 2016, Vol. 36
2016, Vol. 36文章信息
- 袁晓波, 牛得草, 吴淑娟, 蒲向东, 王龙, 滕家明, 傅华
- YUAN Xiaobo, NIU Decao, WU Shujuan, PU Xiangdong, WANG Long, TENG Jiaming, FU Hua.
- 黄土高原典型草原地上生物量估测模型
- Estimation of aboveground biomass in the grassland of the Loess Plateau, Northern China
- 生态学报[J]. 2016, 36(13): 4081-4090
- Acta Ecologica Sinica[J]. 2016, 36(13): 4081-4090
- http://dx.doi.org/10.5846/stxb201411062199
-
文章历史
- 收稿日期: 2014-11-06
- 网络出版日期: 2015-10-28





草地是我国陆地生态系统的重要组成部分。草地生物量不仅在全球碳循环和气候变化与草地生态系统之间的反馈作用中扮演着重要角色[1-4],而且决定着草地生态系统功能稳定性并作为系统能量流动和物质循环的基础,显著影响生态系统的结构[5]。此外,草地生物量的研究也为评估野生动物分布、合理规划草地资源、确定草地载畜量、草原火灾管理、生物多样性研究以及评估草地生态效益提供了重要参考依据[6-9]。因此快速、准确估测草地地上生物量及群落中物种生物量对草地生态系统管理和认识草地各物种的生态位的变化具有重要意义[10]。
目前,草地地上生物量测定的主要方法有直接收获法,该法结果准确,但繁琐、费时,而且对样地破坏性很大,尤其是对群落结构变化的长期监测试验难以实行[11-13]。另一种方法是基于与生物量密切相关的植被因子参数,通过数理方法建立草地地上生物量估测模型,该法可准确、迅速、无破坏性地估测草地生物量[14]。国内外关于植物地上生物量估测模型的研究多见于乔木和灌木的报道,而对草本植物生物量模型估测的研究多基于植被盖度、生物量作为参数[15-17]和群落物种高度和盖度建立了估测模型[18];也有根据大量地上观测数据并结合遥感数据建立了草地地上生物量指数和幂函数统计估测模型[19-20]。
以上研究多集中于植被生长比较均一的草地,但对于植被稀疏、生长不均一、生态环境比较脆弱的黄土高原典型草原地上生物量估测模型的研究尚未见报道。此外,单株生物量估测模型通常用于乔木和灌木,对于从个体水平建立草地生物量估测模型的研究尚未见报道。因此,本文分别通过对黄土高原典型草原植物种群水平和单株水平生物量的实地样方调查,选择株高(H)和盖度(C)的复合因子(C×H)为自变量,应用数理统计方法分别建立单株水平和种群水平不同物种地上生物量估测模型,并对其进行检验、比较其精确性,为该区地上生物量的测定提供了一个快速、准确且不具有破坏性的估测方法。
1 材料与方法 1.1 研究区自然概况研究区设在甘肃省榆中县兰州大学黄土高原国际地面与环境监测站围封草地(35°57′ N,104°09′ E),海拔1965.8 m。该地区为黄土高原残塬梁峁沟壑,属于大陆性半干旱性气候,年均气温6.7 ℃,年降水量约382 mm,年蒸发量1 343 mm,无霜期为90—140 a,年日照时数约2 600 h。该区域塬面原为传统的耕作农田,1986年撂荒,已基本恢复到天然植被。2005年10月设置围栏对草地进行围封。该区植被类型属于半干旱典型草原,植被组成主要为长芒草(Stipa bungeana)、阿尔泰狗娃花(Heteropappus altaicus)、赖草(Leymus secalinus)和米口袋(Gueldenstaedtia verna)等(表 1),土壤为灰钙土[21]。
| 物种Species | 相对高度Relative height | 相对盖度Relative coverage | 相对密度Relative abundance | 重要值Importance value |
| 长芒草Stipa bungeana | 0.29 | 0.51 | 0.25 | 0.35 |
| 阿尔泰狗娃花Heteropappus altaicus | 0.15 | 0.15 | 0.15 | 0.15 |
| 黄花蒿Artemisia annua | 0.09 | 0.06 | 0.13 | 0.09 |
| 黄花棘豆Leguminosae | 0.02 | 0.02 | 0.01 | 0.02 |
| 披针叶黄华Thermopsis lanceolata | 0.04 | 0.02 | 0.03 | 0.03 |
| 赖草Leymus secalinus | 0.18 | 0.02 | 0.07 | 0.09 |
| 米口袋Gueldenstaedtia multiflora | 0.04 | 0.06 | 0.06 | 0.05 |
| 糙隐子草Cleistogenes squarrosa | 0.15 | 0.17 | 0.16 | 0.16 |
| 无芒隐子草Cleistogenes songorica | 0.01 | 0.01 | 0.01 | 0.01 |
| 二裂委陵菜Potentilla bifurca | 0.04 | 0.01 | 0.01 | 0.02 |
| 莓叶委陵菜Potentilla fragarioides | 0.03 | 0.05 | 0.01 | 0.03 |
| 野胡麻Dodartia orientalis | 0.01 | 0.01 | 0.01 | 0.01 |
| 重要值=(相对高度+相对盖度+相对密度)/3 | ||||
在围封样地中选择坡向和地势平坦一致的地段,于2013年8月中旬草地植被生物量达到最大值时,分别进行单株和种群水平取样,单株每个物种随机选取25—600株,分别测量每株高度并用目测法估测其盖度,之后齐地面分株收获。种群水平测定在样地内随机选择80个1m×1m的样方,详细记录每个样方内的物种名称,各物种随机测量5株的高度,取平均值代表该物种高度,同时记录各物种的密度及用目测法估测各物种分盖度,之后将样方内植物分物种齐地面收获。收获的植物样品,带回室内,置于烘箱中于105℃杀青30 min,然后于65 ℃烘干至恒重,称重并计算干重。
1.3 数据分析数据采用 SPSS 20.0软件进行统计分析。采用留一分析法进行模型的建立与精确度的检验,即在所取样本中一部分作为估测模型参数,另一部分用于检验模型精确性。选用株高(H)和盖度(C)的复合因子(CH)为自变量,采用y=ax+b、y= ax2+ bx+c、y=axb等3种最常用的生物量估测模型,建立各物种和各物种单株的生物量估测方程,并通过比较判定系数R2、相对平均误差绝对值RMA和总相对误差RS确定最佳估测模型,同时比较种群水平和单株水平的植物地上生物量估测模型的差异性。将所选单株水平和种群水平合适生物量方程采用留一法带入到群落调查样方,进行群落生物量估测,以(1±10) %作为变化区间,比较两种水平所建生物量估测模型在群落水平的精确性。用Sigmaplot12.5进行制图。相对平均误差绝对值RMA和总相对误差RS计算公式如下[12]:
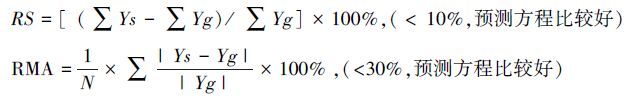
|
式中,N为实验样本数量;Ys 为生物量实测值;Yg为生物量估测值。
2 结果与分析 2.1 单株水平生物量估测模型的建立与验证 2.1.1 生物量估测模型的建立本研究区12种植物单株的生物量估测模型均达显著水平(P<0.05,图 1),且各物种的单株植物生物量均随复合因子(CH)增加而增加,但对于不同物种其增长方式并非完全一致。长芒草、糙隐子草(Cleistogenes squarrosa)、披针叶黄华(Thermopsis lanceolata)、二裂委陵菜(Potentilla bifurca)、莓叶委陵菜(Potentilla fragarioides)和无芒隐子草(Cleistogenes songorica)等6种物种在单株水平上生物量估测的最优模型为y=axb,R2均大于0.75;其中,糙隐子草的相关性最高(R2=0.909) ,二裂委陵菜的相关性最低(R2=0.679) 。米口袋、阿尔泰狗娃花、黄花蒿(Artemisia annua)、赖草、野胡麻(Dodartia orientalis)和黄花棘豆(Leguminosae)等6种植物在单株水平上生物量估测的最优模型为y=ax+b。R2均大于0.87,米口袋的相关性最高(R2=0.935) ,赖草的相关性最低(R2=0.876) 。

|
| 图 1 单株水平各物种最优生物量估测模型 Fig. 1 The best estimation models of species in individual level |
基于单株水平所建立的生物量估测模型,采用留一分析法对各物种的生物量估测值与实测值进行相关性分析,并对建立的生物量估测模型的精确性进行检验。结果表明,各个物种的生物量估测值与实测值相关性均达到了显著水平(P<0.05) ,平均相对误差绝对值RMA均在17%至30%之间,总相对误差RS(一般应<10%)除野胡麻和二裂委陵菜外均<10%(表 2)。
| 物种Species | 样本数Sample size | 标准误差Standard error | r | P | RMA/% | RS/% |
| 长芒草Stipa bungeana | 226 | 0.036 | 0.904 | 0.000* | 28.371 | 1.594 |
| 阿尔泰狗娃花Heteropappus altaicus | 140 | 0.045 | 0.889 | 0.000* | 26.114 | 2.842 |
| 黄花蒿Artemisia annua | 110 | 0.031 | 0.896 | 0.000* | 27.051 | 9.627 |
| 黄花棘豆Leguminosae | 25 | 0.107 | 0.756 | 0.000* | 23.996 | 0.101 |
| 披针叶黄华Thermopsis lanceolata | 40 | 0.076 | 0.887 | 0.000* | 27.712 | 5.739 |
| 赖草Leymus secalinus | 40 | 0.15 | 0.645 | 0.000* | 22.751 | 0.582 |
| 米口袋Gueldenstaedtia multiflora | 50 | 0.055 | 0.868 | 0.000* | 25.608 | -7.693 |
| 糙隐子草Cleistogenes squarrosa | 55 | 0.137 | 0.614 | 0.000* | 17.255 | 0.507 |
| 无芒隐子草Cleistogenes songorica | 15 | 0.068 | 0.991 | 0.000* | 16.956 | 8.197 |
| 二裂委陵菜Potentilla bifurca | 82 | 0.194 | 0.512 | 0.000* | 26.579 | 11.749 |
| 莓叶委陵菜Potentilla fragarioides | 21 | 0.151 | 0.734 | 0.120* | 27.878 | 7.654 |
| 野胡麻Dodartia orientalis | 31 | 0.096 | 0.816 | 0.000* | 30.179 | 19.644 |
| *P<0.05,RMA: 相对平均误差绝对值Average absolute value of relative error; RS: 总相对误差Total relative error | ||||||
由种群水平建立的生物量估测模型均达显著水平(P<0.05,图 2),且各物种的生物量均随复合因子(CH)增加而增加,所有物种生物量最佳估测模型主要表现为线性函数和幂函数。长芒草、旋风草、狗娃花、黄花蒿、米口袋、苦豆子和二裂委陵菜等7种物种的生物量估测的最优模型为y=axb。R2依次为糙隐子草(0.905) >米口袋(0.871) >披针叶黄华(0.837) >二裂委陵菜(0.836) >黄花蒿(0.798) >长芒草(0.736) >阿尔泰狗娃花(0.540) 。赖草、黄花棘豆和其他杂草等3种物种生物量估测的最优模型为y=ax+b。R2均大于0.85,其中,赖草的相关性最高(R2=0.966) ,黄花棘豆的相关性最低(R2=0.851) 。

|
| 图 2 种群水平各物种最优生物量估测模型 Fig. 2 The best estimation models of species in population level |
基于种群水平所建立的生物量估测模型,采用留一分析法将各物种的生物量估测值与实测值作相关性分析,同时对建立的生物量估测模型的精确性进行检验,结果表明(表 3),各个物种的生物量估测值与实测值相关性较好,均达到了显著水平(P<0.05) 。除黄花蒿、黄花棘豆和赖草等3种物种外,其余物种平均相对误差绝对值RMA均<30%,总相对误差RS除长芒草和阿尔泰狗娃花外均大于10%。
| 物种Species | 样本数Sample size | 标准误差Standard error | r | P | RMA/% | RS/% |
| 长芒草Stipa bungeana | 30 | 0.285 | 0.542 | 0.003* | 23.798 | -8.388 |
| 阿尔泰狗娃花Heteropappus altaicus | 30 | 0.118 | 0.82 | 0.000* | 27.148 | -4.431 |
| 黄花蒿Artemisia annua | 24 | 0.244 | 0.758 | 0.000* | 68.641 | 80.005 |
| 二裂委陵菜Potentilla bifurca | 18 | 0.136 | 0.941 | 0.002* | 26.669 | 10.804 |
| 黄花棘豆Leguminosae | 10 | 0.28 | 0.761 | 0.135* | 44.517 | 38.263 |
| 赖草Leymus secalinus | 17 | 0.047 | 0.88 | 0.000* | 30.021 | 65.572 |
| 米口袋Gueldenstaedtia multiflora | 17 | 0.766 | 0.723 | 0.001* | 27.356 | 33.577 |
| 披针叶黄华Thermopsis lanceolata | 14 | 0.118 | 0.968 | 0.000* | 21.95 | 24.216 |
| 糙隐子草Cleistogenes squarrosa | 18 | 0.092 | 0.93 | 0.000* | 24.841 | 19.217 |
| 其他杂草Other weeds | 19 | 0.531 | 0.564 | 0.045* | 28.979 | 25.911 |
分别利用由单株水平和种群水平所建立的单个物种的生物量估测模型,通过累加样方内不同物种的生物量估测值计算20个备检样方的总生物量,并且与20个备检样方的实测总生物量进行比较。通过图 3可知,由单株水平各物种生物量估测模型所得的估测值与实测值非常接近,多数样方的总生物量估测值与实测值均分布在y=x/(1±10%)的区域类。而通过种群水平的各物种生物量估测模型所得的估测值均大于实测值,如图 3所示,所有样方的总生物量估测值与实测值均分布在y=x/(1±10%)的区域上部。

|
| 图 3 两种不同水平地上总生物量实测值与估测值的比较 Fig. 3 Comparison of measured aboveground biomass value and estimated aboveground biomass value in different levels |
通过生物量估测模型计算得到的估测生物量与实测生物量之间的相对关系可以用线性方程较好的表达出来。由生物量实测值与估测值的线性回归分析结果(表 4)表明:在单株水平上,长芒草、阿尔泰狗娃花、黄花蒿、黄花棘豆、披针叶黄华、赖草、米口袋、糙隐子草、无芒隐子草、二裂委陵菜、莓叶委陵菜和野胡麻的校正系数分别为1.050、1.028、0.619、0.709、0.938、1.030、0.833、0.923、0.936、1.051、0.961和0.794,R2值分别为0.893、0.828、0.860、0.719、0.882、0.889、0.891、0.874、0.981、0.701、0.538和0.663,均达到了显著水平(P<0.005) 。
| 水平Level | 物种Species | 拟合方程Fitting equation | 校正系数Correctioncoefficient | 标准误差Standard error | R2 | t | 样本数量Sample size |
| 单株水平 | 长芒草Stipa bungeana | Y=1.050x | 1.05 | 0.025 | 0.893 | 42.61 | 226 |
| Individual | 阿尔泰狗娃花Heteropappus altaicus | Y=1.028x | 1.028 | 0.041 | 0.828 | 25.468 | 140 |
| level | 黄花蒿Artemisia annua | Y=0.619x | 0.619 | 0.022 | 0.86 | 25.303 | 110 |
| 黄花棘豆Leguminosae | Y=0.709x | 0.709 | 0.07 | 0.719 | 7.333 | 25 | |
| 披针叶黄华Thermopsis lanceolata | Y=0.938x | 0.938 | 0.054 | 0.882 | 16.382 | 40 | |
| 赖草Leymus secalinus | Y=1.030x | 1.03 | 0.061 | 0.889 | 16.708 | 40 | |
| 米口袋Gueldenstaedtia multiflora | Y=0.833x | 0.833 | 0.038 | 0.891 | 19.36 | 51 | |
| 糙隐子草Cleistogenes squarrosa | Y=0.923x | 0.923 | 0.05 | 0.874 | 18.619 | 53 | |
| 无芒隐子草Cleistogenes songorica | Y=0.936x | 0.936 | 0.068 | 0.981 | 14.436 | 6 | |
| 二裂委陵菜Potentilla bifurca | Y=1.051x | 1.051 | 0.093 | 0.701 | 13.088 | 82 | |
| 莓叶委陵菜Potentilla fragarioides | Y=0.961x | 0.961 | 0.158 | 0.538 | 4.451 | 21 | |
| 野胡麻Dodartia orientalis | Y=0.794x | 0.794 | 0.165 | 0.663 | 5.779 | 21 | |
| 种群水平 | 长芒草Stipa bungeana | Y=0.924x | 0.924 | 0.246 | 0.437 | 4.407 | 30 |
| Population | 阿尔泰狗娃花Heteropappus altaicus | Y=0.880x | 0.88 | 0.09 | 0.737 | 8.544 | 30 |
| level | 黄花蒿Artemisia annua | Y=1.611x | 1.611 | 0.207 | 0.715 | 7.262 | 24 |
| 二裂委陵菜Potentilla bifurca | Y=1.066x | 1.066 | 0.167 | 0.886 | 6.235 | 7 | |
| 黄花棘豆Leguminosae | Y=0.606x | 0.606 | 0.28 | 0.579 | 2.032 | 5 | |
| 赖草Leymus secalinus | Y=0.344x | 0.344 | 0.047 | 0.775 | 7.184 | 17 | |
| 米口袋Gueldenstaedtia multiflora | Y=4.050x | 4.05 | 0.733 | 0.649 | 4.908 | 17 | |
| 披针叶黄华Thermopsis lanceolata | Y=1.430x | 1.43 | 0.118 | 0.937 | 13.364 | 14 | |
| 糙隐子草Cleistogenes squarrosa | Y=1.129x | 1.129 | 0.106 | 0.878 | 9.3 | 14 | |
| 其他杂草Other weeds | Y=0.725x | 0.725 | 0.139 | 0.636 | 3.965 | 13 | |
| 总生物量 | 单株水平Individual level | Y=0.980x | 0.98 | 0.1 | 0.794 | 8.33 | 20 |
| Total biomass | 种群水平Population level | Y=0.718x | 0.718 | 0.123 | 0.524 | 4.456 | 20 |
在种群水平上,长芒草、阿尔泰狗娃花、黄花蒿、二裂委陵菜、黄花棘豆、赖草、米口袋、披针叶黄华、糙隐子草和其他杂草的校正系数分比为0.924、0.880、1.611、1.066、0.606、0.344、4.050、1.430、1.129和0.725,相对应的R2值分别为0.437、0.737、0.715、0.886、0.579、0.775、0.649、0.937、0.878和0.636,均达到了显著水平(P<0.005) 。在群落总生物量上,实测值与估测值之间也存在着比较好的线性关系,单株水平估测的总生物量的校正系数和R2值分别为0.980和0.794,种群水平估测的总生物量的校正系数和R2值分别为0.718和0.524,通过总生物量的校正系数和R2值的比较,说明单株水平所建的生物量估测模型的精确度优于种群水平所建的生物量估测模型。
3 讨论生物量是研究生态系统基本功能和过程重要参数因子,同时也是研究植物群落结构最重要的特征之一[22]。研究表明以自变量株高和盖度建立估测模型时,其操作比较简单,而且精确度也比较高[23]。因此本研究以植被株高(H)和盖度(C)的复合因子(CH)为自变量,分别从单株水平和种群水平对黄土高原典型草地各个物种进行生物量估测模型的建立,同时采用留一法并结合决定系数r、相对平均误差绝对值RMA和总相对误差RS等对所建生物量估测模型精确性进行评估,并通过校正系数以及群落总生物量估测值和实测值比较单株水平和种群水平所建模型的精确性。结果表明,在黄土高原典型草地,通过对生态参数因子的选择与筛选,无论在单株水平还是种群水平,线性和非线性(幂函数)对该区域生物量的拟合效果更好。这可能是因为在黄土高原地区,由于降水较少,土壤比较贫瘠等原因,致使该区域植物地上和地下的分配模式和过程发生了变化[24],也有研究表明其它模型应用比较少的原因是由于其生态学意义不明确,而且容易产生过度拟合现象[25]。这与张修玉[26]等研究结果表明线性回归方程在进行草地地上生物量估测时,其具有直观、简单的优点一致。我们采用留一分析法对所建立的生物量估测模型的精确性进行检验。检验结果表明在单株水平各个物种的生物量估测值与实测值相关性较好,均达到了显著水平(P<0.05) ,其r值均大于0.6,所有物种生物量估测模型的总相对误差RS除在群落中分布较少的中野胡麻和二裂委陵菜外均小于10%,平均相对误差绝对值RMA均小于30%(图 1)。在种群水平上,虽然各个物种的生物量估测值与实测值相关性均达到了显著水平(P<0.05) ,但多数物种平均相对误差绝对值RMA大于30%,总相对误差RS除长芒草和阿尔泰狗娃花均大于10%(图 2),说明通过单株水平建立的物种生物量估测模型的精度优于种群水平建立的物种生物量估测模型的精度。而且通过两种水平对群落总生物量的估测值和实测值进行比较时发现,在单株水平建立的生物量估测模型的实测值与估测值比较接近,均分布于y=x/(1±10%)的区域类,而在种群水平建立的生物量估测模型的估测值均大于实测值,所有样方的总生物量估测值与实测值均分布于y=x/(1±10%)的区域上部(图 3),其也说明单株水平建立的物种生物量估测模型的精度优于种群水平建立的物种生物量估测模型的精度。这与张典业等[10]人在种群水平,通过提取不同物种生物量密切相关的盖度、高度等植被因子,建立了青藏高原东缘高寒草甸地上生物量的估测模型,地上生物量估测模型在检验时,估测值与实测值拟合效果比较好的结果不相复合。这可能由于在单株水平建立的物种生物量估测模型在提取生态参数因子比较精细,而且样本数也比较多,而在种群水平建立的物种生物量估测模型时,由于盖度以及高度测量的不准确性,产生了比较大的误差。尤其在黄土高原典型草地,由于其植被稀疏以及植被生长的不均一性,通过群落组成以及重要值也发现(表 1),在黄土高原典型草原只有长芒草、赖草、狗娃花和黄花蒿的重要值接近或大于0.1,而其他物种重要值比较小,此时用平均高度代替草层高度进行生物量估测模型的建立更容易产生误差。在校正系数方面也说明了这一点,单株水平建立的生物量的估测模型的校正系数均接近1,而种群水平的校正系数均偏离1(表 4)。因此,在植被比较稀疏或植被生长不均一的草地,通过提取植被因子参数建立生物量估测模型时,应通过单株水平而不是通过种群水平建立地上生物量估测模型,这样可以提高生物量估测模型的精确性和适用性。
因此在进行草本生物量以及相关的生态参数因子调查时,应在允许的条件下多增加调查的样本数量以及提高所测生态参数因子的次数,可以通过单株水平进行相关生态因子的提取,从而提高草本样方的生物量估测模型的精度。本文建立的草本样方的生物量估测模型只适用于黄土高原典型草地,为其他研究者估测黄土高原典型草地生物量提供便利,尤其是对于那些长期在黄土高原典型草地进行长期的固定样方观测相关因素对群落结构的组分、生物量等研究提供了理论基础和依据。对于研究区以外的草地生物量估测还需要进一步验证,由于植被的生长状况会因土壤、气候、群落结构等立地条件的不同而发生变化[27-29],生物量估测模型的建立所选择的参数因子也会不同。除此以外,当所测草本相关变量超出所建模型的测定范围时应用我们所建立的生物量估测模型估测生物量还需要进一步验证。
| [1] | Piao S L, Fang J Y, Ciais P, Peylin P, Huang Y, Sitch S, Wang T.The carbon balance of terrestrial ecosystems in China.Nature,2009,458(7241): 1009–1013. |
| [2] | Fang J Y, Guo Z D, Piao S L, Chen A P.Terrestrial vegetation carbon sinks in China, 1981-2000.Science in China Series D:Earth Sciences,2007,50(9): 1341–1350. |
| [3] | Ma W H, Fang J Y, Yang Y H, Anwar Mohammat.Biomass carbon stocks and their changes in northern China's grasslands during 1982-2006.Science China Life Sciences,2010,53(7): 841–850. |
| [4] | Scurlock J M O, Johnson K, Olson R J.Estimating net primary productivity from grassland biomass dynamics measurements.Global Change Biology,2002,8(8): 736–753. |
| [5] | Ni J.Carbon storage in grasslands of China.Journal of Arid Environments,2002,50(2): 205–218. |
| [6] | Mutanga O, Skidmore A K.Narrow band vegetation indices overcome the saturation problem in biomass estimation.International Journal of Remote Sensing,2004,25(19): 3999–4014. |
| [7] | Mutanga O, Skidmore A K.Hyperspectral band depth analysis for a better estimation of grass biomass (Cenchrus ciliaris) measured under controlled laboratory conditions.International Journal of Applied Earth Observation and Geoinformation,2004,5(2): 87–96. |
| [8] | Ren H R, Zhouc G S, Zhang X S.Estimation of green aboveground biomass of desert steppe in Inner Mongolia based on red-edge reflectance curve area method.Biosystems Engineering,2011,109(4): 385–395. |
| [9] | Duan M J, Gao Q Z, Wan Y F, Li Y, Guo Y Q, Ganzhu Z B, Liu Y T, Qin X B.Biomass estimation of alpine grasslands under different grazing intensities using spectral vegetation indices.Canadian Journal of Remote Sensing:Journal Canadien de Télédétection,2014,37(4): 413–421. |
| [10] | 张典业, 牛得草, 陈鸿洋, 张永超, 傅华.青藏高原东缘高寒草甸地上生物量的估测模型.山地学报,2014,32(4): 453–459. |
| [11] | 李春萍, 李刚, 肖春旺.异速生长关系在陆地生态系统生物量估测中的应用.世界科技研究与发展,2007,29(2): 51–57. |
| [12] | 刘陟, 黄奇, 周延林, 李政海, 孙振, 柳琳秀, 米红胤, 樊亚娟.毛乌素沙地油蒿生物量估测模型研究.中国草地学报,2014,36(4): 24–30. |
| [13] | Redjadj C, Duparc A, Lavorel S, Grigulis K, Bonenfant C, Maillard D, Saïd S, Loison A.Estimating herbaceous plant biomass in mountain grasslands:a comparative study using three different methods.Alpine Botany,2012,122(1): 57–63. |
| [14] | 黄劲松, 邸雪颖.帽儿山地区6种灌木地上生物量估算模型.东北林业大学学报,2011,39(5): 54–57. |
| [15] | Matilla E.Survey of reindeer winter ranges as a part of the Finnish National Forest inventory in 1976-1978.Communicationes Instituti Forestalis Fenniae,1981,99(6): 1–74. |
| [16] | Flombaum P, Sala O E.A non-destructive and rapid method to estimate biomass and aboveground net primary production in arid environments.Journal of Arid Environments,2007,69(2): 352–358. |
| [17] | Montès N.A non-destructive method to estimate biomass in arid environments:A comment on Flombaum and Sala (2007).Journal of Arid Environments,2009,73(6/7): 599–601. |
| [18] | Axmanová I, Tichy L, Fajmonová Z, Hájková P, Hettenbergerová E, Li C F, Merunková K, Nejezchlebová M, Otypková Z, Vymazalová M and Zeleny D.Estimation of herbaceous biomass from species composition and cover.Applied Vegetation Science,2012,15(4): 580–589. |
| [19] | Piao S L, Fang J Y, Zhou L M, Tan K, Tao S.Changes in biomass carbon stocks in China's grasslands between 1982 and 1999.Global Biogeochemical Cycles,2007,21(2): GB2002 |
| [20] | 辛晓平, 张保辉, 李刚, 张宏斌, 陈宝瑞, 杨桂霞.1982-2003年中国草地生物量时空格局变化研究.自然资源学报,2009,24(9): 1582–1592. |
| [21] | Li X D, Fu H, Li X D, Guo D, Dong X Y, Wan C G.Effects of land-use regimes on carbon sequestration in the Loess Plateau, Northern China.New Zealand Journal of Agricultural Research,2008,51(1): 45–52. |
| [22] | 刘长成, 魏雅芬, 刘玉国, 郭柯.贵州普定喀斯特次生林乔灌层地上生物量.植物生态学报,2009,33(4): 698–705. |
| [23] | 杨昆, 管东生.森林林下植被生物量收获的样方选择和模型.生态学报,2007,27(2): 705–714. |
| [24] | 李旭东, 张春平, 傅华.黄土高原典型草原草地根冠比的季节动态及其影响因素.草业学报,2012,21(4): 307–312. |
| [25] | Gao J.Quantification of grassland properties:how it can benefit from geoinformatic technologies?.International Journal of Remote Sensing,2006,27(7): 1351–1365. |
| [26] | 张修玉, 许振成, 宋巍巍, 管东生, 赵晓光, 胡习邦, 张钢锋.西双版纳紫茎泽兰生物量收获的样方选择与模型.草业科学,2010,27(10): 85–90. |
| [27] | 杨昊天, 李新荣, 王增如, 贾荣亮, 刘立超, 高艳红, 李刚.腾格里沙漠东南缘4种灌木的生物量预测模型.中国沙漠,2013,33(6): 1699–1704. |
| [28] | 除多, 德吉央宗, 姬秋梅, 唐红.西藏高原典型草地地上生物量遥感估算.国土资源遥感,2013,25(3): 43–50. |
| [29] | 孙晓芳, 岳天祥, 王情.草地地上生物量高精度曲面建模.遥感学报,2013,17(5): 1068–1076. |