rodents, Institute of Zoology, Chinese Academy of Sciences, Beijing 100101, China;
3. College of Plant Protection, Hunan Agricultural University, Changsha 410128, China;
4. Shandong Plant Protection Station, Jinan 250100, China
农田景观是由耕地、草地、林地、树篱等不同斑块组成的镶嵌体,在景观尺度上表现为物种生存的各类缀化栖地的空间网络结构[1]。近半个世纪以来,农业生产的规模化经营导致农业用地不断扩张、作物种植面积持续扩大和非作物生境的大规模减少,致使农田景观格局日趋单一,而农田景观格局的变化势必会影响到景观中害虫的发生和危害及自然天敌的生态控制功能[2-3]。目前大量的研究表明非作物生境比例比较高的农田景观中自然天敌不仅多样性丰富且种群数量多[2],这是由于非作物生境(农田边缘、休耕地、草地和林地)能够为自然天敌提供必要的寄主、食物和度过不良环境的庇护所[2, 4]。然而害虫种群对景观复杂性响应的研究相对较少且没有统一的认识[5],即便在同一个研究系统中不同害虫种类对景观复杂性的响应也并不一致[6-8],如麦长管蚜和禾谷缢管蚜种群数量的最高值在复杂农业景观中超过简单农业景观,而麦二叉蚜却恰恰相反[8],并且不同景观因子对害虫的影响也并不一致,如裸地、草地和林地对麦蚜种群数量有促进作用,而塑料大棚却能抑制其种群增长[9]。因此,阐明不同景观因子对害虫种群的影响,对于通过生境管理进行害虫控制具有重要的理论和实践意义。
农田景观格局是生物自然过程与人类干扰相互作用形成的, 是各种复杂的自然和社会条件相互作用的结果,同时农田景观格局也制约和影响着各种生态过程[10]。农田景观格局的主要特征可以采用“质、量、形、度”来概括,其中“质”表示农田景观中不同的景观组成,“量”表示不同类型斑块的大小和面积比例等,“形”表示不同斑块类型的形状和排列方式,“度”则反映农田景观格局变化的时间和空间尺度[3]。当然,前人的研究更多集中在农田尺度条件下非作物生境的种类和面积对害虫的影响[2, 4, 8-9],极少关注于更大空间尺度条件下景观格局和害虫发生之间的关系,而通过大尺度多景观因子(质、量、形、度)与害虫种群发生的相关性分析更能明确景观格局因子对农田害虫种群的驱动作用。
小麦是我国华北平原地区主要的粮食作物之一。麦螨是小麦上的主要害虫,主要包括麦圆叶爪螨(Penthaleus major (Duges))和麦岩螨(Petrobia latens (Müller)),两者均属于蛛形纲,蜱螨目,危害十分严重,主要于小麦苗期至抽穗期吸食叶片汁液,常常造成小麦不能抽穗而枯死,严重影响小麦产量[11-12],如阜阳市麦田害螨年发生面积10万hm2,造成小麦产量损失10%—20%[13]。在我国的华北地区,麦田害螨在农田中呈现区域性分布的特点,某些区域发生严重,而另一些区域发生却较轻[11]。这种分布特点可能与区域性景观因子的影响有关,而具体涉及的景观因子尚不明确。
有鉴于此,本文以华北平原地区的山东省为研究区域,24个县级单元为样点,通过对卫星遥感影像和土地覆盖分类数据的分析,获取了样点单元的农田景观格局指数。同时定点调查了样点单元2008—2010年麦田害螨的种群发生量。通过分析景观格局指数和麦田害螨年平均发生量之间的相关关系,以明确影响麦田害螨发生和危害的主要景观因子,为基于农田景观格局的麦田害螨种群生态调控提供理论和技术支撑。
1 材料和方法 1.1 研究区域研究区域为山东省24个地市,分别为淄川、临淄、枣庄、山亭、滕州、东营、垦利、利津、广饶、莱阳、潍坊、临朐、诸城、寿光、安丘、金乡、嘉祥、文登、日照、乐陵、无棣、菏泽、郓城、定陶等24个县市,这24个地区基本覆盖了整个山东的各个区域。
山东省是我国小麦的重要产区和粮食生产基地,小麦年种植面积333万hm2左右,属于暖温带季风气候区,年平均温度12—16℃,≥15℃的有效积温为3300—3800℃,≥10℃的持续天数为190—200 d,降雨量600—1200 mm,年日照时数为2000—2900 h。
1.2 数据获取 1.2.1 麦田害螨发生量的获取根据中华人民共和国农业行业标准—麦蜘蛛测报调查规范(NY/T 615—2012),在山东省每个样点县范围内选取代表性强的不同(景观)类型麦田10块,每块小麦田按照对角线5点取样调查,每个样点调查33.3 cm单行长的麦田害螨数量。每年(2008—2010年)春季麦田害螨活动的高峰期进行调查,每年调查2次。取样时将33.3 cm×17.0 cm的白瓷盘铺在取样点的麦根际,将麦苗轻轻压弯拍打,然后计数,调查麦螨种类及其数量,最后折算为每个县域的平均发生量。以2008—2010年山东省24个县域麦田害螨春季的年平均发生量为基础建立了麦田害螨在全省分布的数据库。
1.2.2 卫星遥感影像和土地覆盖分类本研究使用的遥感数据源为2010年Landsat TM/ETM的中分辨率卫星影像(10—30m)。运用ENVI 5.0遥感图像处理软件,对图像进行几何精校正、图像裁剪和图像增强等预处理。在大量野外实地调查的基础上,建立解译图谱库。对遥感影像进行分类解译,获得2010年山东省土地覆被分类的栅格数据。将山东省区域内土地覆盖类型归为8类:森林、草地、湿地、水体、耕地、园地、居住工业道路交通、其他用地。土地覆被分类的栅格数据精度为30 m×30 m。在地理信息系统(ARCGIS 10.2)的支持下,根据山东省内140个县级单位行政区的矢量数据,把全省土地覆被分类的栅格数据分割成140个景观类型分类图。
1.3 景观格局分析基于山东省140个县域单位的土地覆被分类栅格数据,利用景观格局分析软件FRAGSTATS 4.2,分别在景观水平和斑块水平上计算每个县域的17个景观格局指数。17个景观格局指数的计算如下:(1)斑块类型(PT),省域范围下土地覆盖类型,主要包括森林、草地、湿地、水体、耕地、园地、居住工业道路交通和其他用地;(2)斑块丰富度(PR),PR=m,式中m为景观中不同斑块类型的总数;(3)斑块丰富度密度(PRD),PRD=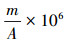

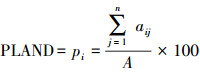
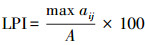
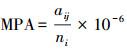
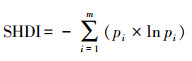
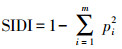
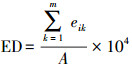
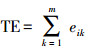
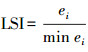
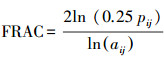
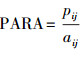
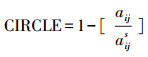

利用PASW Statistics 18.0(2009, IBM, Armonk, NY, USA)对麦田害螨的发生量进行描述性统计。采用R 3.0.3计算景观格局指数与麦螨发生量之间的Pearson相关系数并进行显著性分析。显著性水平设置为P=0.05。
2 结果 2.1 麦田害螨种群数量特征及其分布以山东省为典型代表的华北平原小麦种植区域中,麦田害螨的发生量呈区域性分布的特点。24个县域麦岩螨种群的平均密度为189.92头/33.3 cm单行,最大值为736.00头/33.3 cm单行,最小值为0头/33.3 cm单行(图 1A),淄川、日照和临淄的年平均发生量最大,而无棣、滕州和枣庄的发生量最低,金乡和嘉祥在调查的3年中没有发现麦岩螨(图 2);麦圆叶爪螨的平均密度为250.87头/33.3 cm单行,最大值为579.00头/33.3 cm单行,最小值为15.21头/33.3 cm单行(图 1B),嘉祥、山亭和金乡的发生量最大,而文登、莱阳、东营和广饶的发生量最低(图 2)。

|
| 图 1 以山东省为典型代表的华北平原小麦种植区麦岩螨(A)和麦圆叶爪螨(B)的种群数量特征 Fig. 1 The characteristic of populations of Petrobia latens(A)and Penthaleus major(B)in Shandong province |

|
| 图 2 山东省24县域麦岩螨和麦圆叶爪螨的种群数量 Fig. 2 The density of Petrobia latens and Penthaleus major in each sampling unit in Shandong province |
麦岩螨和麦圆叶爪螨种群的发生同省域范围的斑块类型以及县域尺度下的斑块丰富度和斑块丰富度密度没有相关关系(表 1)。
| 景观组成:质 Landscape composition |
麦岩螨Petrobia latens | 麦圆叶爪螨Penthaleus major | |||
| r | P | r | P | ||
| 斑块类型patch type (PT) | |||||
| 省域范围Province | 森林,草地,湿地,水体,耕地,园地,居工道*,其他 | ||||
| 斑块丰富度patch richness (PR) | |||||
| 县域范围County | 0.1798 | 0.4006 | -0.2949 | 0.1619 | |
| 斑块丰富度密度patch richness density (PRD) | |||||
| 县域范围County | 0.1783 | 0.4046 | 0.1158 | 0.5900 | |
| *居工道包括居住、工业、道路交通等用地 | |||||
麦岩螨种群的发生量与森林的最大斑块面积指数(r=0.4721,P=0.0198)和平均斑块面积(r=0.4723,P=0.0198)呈显著正相关。麦圆叶爪螨的发生量与水体的总面积(r=-0.4439,P=0.0298)、斑块面积比例(r=-0.4933,P=0.0143)、最大斑块面积指数(r=-0.4295,P=0.0362)以及县域范围的平均斑块面积(r=-0.4102,P=0.0465)呈显著负相关,而与县域范围(r=0.5005, P=0.0127)、森林(r=0.4898, P=0.0151)、耕地(r=0.4112, P=0.0459)和居工道(r=0.5420, P=0.0062)的斑块密度呈显著正相关(表 2)。
| 景观构成:量Landscape quantity | 麦岩螨Petrobia latens | 麦圆叶爪螨Penthaleus major | |||
| r | P | r | P | ||
| 斑块类型总面积total class area(CA) | |||||
| 县域范围County | |||||
| 森林Forest | 0.1165 | 0.5877 | 0.0612 | 0.7762 | |
| 草地Grassland | 0.0686 | 0.7676 | 0.0292 | 0.9000 | |
| 水体Water | -0.1613 | 0.4515 | -0.4439 | 0.0298 | |
| 耕地Farmland | -0.2252 | 0.2901 | -0.1858 | 0.3848 | |
| 居工道RIT * | -0.1295 | 0.5464 | -0.1900 | 0.3740 | |
| 斑块面积比例percentage of landscape (PLAND) | |||||
| 县域范围County | |||||
| 森林Forest | 0.2502 | 0.2382 | 0.2828 | 0.1805 | |
| 草地Grassland | 0.1423 | 0.5383 | 0.1683 | 0.4658 | |
| 水体Water | -0.1304 | 0.5437 | -0.4933 | 0.0143 | |
| 耕地Farmland | -0.2199 | 0.3018 | 0.1925 | 0.3674 | |
| 居工道RIT | 0.1443 | 0.5011 | 0.0693 | 0.7476 | |
| 斑块密度patch density(PD) | |||||
| 县域范围County | 0.1290 | 0.5480 | 0.5005 | 0.0127 | |
| 森林Forest | 0.0197 | 0.9274 | 0.4898 | 0.0151 | |
| 草地Grassland | 0.2720 | 0.2329 | 0.2416 | 0.2914 | |
| 水体Water | -0.3250 | 0.1212 | 0.0389 | 0.8566 | |
| 耕地Farmland | 0.1073 | 0.6178 | 0.4112 | 0.0459 | |
| 居工道RIT | -0.0293 | 0.8919 | 0.5420 | 0.0062 | |
| 最大斑块面积指数largest patch index(LPI) | |||||
| 县域范围County | -0.0680 | 0.7521 | 0.2116 | 0.3208 | |
| 森林Forest | 0.4721 | 0.0198 | 0.2422 | 0.2542 | |
| 草地Grassland | -0.0484 | 0.8351 | 0.2284 | 0.3194 | |
| 水体Water | -0.1396 | 0.5153 | -0.4295 | 0.0362 | |
| 耕地Farmland | -0.1212 | 0.5727 | 0.1939 | 0.3640 | |
| 居工道RIT | 0.2112 | 0.3220 | -0.1863 | 0.3834 | |
| 平均斑块面积mean patch area (MPA) | |||||
| 县域范围County | -0.0850 | 0.6928 | -0.4102 | 0.0465 | |
| 森林Forest | 0.4723 | 0.0198 | -0.1988 | 0.3517 | |
| 草地Grassland | -0.1887 | 0.4126 | -0.0310 | 0.8939 | |
| 水体Water | -0.1699 | 0.4275 | -0.2869 | 0.1741 | |
| 耕地Farmland | -0.1017 | 0.6363 | -0.1069 | 0.6192 | |
| 居工道RIT | 0.1952 | 0.3607 | -0.1069 | 0.6192 | |
| Shannon多样性指数shannon′s diversity index (SHDI) | |||||
| 县域范围County | 0.1957 | 0.3594 | -0.3172 | 0.1309 | |
| Simpson多样性指标simpson′s diversity index (SIDI) | |||||
| 县域范围County | 0.1798 | 0.4006 | -0.2629 | 0.2146 | |
| *居工道包括居住、工业、道路交通等用地 | |||||
麦岩螨种群的发生量与水体的景观形状指数(r=-0.4443,P=0.0296)以及森林的分形指数(r=-0.4097,P=0.0468)、周长面积比(r=-0.4148,P=0.0439)和近圆形形状指数(r=-0.4493,P=0.0276)呈显著负相关,而与森林的聚集度指数(r=0.4172,P=0.0426)呈显著正相关。
麦圆叶爪螨种群的发生量与县域范围(r=0.5219,P=0.0089)、森林(r=0.4671,P=0.0214)、耕地(r=0.5534,P=0.0050)和居工道(r=0.5288,P=0.0079)的边界密度,水体的分形指数(r=0.4088,P=0.0473)和周长面积比(r=0.7035,P=0.0001),森林的分形指数(r=0.4229,P=0.0395)和近圆形指数(r=0.5821,P=0.0028),县域范围(r=0.5169,P=0.0097)和居工道(r=0.4156,P=0.0434)的周长面积比呈显著正相关,而与水体的总边缘长度(r=-0.4352,P=0.0336)以及县域范围(r=-0.5219,P=0.0089)、水体(r=-0.7022,P=0.0001)、居工道(r=-0.4255,P=0.0382)的聚集度指数呈显著负相关(表 3)。
| 景观结构:形 Landscape configuration |
麦岩螨Petrobia latens | 麦圆叶爪螨Penthaleus major | |||
| r | P | r | P | ||
| 边界密度edge density (ED) | |||||
| 县域范围County | 0.0957 | 0.6563 | 0.5219 | 0.0089 | |
| 森林Forest | -0.0072 | 0.9733 | 0.4671 | 0.0214 | |
| 草地Grassland | 0.2351 | 0.3051 | 0.1779 | 0.4405 | |
| 水体Water | -0.3942 | 0.0566 | -0.2029 | 0.3416 | |
| 耕地Farmland | 0.0534 | 0.8043 | 0.5534 | 0.0050 | |
| 居工道RIT * | 0.0311 | 0.8854 | 0.5288 | 0.0079 | |
| 总边缘长度total edge (TE) | |||||
| 县域范围County | -0.0720 | 0.7383 | 0.0320 | 0.8821 | |
| 森林Forest | -0.0558 | 0.7955 | 0.1801 | 0.3997 | |
| 草地Grassland | 0.1315 | 0.5698 | 0.0423 | 0.8557 | |
| 水体Water | -0.3316 | 0.1134 | -0.4352 | 0.0336 | |
| 耕地Farmland | -0.1020 | 0.6354 | 0.0173 | 0.9361 | |
| 居工道* RIT | -0.1855 | 0.3856 | 0.0994 | 0.6441 | |
| 景观形状指数landscape shape index (LSI) | |||||
| 县域范围County | 0.0133 | 0.9510 | 0.2522 | 0.2344 | |
| 森林Forest | -0.2720 | 0.1985 | 0.3260 | 0.1201 | |
| 草地Grassland | 0.2241 | 0.3288 | 0.0514 | 0.8250 | |
| 水体Water | -0.4443 | 0.0296 | 0.2940 | 0.1632 | |
| 耕地Farmland | 0.0707 | 0.7427 | 0.2365 | 0.2659 | |
| 居工道RIT | -0.1616 | 0.4506 | 0.2587 | 0.2222 | |
| 分形指数fractal dimension index (FRACT) | |||||
| 县域范围County | 0.0643 | 0.7654 | 0.3454 | 0.0983 | |
| 森林Forest | -0.4097 | 0.0468 | 0.4229 | 0.0395 | |
| 草地Grassland | -0.0299 | 0.8977 | -0.0243 | 0.9166 | |
| 水体Water | -0.1924 | 0.3677 | 0.4088 | 0.0473 | |
| 耕地Farmland | 0.2983 | 0.1569 | 0.2402 | 0.2582 | |
| 居工道RIT | 0.0790 | 0.7137 | -0.2040 | 0.3390 | |
| 周长面积比perimeter area ratio (PARA) | |||||
| 县域范围County | 0.1064 | 0.6207 | 0.5169 | 0.0097 | |
| 森林Forest | -0.4148 | 0.0439 | 0.3820 | 0.0655 | |
| 草地Grassland | -0.0103 | 0.9646 | 0.3017 | 0.1838 | |
| 水体Water | -0.3048 | 0.1475 | 0.7035 | 0.0001 | |
| 耕地Farmland | 0.2823 | 0.1814 | 0.3897 | 0.0598 | |
| 居工道RIT | -0.1384 | 0.5189 | 0.4156 | 0.0434 | |
| 近圆形形状指数related circumscribing circle (CRICLE) | |||||
| 县域范围County | -0.0514 | 0.8116 | -0.1238 | 0.5643 | |
| 森林Forest | -0.4493 | 0.0276 | 0.5821 | 0.0028 | |
| 草地Grassland | -0.0510 | 0.8263 | -0.1378 | 0.5515 | |
| 水体Water | 0.0519 | 0.8098 | 0.5453 | 0.0058 | |
| 耕地Farmland | -0.0178 | 0.9344 | -0.3057 | 0.1463 | |
| 居工道RIT | 0.0519 | 0.8096 | -0.3057 | 0.1463 | |
| 聚集度指数contiguity index (CONTIG) | |||||
| 县域范围County | -0.1028 | 0.6326 | -0.5219 | 0.0089 | |
| 森林Forest | 0.4172 | 0.0426 | -0.3746 | 0.0713 | |
| 草地Grassland | 0.0165 | 0.9433 | -0.3005 | 0.1857 | |
| 水体Water | 0.3051 | 0.1471 | -0.7022 | 0.0001 | |
| 耕地Farmland | -0.2817 | 0.1824 | -0.3880 | 0.0610 | |
| 居工道RIT | 0.1500 | 0.4842 | -0.4255 | 0.0382 | |
| *居工道包括居住、工业、道路交通等用地 | |||||
本文研究结果表明景观因子的“量”和“形”对麦田害螨种群均具有显著的影响,且两种害螨对景观因子的响应并不一致,森林类景观因子和水体类景观因子分别对麦岩螨和麦圆叶爪螨的种群有显著影响。这与其他人的结果相一致即不同种类的害虫,可能由于其食性、活动能力、扩散能力以及对环境条件要求的不同导致对农田景观结构的响应不一样[6-9]。同时本文全面评价了景观格局的“质、量、形、度”对麦田害螨种群的影响,克服了以往的研究中主要关注于非作物生境的类型和面积(或比例)对害虫和自然天敌影响[2, 4],且与农田景观尺度相比,采用县域尺度分析更加能够反映出整个大区域景观格局对麦田害螨种群的影响,对将来优化景观格局进行害虫生态管理具有重要的指导意义。
麦岩螨的发生量受到森林类景观因子的影响,该现象可能与非作物生境为麦岩螨提供食物和庇护场所有关。麦岩螨除主要为害小麦外,还能为害桃、苹果、柳、桑、槐等木本植物和红茅草、马绊草、苦菜子等杂草[15]。在现代化的农田生态系统中,作物生境占据着主要地位,然而由于作物是一种短时收获的植物,生活在其中的害虫和天敌必需在作物建立之前或者收割之后寻找适宜的自然生境或非作物生境度过不利的环境条件,当作物生境条件适宜时,害虫或天敌又重新迁入到作物生境中。在我国山东的冬小麦种植区,小麦和周围的杂草以及果树等构成了基本的农田景观,小麦收割之后,大部分麦岩螨的卵在麦田越夏,一部分麦岩螨可以借助风力迁移到杂草和果树上继续为害或以卵越夏,秋天麦岩螨又借助风力从杂草和木本植物迁回到小麦上为害和越冬。在其他的研究体系中也发现非作物生境可以促进作物生境中害虫种群的加重发生,如在果棉混栽区,6月中下旬绿盲蝽迁出果园进入周围的棉田危害[16]。森林和半自然生境面积的增加可以促进油菜地的两种害虫-油菜露尾甲(Meligethes aeneus)和油菜荚叶瘿蚊(Dasineura brassicae)种群增长[17-18],且草地和条带植被面积的增加也可以促进蚜虫[19]和飞虱种群的增加[7, 20]。Yoshioka等发现一种非本地的意大利黑麦草有利于一种当地半翅目害虫Stenotus rubrovittatus的生长[21]。Taki等的研究也表明人工种植的针叶树对果园害虫斯氏珀蝽(Plautia stali)的种群有正向影响[22]。
麦圆叶爪螨的发生量受到水体的影响,该现象可能与麦圆叶爪螨对湿度的需求有关。麦圆叶爪螨喜高湿畏干燥,相对湿度70%以上,表土含水量20%左右最适于其繁殖危害,主要发生于水浇地、低湿或密植麦田,而干旱的麦田发生较轻[15]。湿度可以直接影响到昆虫的生长、发育、繁殖和生存,如湿度是影响绿盲蝽越冬卵的孵化和种群增长的关键因素,70%和80%的湿度能够显著提高卵和若虫的存活率、延长成虫寿命和提高雌成虫产卵量[23-24],而且湿度也可以通过对寄主植物和天敌的作用对害虫产生间接的影响。高湿度条件下植物生长快、嫩绿更适宜害虫取食,同时降雨带来的高湿度可以影响到天敌的存活和搜寻行为,进而间接的影响到对害虫的生物控制作用,如降雨显著地影响到蚜虫寄生蜂Aphidius rosae的搜寻行为和繁殖能力[25]。因此,湿度的增加,一方面可以直接影响到麦圆叶爪螨本身的生物学特性,促进其种群增加,另一方面食物的适宜取食和对天敌活动的限制间接的影响到麦螨种群的发生量。
总之,景观因子与麦田害螨种群的发生密切相关,两种害螨对景观因子的响应不一致可能与其对食物或环境湿度的需求有关,因此在麦田害螨发生严重的区域,应根据害螨的种类采取合理的生境管理策略,抑制害螨种群数量。
| [1] | Turner M G. Spatial and temporal analysis of landscape patterns[J]. Landscape Ecology , 1990, 4 (1) : 21–30. DOI:10.1007/BF02573948 |
| [2] | Bianchi F J J A, Booij C J H, Tscharntke T. Sustainable pest regulation in agricultural landscapes:a review on landscape composition, biodiversity and natural pest control[J]. Proceedings of the Royal Society B-Biological Sciences , 2006, 273 (1595) : 1715–1727. DOI:10.1098/rspb.2006.3530 |
| [3] | 欧阳芳, 戈峰. 农田景观格局变化对昆虫的生态学效应[J]. 应用昆虫学报 , 2011, 48 (5) : 1177–1183. |
| [4] | Landis D A, Wratten S D, Gurr G M. Habitat management to conserve natural enemies of arthropod pests in agriculture[J]. Annual Review of Entomology , 2000, 45 : 175–201. DOI:10.1146/annurev.ento.45.1.175 |
| [5] | Chaplin-Kramer R, O'Rourke M E, Blitzer E J, Kremen C. A meta-analysis of crop pest and natural enemy response to landscape complexity[J]. Ecology Letters , 2011, 14 (9) : 922–932. DOI:10.1111/j.1461-0248.2011.01642.x |
| [6] | Jonsen I D, Fahrig L. Response of generalist and specialist insect herbivores to landscape spatial structure[J]. Landscape Ecology , 1997, 12 (3) : 185–197. DOI:10.1023/A:1007961006232 |
| [7] | Letourneau D K, Goldstein B. Pest damage and arthropod community structure in organic vs.conventional tomato production in California[J]. Journal of Applied Ecology , 2001, 38 (3) : 557–570. DOI:10.1046/j.1365-2664.2001.00611.x |
| [8] | 赵紫华, 石云, 贺达汉, 杭佳, 赵映书, 王颖. 不同农业景观结构对麦蚜种群动态的影响[J]. 生态学报 , 2010, 30 (23) : 6380–6388. |
| [9] | 赵紫华, 王颖, 贺达汉, 关晓庆, 辛明. 麦蚜和寄生蜂对农业景观格局的响应及其关键景观因子分析[J]. 生态学报 , 2012, 32 (2) : 472–482. |
| [10] | 付梅臣, 胡振琪, 吴淦国. 农田景观格局演变规律分析[J]. 农业工程学报 , 2005, 21 (6) : 54–58. |
| [11] | 钟启谦, 魏鸿钧, 田毓启. 麦园蜘蛛(Penthaleus sp.)及麦长腿蜘蛛(Petrobia latens(Müller))的生物学研究[J]. 植物保护学报 , 1963, 2 (3) : 277–284. |
| [12] | 华南农学院. 农业昆虫学(上册). 北京: 农业出版社, 1981. |
| [13] | 许殿武, 于小平, 刘东恒. 阜阳市小麦红蜘蛛发生特点及原因分析[J]. 安徽农业科学 , 2007, 35 (1) : 135–135. |
| [14] | 郑新奇, 付梅臣. 景观格局空间分析技术及其应用. 北京: 科学出版社, 2010. |
| [15] | 仵均祥, 袁国辉, 史树森, 贝纳新. 农业昆虫学. 北京: 中国农业出版社, 2002. |
| [16] | 门兴元, 于毅, 张安盛, 李丽莉, 张思聪.枣一棉生态区绿盲蜷季节性发生与转移规律//植保科技创新与病虫防控专业化——中国植物保护学会2011年学术年会论文集.北京:中国农业科学出版, 2011:1-898. |
| [17] | Zaller J G, Moser D, Drapela T, Schm ger C, Frank T. Effect of within-field and landscape factors on insect damage in winter oilseed rape[J]. Agriculture Ecosystems & Environment , 2008, 123 : 233–238. |
| [18] | Zaller J G, Moser D, Drapela T, Schmöger C, Frank T. Insect pests in winter oilseed rape affected by field and landscape characteristics[J]. Basic and Applied Ecology , 2008, 9 (6) : 682–690. DOI:10.1016/j.baae.2007.10.004 |
| [19] | Meyhöfer R, Klug T, Poehling H M. Are landscape structures important for the colonization of spinach fields by insects?[J]. IOBC/WPRS Bulletin , 2008, 34 : 69–72. |
| [20] | Grilli M P, Bruno M. Regional abundance of a planthopper pest:the effect of host patch area and configuration[J]. Entomologia Experimentalis et Applicata , 2007, 122 (2) : 133–143. DOI:10.1111/eea.2007.122.issue-2 |
| [21] | Yoshioka A, Takada M B, Washitani I. Landscape effects of a non-native grass facilitate source populations of a native generalist bug, Stenotus rubrovittatus, in a heterogeneous agricultural landscape[J]. Journal of Insect Science , 2014, 14 (110) : 1–14. DOI:10.1673/031.014.110 |
| [22] | Taki H, Tabuchi K, Iijima H, Okabe K, Toyama M. Spatial and temporal influences of conifer planted forests on the orchard pest Plautia stali(Hemiptera:Pentatomidae)[J]. Applied Entomology and Zoology , 2014, 49 (2) : 241–247. DOI:10.1007/s13355-013-0242-y |
| [23] | Lu Y H, Wu K M. Effect of relative humidity on population growth of Apolygus lucorum(Heteroptera:Miridae)[J]. Applied Entomology and Zoology , 2011, 46 (3) : 421–427. DOI:10.1007/s13355-011-0058-6 |
| [24] | Pan H S, Liu B, Lu Y H, Desneux N. Identification of the key weather factors affecting overwintering success of Apolygus lucorum eggs in dead host tree branches[J]. PLoS ONE , 2014, 9 (4) : e94190. DOI:10.1371/journal.pone.0094190 |
| [25] | Fink U, Völkl W. The effect of abiotic factors on foraging and oviposition success of the aphid parasitoid, Aphidius rosae[J]. Oecologia , 1995, 103 (3) : 371–378. DOI:10.1007/BF00328627 |