 2016, Vol. 36
2016, Vol. 36文章信息
- 黄甫昭, 丁涛, 李先琨, 郭屹立, 王斌, 向悟生, 文淑均, 李冬兴, 何运林
- HUANG Fuzhao, DING Tao, LI Xiankun, GOU Yili, WANG Bin, XIANG Wusheng, WEN Shujun, LI Dongxing, HE Yunlin.
- 弄岗喀斯特季节性雨林不同群丛物种多样性随海拔的变化
- Species diversity for various associations along an altitudinal gradient in the karst seasonal rainforest in Nonggang
- 生态学报[J]. 2016, 36(14): 4509-4517
- Acta Ecologica Sinica[J]. 2016, 36(14): 4509-4517
- http://dx.doi.org/10.5846/stxb201411232324
-
文章历史
- 收稿日期: 2014-11-23
- 网络出版日期: 2015-10-30





2. 广西友谊关森林生态系统定位观测研究站, 凭祥 532699;
3. 广西师范大学生命科学院, 桂林 541002
2. Guangxi Youyiguan Forest Ecosystem Research Station, Pingxiang 532699, China;
3. College of Life Sciences, Guangxi Normal University, Guilin 541002, China
我国喀斯特分布面积约占国土面积的1/3,是世界上喀斯特分布面积最广的国家,主要分布在以云贵高原为中心的贵州、云南、广西、重庆等地,该区是全球三大喀斯特集中分布区中面积最大、岩溶发育最强烈的地区,也是石漠化最严重和生态环境最脆弱的地区之一[1]。喀斯特地区生态系统脆弱,环境容量小,抗干扰能力差,植被遭到破坏后地表土壤流失迅速,部分区域甚至出现石漠化[2]。如何利用喀斯特地区原生植被治理喀斯特地区石漠化,成为近年来我国众多学者的关注焦点。
原生性喀斯特森林是喀斯特石漠化地区生态重建的重要参考系,深入研究原生性喀斯特森林的群落特征,对揭示其生态系统的运行规律,指导植被恢复具有重要意义。多年来,学者们对喀斯特原生性森林开展了各方面的研究,蒋国芳[3]、宋同清[4]、侯满福[5]、张忠华[6]等对我国亚热带喀斯特常绿落叶阔叶混交林群落物种组成、多样性特征等开展了研究,Polak、Brewer、Felfili分别在印度尼西亚、伯利兹和巴西中部对热带喀斯特森林的植物多样性进行了研究[7-9]。但不同气候带的喀斯特森林具有不同的区系成分、结构和外貌[10],热带、亚热带喀斯特森林的研究结果在北热带喀斯特季节性雨林不一定适用。
北热带喀斯特季节性雨林(tropical karst seasonal rainforest)是我国北热带喀斯特山地具有地带性植被特征的地域性植被[11],该植被分布区内广泛分布着峰丛、槽谷、洼地等地形地貌,小生境复杂多样[2],群落结构也复杂多变、物种多样性从峰丛至洼地变化剧烈。截止目前,关于北热带喀斯特季节性雨林物种多样性特征及其从洼地至峰丛随相对海拔变化规律还鲜有研究,仅有的报道也只是在物种组成特征或在植被分类研究中对其进行概述[12-15]。本文以弄岗15hm2北热带喀斯特季节性雨林样地的调查数据为基础,在对样地进行群丛划分的基础上,采用丰富度指数、物种多样性指数和均匀度指数对不同群丛的物种多样性及其随海拔变化进行研究,揭示各群丛物种多样性特征及其随相对海拔变化规律,为弄岗北热带季节性雨林的生物多样性保护和喀斯特退化生态系统的植被恢复提供理论参考。
1 材料与方法 1.1 研究区域概况弄岗国家级自然保护区位于我国广西壮族自治区龙州县东边、宁明县北边(106°42′28″—107°04′54″E,22°13′56″—22°33′09″N),保护区呈西北-东南长条状分布,总面积10080hm2。该区为热带季风气候,每年有7个月平均气温达22℃,最冷月平均气温13℃,年最高气温37—39℃,≥10℃的积温为7344—7930℃,无霜期可达351d以上。年平均降水量为1150—1550mm。土壤主要有黑色石灰土和棕色石灰土[16]。
保护区内植物资源极为丰富,有蕨类植物和种子植物172科709属1454种,其中有包括金花茶(Camellia petelotii)、蚬木(Excentrodendron tonkinense)、金丝李(Garcinia paucinervis)等国家珍稀濒危植物33种,广西重点保护野生植物73种,广西特有植物101种,喀斯特(岩溶)特有植物278种。被子植物以大戟科、茜草科、蝶形花科、禾本科、菊科等泛热带性质(或世界广布)科为优势[14, 17]。
该区属于喀斯特“峰丛-洼地”地貌,从洼地到山顶的绝对高度一般只有200—300m,但水热条件差异很大,植被垂直变化很明显。从低海拔往高海拔,包含了3种主要生境类型:(1) 洼地及其边缘,主要分布有喜湿耐阴性强的种类,如海南风吹楠(Horsfieldia hainanensis)、东京桐(Deutzianthus tonkinensis)、中国无忧花(Saraca dives)等;(2)坡地中部,主要分布有蚬木、肥牛树(Cephalomappa sinensis)、闭花木(Cleistanthus sumatranus)等旱生性较强的植物种类;(3)山顶及其周围,主要以细叶谷木(Memecylon scutellatum)、毛叶铁榄(Sinosideroxylon pedunculatum)、米念芭(Tirpitzia ovoidea)为主的山顶矮林[18]。
1.2 样地设置与数据收集弄岗样地位于弄岗保护区弄岗片核心区弄姆皇,地理位置22°25′N、106°57′E,样地东西长500m,南北宽300m。样地包含一个小型山峰和一个较完整的洼地,相对海拔高差只有190m,但生境异质性强,包括了从山顶到洼地完整的“峰丛-洼地”生境类型系列。样地山顶岩石裸露度高,气候干热,洼地伴有季节性水淹,气候阴湿,是桂西南地区“峰丛-洼地”地貌的典型代表。依照CTFS全球森林生物多样性监测规范[19],整个样地用全站仪划分为1, 500个10m×10m的样方(为使后续研究结果能和其他样地进行比较,在进行数据分析时采用CTFS统一的20m×20m)。调查时,用铝牌标记并测量样方内所有胸径≥1cm的木本植物,记录其种名、胸径、坐标及生长状况。样地于2011年底完成第一次调查工作,获得样地第一次植被及地形因子数据(图 1)。

|
| 图 1 弄岗样地的三维地形图。 Fig. 1 Topography of the Nonggang plot |
样地包含了山峰和洼地,地形变异很大,海拔范围180—370m,凹凸度从-5.0m到12.7m,坡度从5.6—68.0°,坡向从0.7—358.1°。样地内胸径≥1cm的木本植物有223种,隶属于56科157属,独立个体总数为68010株,包括分枝、萌枝在内的活体个数为95471株[16]。
1.3 各群丛分布及其特征黄甫昭等[15]根据各样方物种重要值和地形因子,采用多元回归树方法将整个样地划分8个群丛,并对各群丛进行命名,各群丛名称、分布情况及主要特征分别见表 1、图 2、表 2。群丛划分具体方法和步骤详见黄甫昭等[15]文献。
| 代号Codes | 群丛名称Association |
| A | 对叶榕-苹婆-董棕群丛Asso. Ficus hispida-Sterculia monosperma-Caryota obtusa |
| B | 南方紫金牛-苹婆-中国无忧花群丛Asso. Ardisia thyrsiflora-Sterculia monosperma-Saraca dives |
| C | 山石榴-广西牡荆-蚬木群丛Asso. Catunaregam spinosa-Vitex kwangsiensis-Excentrodendron tonkinense |
| D | 垂茉莉-苹婆-广西棋子豆群丛Asso. Clerodendrum wallichii-Sterculia monosperma-Cychidendron guangxiensis |
| E | 米仔兰-闭花木-蚬木群丛Asso. Aglaia odorata-Cleistanthus sumatranus-Excentrodendron tonkinense |
| F | 割舌树-闭花木-蚬木群丛Asso. Walsura robusta-Cleistanthus sumatranus-Excentrodendron tonkinense |
| G | 枝花流苏树-广西牡荆-蚬木群丛Asso.Chionanthus ramiflorus-Vitex kwangsiensis-Excentrodendron tonkinense |
| H | 齿叶黄皮-黄梨木-蚬木群丛Asso.Clausena dunniana-Boniodendron minius-Excentrodendron tonkinense |

|
| 图 2 弄岗样地群丛分布图 Fig. 2 Map of the association in the Nonggang plot |
| 群丛 Ass. |
样方数 Number of plot |
相对海拔/m Relative altitude |
坡度/(°) Slope |
坡向 Aspect |
物种数Number of species | |||
| 群丛整体 Association |
乔木层 Tree layer |
亚乔木层 Subtree layer |
灌木层 Shrub layer |
|||||
| A | 22 | < 12.9 | < 31.5 | 谷底 | 27 | 13 | 7 | 7 |
| B | 23 | 12.9—44.6 | < 31.5 | 半阳坡、阳坡 | 26 | 11 | 8 | 7 |
| C | 68 | 44.6—76.3 | >31.5 | 阳坡 | 32 | 16 | 10 | 6 |
| D | 77 | 44.6—76.3 | >31.5 | 阴坡 | 31 | 15 | 10 | 6 |
| E | 61 | 76.3—135.3 | 31.5—41.2 | 半阴坡 | 29 | 14 | 11 | 4 |
| F | 47 | 76.3—135.3 | >41.2 | 半阳坡 | 30 | 14 | 12 | 4 |
| G | 28 | 76.3—135.3 | < 41.2 | 垭口 | 38 | 19 | 12 | 6 |
| H | 49 | >135.3 | >41.2 | 山顶 | 34 | 11 | 15 | 7 |
根据样地各物种的平均胸径和最大胸径,结合样地调查过程中积累的经验,将所调查物种划归3个层次:乔木层、亚乔木层和灌木层。基于群丛划分结果,分别计算群丛所包含样方整体及样方各层片多样性指数,并分别求算它们的平均值,得到各群丛整体及各层片的平均多样性指数和标准误。根据前人对各种多样性指数的应用和评价[20-22],本文选用以下4个常用的指数:
Patrick指数
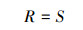 (1)
(1)
Shannon指数
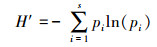 (2)
(2)
Simpson指数
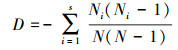 (3)
(3)
Pielou指数
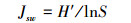 (4)
(4)
式中,S为样方内的物种数;pi为物种i在样方内的相对多数;N为样方内为所有物种的多度之和;Ni为样方内第i个种的多度。
1.4.2 各群丛的差异性比较不同群丛多样性指数差异时,首先检验不同群丛生物多样性指数的正态性及方差齐性,如果多样性指数具有正态性而方差又齐性,则使用单因素方差分析进行检验,并用Duncan的方法进行多重比较;反之则使用Kruskal-Wallis进行检验,并用Wilcoxon Rank Sum Tests进行多重比较。数据分析、统计使用R 3.0、OriginPro 8.6和Excel完成。
2 结果与分析 2.1 群丛多样性及其与相对海拔的关系由各群丛多样性指数变化图(图 3)可以看出,Patrick丰富度指数、Shannon指数和Pielou指数几乎呈一样的倒“W”型变化,处于中坡以下(弄岗样地相对海拔76.3m以下)群丛(群丛A、B、C、D),这3个指数随海拔增高呈“中间高,两头低”即处于该地段中部的群丛的多样性高,相对海拔较低和较高的群丛的多样性低;中坡以上群丛(群丛E、F、G、H)这3个指数随着相对海拔升高而增大。而Simpson指数呈相反的“W”型变化趋势,中坡以下群丛Simpson指数呈“两头高,中间低”;中坡以上群丛Simpson指数随着海拔升高而减小。其中,群丛G、H、C的Patrick指数较高(分别为37.64、34.12、31.78),群丛B和A的最低(分别为26.26、26.64),其余群丛处于中间位置。而图 3表明群丛G、H的Shannon指数、Pielou指数最高,群丛A、E的最低,其余群丛居中。图 3还呈现出相反的规律,Shannon、Pielou指数最低的群丛A、E,Simpson指数最高,Shannon、Pielou指数最高的群丛G、H,Simpson指数最低。

|
| 图 3 弄岗样地各群丛物种丰富度指数、多样性指数和均匀度指数变化 Fig. 3 Variation of richness index, diversity index and evenness index of eight associations in the Nonggang plot |
对各群丛多样性指数进行Kruskal-Wallis检验表明,各群丛的Patrick指数、Shannon指数、Simpson指数和Pielou指数都呈极显著差异(表 3),进一步的Wilcoxon多重比较(表 4)表明,8个群丛的多样性排序为:G > H > C > B > D > F > E > A。呈现出处于喀斯特“峰丛-洼地”垭口的群丛(群丛G)的物种多样性最高,其次为处于山顶的群丛(群丛H),然后是处于坡面中下部的群丛(群丛C、B、D),次末的为坡面中上部的群丛(群丛F、E),最低的是处于谷底的群丛(群丛A)。
| 多样性指数 Diversity indices |
χ2 | P | 多样性指数 Diversity indices |
χ2 | P |
| Patrick指数 Patrick index |
59.761 | 1.68×10-10 | Shannon指数 Shannon index |
83.723 | 2.39×10-15 |
| Simpson指数 Simpson index |
89.636 | 1.47×10-16 | Pielou指数 Pielou index |
77.755 | 3.95×1014 |
| 群丛类型 Associations |
Patrick指数 Patrick index |
Shannon指数 Shannon index |
Simpson指数 Simpson index |
Pielou指数 Pielou index |
| A | 26.6363ABa | 2.2412Aa | 0.7710Aa | 0.6818Aa |
| B | 26.2608Aa | 2.5555ABa | 0.8742Bb | 0.7863ABcd |
| C | 31.7794BCDbc | 2.6094Bb | 0.8711Bb | 0.7587Abc |
| D | 31.2987ACbc | 2.5523ABa | 0.8615Bb | 0.7450Ab |
| E | 29.2166ABab | 2.3296Aa | 0.7851Aa | 0.6934Aa |
| F | 29.9791ABCab | 2.4965ABa | 0.8476ABab | 0.7371Aab |
| G | 37.6428Dd | 2.9318Cc | 0.9099Cc | 0.8116Bd |
| H | 34.1224CDcd | 2.8363Cc | 0.9034Cc | 0.8057Bd |
| 同一列不同大小写字母分别表示差异经过wilcoxom检验达到极显著(P < 0.01)和显著水平(P < 0.05) | ||||
群丛间多样性差异呈现出空间相邻群丛的差异小于空间相离群丛。空间相离的群丛即使多样性差异不明显,但物种组成差异也会很大。如表 1、表 4所示,群丛A、E的各指数都没有显著差异,但各层的优势种却截然不同,群丛A乔木层、亚乔木层的优势种分别为董棕、苹婆,群丛E的分别为蚬木、闭花木;而空间相邻的A、B、D群丛虽然个别指数存在差异,但它们亚乔木层的优势种都为苹婆。
2.2 群丛各层次多样性及其与相对海拔的关系由群丛各层次Patrick和Shannon指数变化(图 4)可以看出,大部分群丛乔木层的Patrick和Shannon指数大于亚乔木层,只有群丛H乔木层Patrick指数低于亚乔木层,群丛E、H的乔木层Shannon指数低于亚乔木层;同时,亚乔木层的Patrick丰富度指数和Shannon指数大于灌木层,只有群丛B的Shannon指数例外。此外,亚乔木层的Patrick和Shannon指数值呈现随相对海拔增高逐渐增高的变化趋势,其他层次的没有明显的变化规律。

|
| 图 4 弄岗样地各群丛乔木层、亚乔木层和灌木层物种多样性指数 Fig. 4 Species diversity indices of the tree, subtree and shrub layer of eight associations in the Nonggang plot |
由群丛各层次Simpson指数变化可以看出,乔木层的Simpson指数低于其他两个层次的,只有群丛E、H的高于亚乔木层。灌木层的Simpson指数普遍高于亚乔木层和乔木层,只有群丛B的低于亚乔木层。同时,亚乔木层的Simpson指数呈现出随相对海拔增高逐渐降低变化趋势。
群丛各层次Pielou指数变化表明,不同层次Pielou指数呈交叉性变化,没有哪个层次的指数明显高于其他层次。乔木层Pielou指数随相对海拔增高呈下降趋势;亚乔木层则相反,随相对海拔增高而增大;灌木层Pielou指数随相对海拔变化没有明显变化,表 5的Kruskal-Wallis检验也表明灌木层Pielou指数在各群丛之间没有显著差异(χ2=4.54,P=0.716)。
| 群丛层次 Layer of association |
Patrick指数 | Shannon指数 | Simpson指数 | Pielou指数 | ||||
| χ2 | P | χ2 | P | χ2 | P | χ2 | P | |
| 乔木层 Tree layer |
82.26 | 4.77×10-15 | 103.03 | 2.55×10-19 | 82.26 | 4.77×10-15 | 127.55 | 2.04×10-24 |
| 亚乔木层 Subtree layer |
130.59 | 4.72×10-25 | 144.18 | 6.76×10-28 | 130.59 | 4.72×10-25 | 102.08 | 4.02×10-19 |
| 灌木层 Shrub layer |
94.69 | 1.34×10-17 | 72.99 | 3.66×10-13 | 94.69 | 1.34×10-17 | 4.54 | 7.16×10-1 |
相对海拔是影响喀斯特山地水热条件分布的主要因素,它通过影响光照、水分及土壤资源的再分配进而影响植物群丛的分布及结构[1]。弄岗样地各类群丛占据样地不同的地形部位,随着相对海拔变化呈明显规律性分布,群丛分异主要源于相对海拔变化[15]。群丛所处相对海拔不同,水分、热量、光照及土壤等条件都有所差别,从而使群丛的物种组成和多样性也有所不同。
不同群丛多样性指数分析表明,弄岗喀斯特季节性雨林各群丛的多样性指数存在极显著差异,Patrick、Shannon和Pielou指数随着相对海拔升高呈两段式变化,中坡以下,多样性随相对海拔增高呈“中间高两头低”变化规律,中坡以上则随相对海拔增高而增大,Simpson指数呈相反变化趋势。这是因为Simpson指数表征的是多样性的集中性,集中性越高,说明群丛中少数物种占据绝对优势,从而会排斥其他物种的生存空间,进而使整个群落的物种丰富度和多样性下降,所以Simpson指数和其他3个指数呈相反变化规律。
先前的研究表明物种多样性随海拔变化有5种模式[23],即1)物种多样性与海拔高度负相关,2)物种多样性中海拔最高,3)物种多样性中海拔较低,4)物种多样性与海拔高度正相关,5)物种多样性与海拔高度无关。弄岗喀斯特季节性雨林物种多样性随海拔变化规律与这几种模式都不一样,或者说不完全一样,呈现出两种模式的组合,中坡以下(弄岗样地相对海拔76.3 m以下)与第2种模式一致,中坡以上(弄岗样地相对海拔大于76.3 m)与第1种模式一致。这是由于相对海拔较低的地带,水分充足甚至过量(谷底会有季节性水淹),但光照条件较差,随着相对海拔增高光照条件逐渐改善,物种丰富度和多样性指数也逐渐增加,但上升到一定程度高度之后,水分条件变差(喀斯特地区土层稀少,加上特殊的双层结构,水分会迅速从下层流失,造成干旱),物种丰富度和多样性指数又逐渐下降。中坡以上地带,随着海拔升高坡度逐渐增大,气温剧烈变化的石崖也越来越多,产生了许多异质性的微生境,加上生境条件严酷,没有那个物种在相对海拔较高的群丛形成绝对优势,从而支持更多物种在该地带共存。
同时,样地多样性最高的也并非处于山顶的群丛H而是处于次高位置的群丛G,这是因为群丛G处于“峰丛-洼地”垭口部位,土层相对连续,水分条件优越,同时垭口相对海拔较高,且呈东西走向,光照条件较好,水热组合最佳,资源可用性最高,因此,群丛G的物种丰富度和多样性指数是整个样地最高的。由此可以看出,弄岗喀斯特季节性雨林不同群丛物种多样性变化主要是由于能量差异和空间异质性。
通过对群丛不同层次多样性进行研究发现,弄岗北热带喀斯特季节性林各层次多样性呈现出乔木层>亚乔木层>灌木层,这与张忠华[24]研究木论亚热带喀斯特常绿阔叶林得到的灌木层>亚乔木层>乔木层结果截然相反,与李宗善[25]对西双版纳热带山地雨林的研究结果一致。这是因为热带地区水热条件要比亚热带优越,可以支持乔木层有更多的物种共存[26],致使北热带喀斯特带季节性雨林乔木层的物种多样性大于其他层次。同时由于乔木层物种多样性较高,对亚乔木形成比较大的荫蔽作用,致使亚乔木层的物种多样性较小,而灌木层由于乔木层和亚乔木层的双重遮蔽,只有一些比较耐阴的种类才能生长,所以它的物种多样性最低。群丛H的结构特征也恰好验证了这种推论,群丛H乔木层Patrick和Shannon指数是所有群丛中最低的,且低于亚乔木层,这是因为群丛H处于山顶,是所有群丛中条件最恶劣的,这种恶劣条件难以支持乔木层大量物种共存,所以乔木层物种多样性较低;而没有被乔木层大量遮蔽的亚乔木层得到比较好的发展,物种多样性较高;灌木层由于受到乔木层和亚乔木层的双重遮蔽,多样性依然最低。因此,在喀斯特植被退化区,特别是那些环境条件特别恶劣的区域进行植被恢复时,要谨慎选择大乔木,应更多选择喜光、耐旱的灌木、小乔木。
致谢: 感谢加拿大湖首大学生物系Azim U. Mallik教授和广西植物研究所姚月锋博士对写作的帮助。| [1] | 袁道先, 章程. 岩溶动力学的理论探索与实践[J]. 地球学报 , 2008, 29 (3) : 355–365. |
| [2] | 袁铁象, 张合平, 欧芷阳, 谭一波. 地形对桂西南喀斯特山地森林地表植物多样性及分布格局的影响[J]. 应用生态学报 , 2014, 25 (10) : 2803–2810. |
| [3] | 蒋国芳. 木论喀斯特森林区生物多样性初报[J]. 生物多样性 , 1995, 3 (2) : 91–94. |
| [4] | 宋同清, 彭晚霞, 曾馥平, 王克林, 曹洪麟, 李先琨, 覃文更, 谭卫宁, 刘璐. 喀斯特峰丛洼地不同类型森林群落的组成与生物多样性特征[J]. 生物多样性 , 2010, 18 (4) : 355–364. |
| [5] | 侯满福, 沈庆庚, 覃海宁. 贵州茂兰喀斯特原生性森林群落物种多样性特征[J]. 广西师范大学学报:自然科学版 , 2011, 29 (1) : 60–65. |
| [6] | 张忠华, 胡刚, 祝介东, 倪健. 喀斯特常绿落叶阔叶混交林物种多度与丰富度空间分布的尺度效应[J]. 生态学报 , 2012, 32 (18) : 5663–5672. |
| [7] | Polak Marcel. The botanical diversity in the Ayawasi area, Irian Jaya, Indonesia[J]. Biodiversity and Conservation , 2000, 9 (10) : 1345–1375. DOI:10.1023/A:1008974730159 |
| [8] | Brewer Steven W, Rejmánek Marcel, Webb Molly AH, Fine Paul VA. Relationships of Phytogeography and diversity of tropical tree species with limestone topography in southern Belize[J]. Journal of Biogeography , 2003, 30 (11) : 1669–1688. DOI:10.1046/j.1365-2699.2003.00971.x |
| [9] | Jeanine M Felfili, André R Terra Nascimento, Christopher William Fagg, Elisa Maria Meirelles. Floristic composition and community structure of a seasonally deciduous forest on limestone outcrops in Central Brazil[J]. Revista Brasileira de Botânica , 2007, 30 (4) : 611–621. |
| [10] | 李先琨, 苏宗明, 吕仕洪, 欧祖兰, 向悟生, 区智, 陆树华. 广西岩溶植被自然分布规律及对岩溶生态恢复重建的意义[J]. 山地学报 , 2003, 21 (2) : 129–139. |
| [11] | 苏宗明, 赵天林, 黄庆昌. 弄岗自然保护区植被调查报告[J]. 广西植物 , 1988 (增刊) : 185–214. |
| [12] | 王献溥, 孙世洲, 李信贤. 广西石灰岩季节性雨林分类的研究[J]. 植物研究 , 1998, 18 (4) : 428–447. |
| [13] | 胡春园, 汪正祥, 雷耘. 弄岗自然保护区石灰岩山地季雨林的群落分类及分布[J]. 华中师范大学学报(自然科学版) , 2012, 46 (6) : 725–732. |
| [14] | 黄俞淞, 吴望辉, 蒋日红, 刘晟源, 刘演, 李先琨. 广西弄岗国家级自然保护区植物物种多样性初步研究[J]. 广西植物 , 2013, 33 (3) : 346–355. |
| [15] | 黄甫昭, 王斌, 丁涛, 向悟生, 李先琨, 周爱萍. 弄岗北热带喀斯特季节性雨林群丛数量分类及与环境的关系[J]. 生物多样性 , 2014, 22 (2) : 157–166. |
| [16] | 王斌, 黄俞淞, 李先琨, 向悟生, 丁涛, 黄甫昭, 陆树华, 韩文衡, 文淑均, 何兰军. 弄岗北热带喀斯特季节性雨林15ha监测样地的树种组成与空间分布[J]. 生物多样性 , 2014, 22 (2) : 141–156. |
| [17] | 梁畴芬, 梁建英, 刘兰芳, 莫新礼. 弄岗自然保护区植物区系调查报告[J]. 广西植物 , 1988 (增刊) : 83–184. |
| [18] | 苏宗明. 广西弄岗石灰岩山森林类型的分类问题[J]. 广西植物 , 1981, 1 (2) : 7–10. |
| [19] | Condit R. Research in large, long-term tropical forest plots[J]. Trends in Ecology and Evolution , 1995, 10 (1) : 18–23. DOI:10.1016/S0169-5347(00)88955-7 |
| [20] | 马克平, 刘玉明. 生物群落多样性的测度方法Ⅰα多样性的测度方法(下)[J]. 生物多样性 , 1994, 2 (4) : 231–239. |
| [21] | 邱扬, 李湛东, 张玉钧, 徐化成, 于汝元. 火干扰对大兴安岭北部原始林下层植物多样性的影响[J]. 生态学报 , 2006, 26 (9) : 2863–2869. |
| [22] | Bandeira Benjamin, Jamet Jean Louis, Jamet Dominque, Ginoux Jean Marc. Mathematical convergences of biodiversity indices[J]. Ecological Indicators , 2013, 29 (6) : 522–528. |
| [23] | 贺金生, 陈伟烈. 陆地植物群落物种多样性的梯度变化特征[J]. 生态学报 , 1997, 17 (1) : 91–99. |
| [24] | 张忠华.喀斯特森林植被种群生态学与群落稳定性分析[D].上海:华东师范大学, 2011. |
| [25] | 李宗善, 唐建维, 郑征, 李庆军, 罗成昆, 刘正安, 李自能, 段文勇, 郭贤明. 西双版纳热带山地雨林的植物多样性研究[J]. 植物生态学报 , 2004, 28 (6) : 833–843. |
| [26] | 黄建辉, 高贤明, 马克平, 陈灵芝. 地带性森林群落物种多样性的比较研究[J]. 生态学报 , 1997, 17 (6) : 611–618. |