 2016, Vol. 36
2016, Vol. 36文章信息
- 吴青怡, 曹振东, 付世建
- WU Qingyi, CAO Zhendong, FU Shijian.
- 鳊鱼和宽鳍鱲幼鱼流速选择与运动能量代谢特征的关联
- Flow velocity selection and its relationship to locomotive energetic metabolism in Chinese bream (Parabramis pekinensis) and pale chub (Zacco platypus)
- 生态学报[J]. 2016, 36(13): 4187-4194
- Acta Ecologica Sinica[J]. 2016, 36(13): 4187-4194
- http://dx.doi.org/10.5846/stxb201411162272
-
文章历史
- 收稿日期: 2014-11-16
- 网络出版日期: 2015-10-29

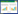




鱼类作为脊椎动物的最大类群,不仅种类繁多、生活史复杂,而且栖息生境多变且差异明显。对于绝大多数鱼类而言,游泳运动与其生存、生长和繁衍等生命活动密切相关[1-3],且对鱼类逃避敌害、求偶展示、洄游以及探寻最佳栖息地均具有重要意义[3-4]。在相关研究中,鱼类的稳定游泳运动能力受到研究者的广泛关注,其中,临界游泳速度(即最大持续有氧游泳速度)(Ucrit)及最大代谢率(MMR)常作为衡量鱼类稳定游泳能力的重要指标[5-6]。野外实验研究发现,鱼类对栖息地的水流速度具有偏好性,即鱼类存在偏好水流速度[7-8],这种偏好可能与鱼类的游泳能力密切关联;并且这类研究还涉及诸如温度、食物、捕食者等其他因子影响鱼类对栖息地的选择[7-8]。另外,有研究者在不同水流环境的实验条件下独立考察鱼类游泳行为,将停留时间最长的水流速度作为实验鱼的偏好游泳速度(Upref)[9-10]。
尽管鱼类对生境中水流速度的选择受诸多因子影响,但毫无疑问游泳过程的能量利用效率是影响这种选择的的重要的原因之一。鱼类的游泳效率是指单位体重的鱼类移动单位距离所消耗的能量,以单位位移耗能(COT)表示[10]。由于静止代谢(RMR)[10-12]的存在,COT曲线随着游泳速度的增加呈现先下降后上升的趋势,因此曲线存在一个最低点。该点单位位移耗能最少,能量效率最高,其所对应的游泳速度被称为最适游泳速度(Uopt)[13-15]。
Upref是鱼类在梯度水流速度环境中的行为反应,而COT曲线导出的Uopt则集中反映鱼类的运动代谢特征。有研究者对二者的关系进行了探讨,发现溪红点鲑(Salvelinus fontinalis)存在一个Uperf范围,且存在着与其Uopt相重叠的现象。因此研究者认为溪红点鲑的Uopt能够反映其Uperf [9],其Uperf的选择与运动能量效率密切关联。然而,鱼类对Uperf的选择更多是由于生存胁迫(天敌等)和生长、繁衍需求(觅食求偶)所致[16-17]。仅以鱼类的能量消耗进行分析,不难发现静止状态是最节约能量的。据此推论:当生存胁迫和需求发生变化时,鱼类的Uperf和Uopt有可能出现分离的现象。因此本研究的第1个目标是研究两种淡水鲤科鱼类的Uperf和Uopt是否存在关联。
已有研究发现激流生境与缓流生境生存的鱼类在形态上存在差异,前者更趋于流线体型[18-19];而且激流生境鱼类的Uperf通常高于生活于缓流环境的鱼类[19];研究还发现这两种类型鱼类的运动代谢特征也存在差异[19]。那么,这两种不同生境生活的鱼类Uperf的不同是否受运动能量代谢特征的影响是一个值得关注的问题。据此本研究的第2个目标是考察生存于不同水流环境两种鱼类的Uperf与运动能量代谢特征之间的关联。
为实现上述目标,本研究分别选取喜好缓流生境的鳊鱼(Parabramis pekinensis)和喜好激流生境的宽鳍鱲(Zacco platypus)作为实验对象[19-20] (图 1)。首先测定鳊鱼和宽鳍鱲的Ucrit、不同游泳速度下的MO2,以此计算COT并导出最适游泳速度Uopt;然后利用流速梯度选择仪获取实验鱼的流速选择行为摄像资料,进而得到两种鱼的流速度选择特征。最终通过分析,揭示两种实验鱼的速度选择特征与运动能量代谢特征的关联。

|
| 图 1 宽鳍鱲和鳊鱼 Fig. 1 Pale chub (Zacco platypus) and Chinese bream (Parabramis pekinensis) in the present study |
在2013年8月在乌江支流捕获本研究所用的宽鳍鱲幼鱼(生境水流速度范围1.5—4 m/s)[19],鳊鱼幼鱼在2013年10月购于四川省水产学校(生境为静水),将它们置于本实验室规格为1.2 m × 0.55 m × 0.55 m 的自净化循环控温水槽内驯养15 d。驯化期间,每天以通威商业颗粒饲料饱足投喂1次,投喂1 h后立即清除粪便和残饵,日换水量约为驯化水体的10%。水温控制在(25 ± 1.0) ℃,用充气泵不断向水体充入空气,水体溶氧水平大于90%饱和溶氧水平,光周期为12L∶12D。驯化结束后,挑选鱼体健康的鳊鱼[体重(6.38±0.76) g,体长(6.90 ± 0.45) cm]和宽鳍鱲[体重(3.09 ± 0.57) g,体长(5.96 ± 0.28) cm]各21尾作为实验对象。
1.2 实验方案实验分为两个部分。
第一部分随机挑选出鳊鱼和宽鳍鱲幼鱼各8尾,在水温(25 ± 1) ℃下测定各自在不同游泳速度下(直至最大游泳速度)的MO2,据此计算两种鱼不同速度下的COT、Ucrit 、Uopt和RMR。
第二部分根据两种鱼的Ucrit和Uopt水流速度范围设置为2.5—15倍体长(BL),分别挑选出鳊鱼和宽鳍鱲幼鱼13尾,在上述相同水温条件下进行游泳行为的视频拍摄,随后对视频资料进行分析获取两种鱼的偏好游泳速度。
1.3 测定方法及主要参数 1.3.1 Ucrit的测定本实验测定设备采用自行设计的密闭式鱼类游泳代谢测定仪(专利号: 200920127986.3和200920127988.2) ,其结构与工作原理在相关文献[21-22] 已有报道。首先将游泳代谢仪浸没于恒温(25 ± 0.5) ℃水槽中,然后将实验鱼放入流速为7.0 cm/s(约为实验鱼的1 倍BL)游泳管中适应1 h,以消除转移胁迫的影响;随后采用逐步提速法进行Ucrit测定。测定以7 cm/s为起始速度,以速度增量(ΔV)为7.0 cm/s,持续时间(ΔT)为20 min,游泳速度不断增加直至实验鱼达到运动力竭状态。力竭状态的评判标准为实验鱼停靠在游泳管末端筛板20 s 以上[21-22]。测定过程中每20 min 换水1次,以确保密闭游泳管内的水体溶氧水平不低于饱和状态的70%。使用溶氧仪(HQ30,Hach Company,Ioveland,Colorado,USA)进行监测溶氧水平。最后测量实验鱼的体重与体长。Ucrit的计算公式如下:
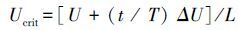
|
(1) |
式中,Ucrit 为临界游泳速度(BL/s),U 为能够完成设定时间(20 min)的最大游泳速度(cm/s),ΔU为速度增量(7.0 cm/s),T为设定的持续游泳历时(20 min),t为未能完成设定历时的实际持续游泳时间(t<20 min),L为实验鱼体长(cm)以减少体长对游泳速度的影响。另外,因本实验鱼的鱼体横截面积未超过游泳管截面积的10%,所以Ucrit无需校正[23]。
1.3.2 MO2的测定在测定Ucrit 的过程中,每2 min 测定1次游泳代谢仪内封闭水体的溶氧值,以水体的体积和溶氧值随时间变化斜率的绝对值计算出每尾实验鱼的运动过程MO2。实验开始前和在测定细菌好氧实验结束将实验鱼取出后,均在水流速度为35 cm/s时对游泳代谢仪重新密闭并进行20 min的耗氧率测定,以扣除细菌耗氧的影响[23]。MO2的计算公式如下:
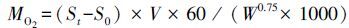
|
(2) |
式中,MO2代表运动耗氧率(mg kg-1 h-1),St为每档期间(20min)溶氧值随时间变化的斜率(mg O2/min),S0 为空白对照溶氧值的变化斜率,V为运动代谢仪的系统体积(3 L),60为60 min。W表示实验鱼的体重(g)。由于两种实验鱼的体重差异显著,因此采用 W0.75进行MO2校正[24-25]。在Ucrit测定过程中最大的MO2值作为MMR。
1.3.3 COT、RMR及Uopt的计算单位位移耗能(COT)
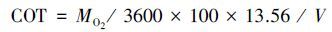
|
(3) |
式中,COT 为单位移动距离能耗(J kg-1 m-1),MO2为实验鱼非体重指数校正的耗氧率(mg O2 kg-1 h-1),V 则表示实验鱼游泳速度(m/h),13.56为氧热当量系数(J /mg O2)[26]。
为直接的表达游泳速度和能耗的关系,本研究还计算了单位位移净耗能(COT net):
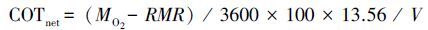
|
(4) |
式中,V为游泳速度(cm/s)。
由于MO2的计算方式可以表示为:
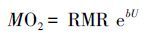
|
(5) |
则COT计算公式为:
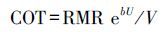
|
(6) |
式中,RMR为静止代谢率(mg kg-1 h-1),b为常数。
根据COT公式求导,当COT′=0 时,可得出Uopt 的计算公式如下[27]:
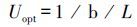
|
(7) |
式中,Uopt为最适游泳速度(BL/s),L为实验鱼的体长(cm)。
1.3.4 速度选择分析流速选择在实验室自行研制的梯度流速选择仪(图 2)中测定。梯度流速选择仪一端为流速可控的水泵(图 2D),与水泵相连的是导流筛板(图 2E),其后与直径呈线性增大的圆锥形流速选择泳道相连(图 2C),流速选择泳道的末端设有防逃逸的多孔防逸筛板(图 2H),流速选择仪置于恒温水槽中(图 2F),摄像机(图 2B)置于流速选择仪的正上方。实验时首先将实验鱼转入流速选择泳道中适应1 h[10],以消除转移胁迫的影响。适应完毕开始摄像3次,每次摄像时间为2 h,每隔4 h重复摄像一次,共获得到6 h的摄像资料。为验证Uperf和Uopt是否重叠,本研究水流速度设置范围为18.6—102.7 cm/s,此流速范围的划分根据前期实验工作和本次采样调查而设定。梯度流速仪被等距离划分为5个区域,其流速范围见图 3。然后通过软件Ethvision XT9分析得出实验鱼在每个速度区域的停留时间以及出入频次。

|
| 图 2 鱼类流速选择仪结构示意图 Fig. 2 The structure of fish preferred swimming speed determining device |

|
| 图 3 鳊鱼和宽鳍鱲在各速度下的停留时间百分比(Pt)和出入频率百分比(Pf)(平均值 ± 标准误,n=13) Fig. 3 The Pt and Pf of Chinese bream and pale chub (Mean ± SE,n=13) |
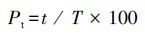
|
(8) |
式中,Pt为实验鱼在速度区域的停留时间百分比(%),t为鱼在速度区域的停留时间(s),T为每条鱼的录像时间(s)。
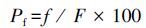
|
(9) |
式中,Pt为实验鱼在速度区域出入的频率百分比(%),t为鱼在速度区域出入次数,T为每条鱼的录像期间在所有区域出入的总次数。
1.4 数据统计实验数据以Excel 2007进行常规计算,接着采用SPSS 17.0进行数据分析。其中游泳速度和鱼类种类对MO2和COT的影响采用可重复测量的双因素协方差分析(体重为协变量);鱼种间Ucrit、COT、MMR和Uopt的差异采用t检验分析;不同鱼种在各速度区域Pt和Pf采用双因素方差分析。统计值均以平均值±标准误(Mean ± SE)表示,显著水平为 P<0.05。
2 结果 2.1 鳊鱼和宽鳍鱲的Pt 和Pf分析实验鱼在流速选择仪中行为的摄像资料,结果显示:鳊鱼和宽鳍鱲在(18.6—23.8 cm/s)区域Pt和Pf均显著大于其他流速区域(所有P<0.05) ,且随着水流速度的增加,鳊鱼和宽鳍鱲的Pt和Pf均逐渐减少。经双因素方差分析表明体长体重经对Pt和Pf无影响;水流速度和种类均对Pt和Pf有显著影响(P<0.001,F=9.688,df=4) ;鳊鱼在低流速区域的Pt和Pf显著大于宽鳍鱲(P<0.05) 。鳊鱼在大于23.8 cm/s流速区域的Pt和Pf在总体上有小于宽鳍鱲的趋势,且在大于43.8 cm/s的水流速度区域鳊鱼的Pt和Pf均显著小于宽鳍鱲(P<0.05) 。
2.2 鳊鱼和宽鳍鱲的运动MO2曲线、Ucrit和MMR随游泳速度的增加,两种鱼的MO2均显著上升(P<0.05) (图 4),但鳊鱼的MO2曲线上升速率更快(交互作用:P<0.001,F=17.459,df=1) ;在低于21(cm/s)游泳速度时鳊鱼的MO2显著低于宽鳍鱲,而在高于63cm/s游泳速度时前者的MO2显著高于后者。

|
| 图 4 鳊鱼和宽鳍鱲在不同游泳速度下的耗氧率曲线、最大有氧代谢率和临界游泳速度 (平均值±标准误,n=8) Fig. 4 The oxygen consumption at different swimming speeds,maximum metabolic rate and critical swimming speed of Chinese bream and pale chub (Mean±S E,n=8) |
鳊鱼的Ucrit显著低于宽鳍鱲,但二者的MMR没有显著差异(P<0.05) (图 4)。
2.3 鳊鱼和宽鳍鱲的COT曲线、RMR和Uopt随游泳速度的增加,两种鱼的COT均显著下降(P<0.05) ,在高游泳速度下曲线下降趋势逐渐减弱直至趋于平稳(图 5);其中宽鳍鱲的下降速度更快(交互作用P<0.05) ,在低于21cm/s游泳速度时鳊鱼的COT低于宽鳍鱲,而在高于63cm/s游泳速度时前者高于后者。值得注意的是:两种实验鱼的COTnet随着游泳速度增加差异逐渐增大,前者显著高于后者(P<0.05) 。

|
| 图 5 两种实验鱼的单位距离耗能、静止代谢、和最适速度(平均值±标准误,n=8) Fig. 5 The cost of transport curves,resting metabolic rate and optimal swimming speed of Chinese bream and pale chub (Mean±SE,n=8) |
鳊鱼幼鱼的RMR和Uopt均显著小于宽鳍鱲(P<0.001) (图 5)。
3 讨论 3.1 实验鱼流速选择与最适速度的关联研究显示,溪红点鲑的Uperf在0.78—0.95 BL/s之间[7],Uopt为1.02 BL/s,其Uopt与其Uperf几乎完全重叠,因此可以将溪红点鲑的Uopt用以反映其Uperf [9]。本研究结果显示,鳊鱼和宽鳍鱲的Uopt 分别为6.2和11.56BL /s;而鳊鱼和宽鳍鱲在最小速度区域(18.6—21.3 cm/s) 的Pt和Pf均显著大于其它区域,表明鳊鱼和宽鳍鱲的Uperf均等于或者小于本实验设定的最小速度范围(2.5—3.5 BL/s),这远远小于两种实验鱼的Uopt,这与溪红点鲑的研究结果存在差异。通过上述结果分析,两种实验鱼的Uopt与Uperf存在着明显的分离现象。
研究发现杜父鱼(Cottus bairdi)和长鼻鲦鱼(Rhinichthys cataractae)由于形态上的适应(体型呈圆筒流线型),在选择栖息地时不完全受能量限制,而虹鳟(Oncorhynchus mykiss)和鲦鱼(Clinostomus funduloides)存在流速偏好且其偏好受季节影响[27-28];许氏鹦哥鱼(Scarus schlegeli)和叉斑锉鳞鲀(Rhinecanthus aculeatus)寻食时呈现较稳定的巡游速度[29],有关银大马哈鱼(Oncorhynchus keta)和虹鳟的研究发现其捕食时的巡游速度会显著影响其捕食成功率[30]。而伏击取食的南方鲇(Silurus meridionalis)和红点鲑(Salvelinus leucomaenis)则在非捕食的大部分时间里几乎静止不动[31-32]。因此鱼类的Uperf会因为季节、食物丰度、生活史等方面的影响而表现出很强烈的环境依赖性,甚至还可能由于形态适应的特征使少数种类不存在稳定的Uperf。本研究的两种实验鱼的Uperf和Uopt出现分离的现象,可能是由于实验鱼在没有捕食压力的情况下所表现出的一种节约能量的对策所致,也可能是由于实验水体中食物缺乏所致。
3.2 鳊鱼和宽鳍鱲的速度选择及其能量代谢特征生存于不同流水环境的鱼类,它们的游泳能力存在明显差异且所选择的水流环境也不尽相同[18-19],这种差异产生的机制可能与外部形态、内部结构以及选择压力有关[27-28],还可能与其能量代谢特征相关。本研究结果显示,尽管鳊鱼和宽鳍鱲在最低速度区域18.6—23.8(cm/s)的Pt和Pf均较高,然而鳊鱼却显著高于宽鳍鱲,进而导致鳊鱼在速度更高的其它流速区域Pt和Pf均明显小于宽鳍鱲。由此可见,与鳊鱼相比在激流水体生存的宽鳍鱲更偏好较高的水流速度。另有研究发现,在高水流速度环境中生存的鱼类游泳运动能力更强,且身体形状更趋于流线型,在静水环境生活的鱼类游泳效率更低[18-19]。本研究结果显示,随着游泳速度的增大,鳊鱼的COTnet显著高于宽鳍鱲,即在较快的游泳速度下鳊鱼的运动能量效率更低。根据两种实验鱼的水流速度选择特征可以看出鱼类的运动能量效率与水流速度选择特征可能存在某种联系,即游泳能量效率越高的实验鱼其所选择水流速度也越高,反之亦然。据此本研究结果表明,生存于不同水流生境鱼类的水流速度选择特征存在明显差异,在激流环境中生存的鱼类更加偏好高的水流速度,这可能与其在较高游泳速度范围内具有较高的能量效率有关。
鱼类的游泳运动能力及其能量代谢的研究有助于揭示自然演化的方向和生理功能的适应特征,而鱼类水流速度选择的偏好及其特征分析则有利于理解环境对鱼类运动强度和游泳方式差异的影响,因此开展有关生物和非生物环境对鱼类的水流偏好影响的研究将可能成为有价值的研究方向。
| [1] | Katz S L, Syme D A, Shadwick R E.High-speed swimming:Enhanced power in yellowfin tuna.Nature,2001,410(6830): 770–771. |
| [2] | Handelsman C, Claireaux G, Nelson J A.Swimming ability and ecological performance of cultured and wild European sea bass (Dicentrarchus labrax) in coastal tidal ponds.Physiological and Biochemical Zoology,2010,83(3): 435–445. |
| [3] | Langerhans R B, Reznick D N. Ecology and evolution of swimming performance in fishes:predicting evolution with biomechanic//Domenici P, Kapoor B G, eds. Fish Locomotion:An Etho-ecological Perspective. British Isles:Science Publishers, 2010:200-248. |
| [4] | Domenici P. Escape responses in fish:kinematics, performance and behavior//Domenici P, Kapoor B G, eds. Fish Locomotion:An Etho-ecological Perspective. British Isles:Science Publishers, 2010:123-170. |
| [5] | Fry F E J, Hart J S.The relation of temperature to oxygen consumption in the goldfish.The Biological Bulletin,1948,94(1): 66–77. |
| [6] | Brett J R, Groves T D D. Physiological energetics//Hoar W S, Randall D J, Brett J R, eds. Fish Physiology. New York:Academic Press, 1979:279-352. |
| [7] | Chun S N, Cocherell S A, Cocherell D E, Miranda J B, Jones G J, Graham J, Klimley A P, Thompson L C, Cech J J Jr.Displacement velocity preference, and substrate use of three native California stream fishes in simulated pulsed flows.Environmental Biology of Fishes,2011,90(1): 43–52. |
| [8] | Holm C F, Armstrong J D, Gilvear D J.Investigating a major assumption of predictive in stream habitat models:is water velocity preference of juvenile Atlantic salmon independent of discharge?.Journal of Fish Biology,2001,59(6): 1653–1666. |
| [9] | Tudorache C, O'Keefe R A, Benfey T J.Optimal swimming speeds reflect preferred swimming speeds of brook charr (Salvelinus fontinalis Mitchill, 1874).Fish Physiology and Biochemistry,2011,37(2): 307–315. |
| [10] | Tudorache C, de Boeck G, Claireaux G. Forced and preferred swimming speeds of fish:a methodological approach//Palstra Arjan P, Planas Josep V, eds. Swimming Physiology of Fish:Toward Using Exercise to Farm a Fit Fish in Sustainable Aquaculture. New York:Springer, 2013:81-108. |
| [11] | Hinch S G, Rand P S.Optimal swimming speeds and forward-assisted propulsion:energy-conserving behaviours of upriver-migrating adult salmon.Canadian Journal of Fisheries and Aquatic Sciences,2000,57(12): 2470–2478. |
| [12] | 付成, 曹振东, 付世建.温度和饥饿对鲤鱼幼鱼静止代谢率及自发运动的影响.动物学杂志,2012,47(2): 85–90. |
| [13] | Ware D M.Growth, metabolism, and optimal swimming speed of pelagic fish.Journal of the Fisheries Research Board of Canada,1975,32(1): 33–41. |
| [14] | Videler J J. Fish Swimming. New York:Chapman and Hall, 1993. |
| [15] | Claireaux G, Couturier C, Groison A L.Effect of temperature on maximum swimming speed and cost of transport in juvenile European sea bass (Dicentrarchus labrax).The Journal of Experimental Biology,2006,209(17): 3420–3428. |
| [16] | Chan M D, Dibble E D, Kilgore K J.A laboratory examination of water velocity and substrate preference by age-0 gulf sturgeons.Transactions of the American Fisheries Society,1997,126(2): 330–333. |
| [17] | Van Liefferinge C, Seeuws P, Meire P, Verheyen R F.Microhabitat use and preferences of the endangered Cottus gobio in the River Voer, Belgium.Journal of Fish Biology,2005,67(4): 897–909. |
| [18] | Fu S J, Cao Z D, Yan G J, Fu C, Pang X.Integrating environmental variation, predation pressure, phenotypic plasticity and locomotor performance.Oecologia,2013,173(2): 343–354. |
| [19] | Yan G J, He X K, Cao Z D, Fu S J.An interspecific comparison between morphology and swimming performance in cyprinids.Journal of Evolutionary Biology,2013,26(8): 1802–1815. |
| [20] | 丁瑞华. 四川鱼类志. 成都: 四川科学技术出版社, 1994237–238. |
| [21] | Lee C G, Farrell A P, Lotto A, Hinch S G, Healey M C.Excess post-exercise oxygen consumption in adult sockeye (Oncorhynchus nerka) and coho (O. kisutch) salmon following critical speed swimming.The Journal of Experimental Biology,2003,206(18): 3253–3260. |
| [22] | 杨晗, 曹振东, 付世建.尾鳍缺失对鳊鱼幼鱼游泳能力、能量效率与行为的影响.水生生物学报,2013,37(1): 157–163. |
| [23] | Alsop D H, Wood C M.The interactive effects of feeding and exercise on oxygen consumption, swimming performance and protein usage in juvenile rainbow trout (Oncorhynchus mykiss).The Journal of Experimental Biology,1997,200(Pt 17): 2337–2346. |
| [24] | Reidy S P, Kerr S R, Nelson J A.Aerobic and anaerobic swimming performance of individual Atlantic cod.The Journal of Experimental Biology,2000,203(2): 347–357. |
| [25] | Fu S J, Xie X J, Cao Z D.Effect of meal size on postprandial Metabolic response in southern catfish (Silurus meridionalis).Comparative Biochemistry and Physiology Part A:Molecular & Integrative Physiology,2005,140(4): 445–451. |
| [26] | Randall D.The control of respiration and circulation in fish during exercise and hypoxia.The Journal of Experimental Biology,1982,100(1): 275–288. |
| [27] | Facey D E, Grossman Gary D.The metabolic cost of maintaining position four North American stream fishes:effects of season and velocity.Physiological Zoology,1990,63(4): 757–776. |
| [28] | Facey D E, Grossman G D.The relationship between water velocity, energetic costs, and microhabitat use in four North American stream fishes.Hydrobiologia,1992,239(1): 1–6. |
| [29] | Korsmeyer K E, Steffensen J F, Herskin J.Energetics of median and paired fin swimming, body and caudal fin swimming, and gait transition in parrotfish (Scarus schlegeli) and triggerfish (Rhinecanthus aculeatus).The Journal of Experimental Biology,2002,205(9): 1253–1263. |
| [30] | Piccolo J J, Hughes N F, Bryant M D.Water velocity influences prey detection and capture by drift-feeding juvenile coho salmon (Oncorhynchus kisutch) and steelhead (Oncorhynchus mykiss irideus).Canadian Journal of Fisheries and Aquatic Sciences,2008,65(2): 266–275. |
| [31] | Fu S J, Xie X J.Nutritional homeostasis in carnivorous southern catfish (Silurus meridionalis):is there a mechanism for increased energy expenditure during carbohydrate overfeeding?.Comparative Biochemistry and Physiology Part A:Molecular & Integrative Physiology,2004,139(3): 359–363. |
| [32] | Metcalfe N B, Valdimarsson S K, Fraser N H C.Habitat profitability and choice in a sit-and-wait predator:juvenile salmon prefers lower currents on darker nights.Journal of Animal Ecology,1997,66(6): 866–875. |