 2016, Vol. 36
2016, Vol. 36文章信息
- 胡晶晶, 原国辉, 李洋洋, 郭线茹, 王琼, 郭帅帅, 李为争
- HU Jingjing, YUAN Guohui, LI Yangyang, GUO Xianru, WANG Qiong, GUO Shuaishuai, LI Weizheng.
- 棉铃虫低龄幼虫对花香挥发物的联系性学习
- Associative learning in early-instar larvae of Helicoverpa armigera in response to several floral volatiles
- 生态学报[J]. 2016, 36(13): 4204-4210
- Acta Ecologica Sinica[J]. 2016, 36(13): 4204-4210
- http://dx.doi.org/10.5846/stxb201411022145
-
文章历史
- 收稿日期: 2014-11-02
- 网络出版日期: 2015-10-29

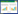




联系性学习是昆虫在复杂多变的环境中适应的基本机制之一。当前联系性学习行为的研究主要集中在成虫阶段,受学习调控的行为包括觅食、产卵寄主选择、聚集、避敌甚至与交配有关的性信息素通讯等[1-6]。可能是由于成虫迁移能力和幼虫相比强得多,涉及的生命行为不仅仅是取食。鳞翅目的105种昆虫大田统计资料表明,低龄幼虫面临着各种死亡因子的威胁,自然死亡率平均为66%—82% [7],但幼虫也能通过近距离的扩散进行有限的逃避[8-10],例如海灰翅夜蛾Spodoptera littoralis幼虫可以通过调整自身的取食位置,矫正母代雌成虫在棉花植株上不适当的产卵位置选择[10]。很显然取食是低龄幼虫期最关键的生命行为,如果低龄幼虫在取食过程中能够事先记住一些与食物伴随的关键化学信息,则在因机械扰动、捕食未遂或干旱等被迫离开寄主植物后,有助于搜索与已适应的植物气味相似的种类,从而降低寄主转移后被迫重新调整代谢途径而付出的生理代价[10]。弄清幼虫联系性学习的规律,有助于通过人为干预农业生态系统来强化这些发育阶段自然死亡因子的效果,并为培育不易被幼虫取食适应的作物品种提供参考依据。然而,目前幼虫联系性学习仅在海灰翅夜蛾 [11]、苹果蠹蛾Cydia pomonella[12]和黑腹果蝇Drosophila melanogaster[13]中报道过,而经济重要性很高的大部分农业害虫还未见相关研究。
棉铃虫Helicoverpa armigera是实夜蛾属的一种多食性种类,低龄幼虫嗜食许多农作物的花器[14],这些器官与营养生长部分的挥发性物质有较大的差异,前者以芳香族化合物和单萜类为主,后者以6个碳原子的绿叶气味为主。此前本课题组以蔗糖溶液为无条件刺激研究了成虫对2种关键花香气味的联系性学习,发现成虫对苯乙醛的学习表现比乙酸苯甲酯更好,且外周感觉系统水平的改变不能解释学习诱导的气味偏好性变化 [15]。棉铃虫幼虫不能象成虫一样单独依赖蔗糖等纯化学物质生长发育。为此,作者使棉铃虫初孵幼虫在人工饲料(无条件刺激)取食时,分别暴露于7种花香气味下连续7 d,逐日取样测试其学习表现;然后,为了判断气味偏好性的改变是生理性成熟还是联系性学习造成的,测试了不同日龄的无经历幼虫对苯乙醛的偏好反应,期望弄清低龄幼虫是否具有嗅觉联系性学习能力,以及学习表现与同种成虫相比的强弱。
1 材料与方法 1.1 供试昆虫棉铃虫蛹购于河南济源白云实业有限公司,放在室内人工气候箱内饲养,饲养条件为日温(28±2) ℃,夜温(26±2) ℃,相对湿度(50±10) %,光周期16 L∶8 D。成虫饲喂5%蔗糖溶液,幼虫用麦胚基饲料连续饲养10代以上,在本试验中该饲料也用作无条件刺激物。制备方法如下:将1400 mL蒸馏水煮沸并分成两份,一份600 mL,另一份800 mL。第1份中加入3 g山梨酸并搅拌,完全溶解后加入150 g小麦胚芽粉、80 g黄豆粉和25 g烘焙酵母;第2份中加入20 g琼脂、20 g蔗糖和40 g干酪素并搅拌均匀。随后将二者混合,待温度降至40℃左右时,加入3 g 维生素C和8g复合维生素,最终搅拌均匀并在4 ℃下保存。
1.2 供试试剂条件刺激为7种花香挥发物:乙酸苯甲酯、苯甲醛、苯乙醛、苯甲醇、β-苯乙醇、香叶醇和外消旋芳樟醇。苯甲醛、苯甲醇购于北京化学试剂公司;乙酸苯甲酯购于中国医药集团(上海)化学试剂公司,纯度为95%;苯乙醛购买于Sigma-Aldrich Co Ltd.,纯度为98%;β-苯乙醇、芳樟醇、香叶醇购于Fluka 化学试剂公司,分析纯。上述花香挥发物各200μL分别注入10 mL玻璃瓶中,将煮沸后并在室温下冷却到60℃左右的2%琼脂(Amresco,99%)胶液分装到各个小瓶中(8 mL/瓶),迅速盖紧瓶塞并摇匀,当瓶壁温度与室温接近时停止摇动使样品凝固,制成0.5 cm ID×0.5 cm H柱状小块,条件化之后的气味偏好性测试仍然使用这些气味源,另外还制备了空白琼脂胶作为对照。
1.3 学习训练收集棉铃虫卵放入人工气候箱中,孵化条件同上。每日8:00. 和17:00. 各检查一次孵化情况,获得日龄整齐的幼虫。每日18: 00. 取当日孵化个体随机分配到两个试验组:(1) 单独US暴露组(US-only group):在14.0 cm ID × 2.0 cm H培养皿底部铺上湿滤纸,正中央放入3.0 cm L × 3.0 cm W× 1.5 cm H的饲料块,每个饲料块上部中央插放一支装有琼脂胶柱的1 mL离心管。小心挑取20头初孵幼虫放在饲料块上,盖上盖子之后放入人工气候箱;(2) 条件化组(Conditioning group):操作同上,饲料块上方离心管中放入不同花香挥发物制备的琼脂胶柱,使幼虫在花香气味氛围下取食24 h。本组又分为7个亚组,分别使用上述7种花香气味作为条件刺激。不同处理组放在隔离环境中,很少发现训练期间试虫逃逸到培养皿外面的现象。
1.4 气味偏好性测定为了克服情境依赖性学习现象,气味偏好性测试所用装置仍为14.0 cm ID × 2.0 cm H的培养皿,且测试气味源仍然使用和上述学习训练相同的花香挥发物琼脂胶柱和空白琼脂胶柱进行配对。训练后24 h进行第1次测试。测试在洁净无味的暗室中进行,在底部铺有湿滤纸的培养皿内,沿一条直径的两端分别放置花香挥发物琼脂胶柱和空白琼脂胶柱,然后由3人配合同时在每个培养皿中放入相应训练的幼虫5头,盖上盖子后立刻放在1.5 W红色LED灯泡下观察。幼虫在湿滤纸上爬行一段时间后,会移动到处理或对照琼脂胶柱上。及时移除做出选择反应的个体以便使社会性相互作用最小化,直到所有5头幼虫全部做出选择为止。预试验发现一般在10 min测试时间内所有个体均会做出选择。测试结束后,试虫返回相应处理环境中继续学习训练,但更换新的培养皿、滤纸、饲料块和气味源。24 h进行第2次测试,依此类推,直到训练后第7天,以便确定训练时长对学习表现的影响,该过程跨越了2个龄期。7种挥发物的条件化组每组累计测试50头个体。
1.5 不同日龄的无经历组幼虫对苯乙醛的趋性反应在供试的7种花香挥发物中苯乙醛条件化效应最明显。因此,测试了无经历组幼虫对苯乙醛 vs 空白琼脂胶的偏好选择反应,用来鉴别联系性学习效应与生理性成熟。为了同条件化组测试对应,仍然跟踪测试7 d。不同之处在于每批幼虫仅测试1次,即将孵化当日的大量幼虫放在人工饲料上饲养,24 h后挑取50头进行第1次测定,48 h后另外挑取50头进行第2次测定,依此类推。
1.6 统计分析采用SPSS 19.0统计软件分析数据。每个处理组每次测试得到的是二项分布的频次数据,采用Yate修正的χ2测验来分析两种气味源被选择频次的差异。
2 结果与分析 2.1 棉铃虫低龄幼虫对7种花香挥发物的联系性学习表现时相棉铃虫低龄幼虫经7种常见花香挥发物不同时间的条件化后在条件化气味源和对照配对测试条件下的趋性反应结果见图 1,对应的χ2测验见表 1。

|
| 图 1 棉铃虫低龄幼虫对花香挥发物-食物联结联系性学习表现的时相 Fig. 1 Temporal pattern of the associative learning performance of H. armigera early-instar larvae to several floral volatiles associated with the artificial diet |
| 条件刺激 Conditioning stimuli | 经历不同条件化时间(d)的气味选择频次卡方值χ2 values of choice frequencies subjected to different times (d) of conditioning | ||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| 苯乙醛 Phenylacetaldehyde | 6.63 | 0.02 | 12.50 | 5.78 | 0.02 | 1.62 | 1.62 |
| 苯甲醛 Benzaldehyde | 2.42 | 1.62 | 0.98 | 0.02 | 0.02 | 0.50 | 0.50 |
| 芳樟醇 Linalool | 0.98 | 0.02 | 1.62 | 4.50 | 0.02 | 10.58 | 1.62 |
| 乙酸苯甲酯 Benzyl acetate | 2.42 | 0.02 | 1.62 | 0.02 | 0.50 | 0.02 | 0.18 |
| 香叶醇 Geraniol | 0.02 | 0.18 | 0.50 | 0.98 | 0.02 | 0.02 | 0.50 |
| 苯甲醇 Phenyl methanol | 0.50 | 0.98 | 0.02 | 0.02 | 0.18 | 0.02 | 0.02 |
| 苯乙醇 Phenethylalcohol | 0.02 | 1.62 | 4.50 | 3.38 | 0.98 | 0.18 | 0.02 |
在7次测定中,苯乙醛条件化组第1天、第3天和第4天表现出对苯乙醛的显著或极显著偏好性(第1天:χ2= 6.63;第3天:χ2= 12.50;第4天:χ2= 5.78) ,尤其是第1天和第3天,选择苯乙醛的百分率分别达到了70%和76%,芳樟醇条件化组在2次测定中处理被选频次显著或极显著多于对照(第4天:χ2=4.50;第6天:χ2=10.58) ,苯乙醇条件化组仅有1次测定中对苯乙醇的选择频次显著多于对照(第3天:χ2= 4.50) 。无论经历多长训练时间,对苯甲醛、苯甲醇、乙酸苯甲酯和香叶醇均不会表现出显著嗅觉偏好。另外,处理选择百分率的变化趋势与条件化时间长短没有明显关系,例如苯乙醛和苯乙醇条件化时第3天选择处理的百分率最高,而芳樟醇条件化时则是第5天最高。
2.2 不同日龄的无经历组幼虫对苯乙醛的嗅觉偏好性棉铃虫雌蛾对苯乙醛的学习表现强于对乙酸苯甲酯的学习表现[15],上述测试再次发现苯乙醛条件化组的棉铃虫低龄幼虫对苯乙醛的选择频次显著高于对照的次数最多。因此,进一步研究了无花香挥发物接触经历的棉铃虫低龄幼虫在苯乙醛 vs CK配对条件下的嗅觉选择反应,结果如图 2。无经历组幼虫无论24 h取食后经历的时间长度如何,均没有对苯乙醛表现出明显的偏好,证实上述测试中气味偏好性的改变并非生理性成熟所致。

|
| 图 2 不同日龄的无经历组棉铃虫幼虫对苯乙醛的气味偏好性 Fig. 2 Olfactory preference of nave H. armigera larvae of different day-ages to phenylacetaldehyde |
自然界鳞翅目初孵幼虫能量储备有限[16],却面临着大量的威胁,如叶毛、叶蜡、坚硬组织、乳胶管、腺体、毒素、植物诱导防卫、多变的微栖境和生物致死因子等[7]。低龄幼虫可以通过有限的扩散运动主动逃避这些严酷的条件[13, 17-18]。棉铃虫多在寄主花器或果实上取食,取食过程中如果能够记忆某些关键花香挥发物,在被迫离开寄主植物后就更容易重新定殖到同一植物种类上,降低食物混合的负面影响[19]。每次训练中条件刺激和无条件刺激的间隔(inter-stimulus interval)和相邻训练的间隔(inter-trial internal)严格控制,海灰翅夜蛾和黑腹果蝇幼虫才有较好学习表现[7, 9]。然而,幼虫野外取食环境始终弥散着寄主挥发物,这样的条件下试虫是否仍然会表现出良好学习能力尚不清楚。
选择7种花香挥发物研究了棉铃虫低龄幼虫的联系性学习,这些挥发物在许多寄主植物花器的顶空挥发物中发现[20]。其中,棉铃虫的姊妹种烟青虫H. assulta成虫受开花期烟草花顶空挥发物中的外消旋芳樟醇引诱[21],在棉铃虫[5]、烟芽夜蛾Heliothis virescens[22]、烟草天蛾Manduca sexta[23]成虫学习研究中使用过;烟芽夜蛾、棉铃虫和烟青虫成虫触角存在对香叶醇反应最强的感觉神经元[24],该物质在烟芽夜蛾[6, 22]、海灰翅夜蛾[7, 25-26]和烟草天蛾[27]成虫学习研究中使用过;夜蛾花香引诱剂的2种关键成分[28]苯乙醛和乙酸苯甲酯在棉铃虫[5, 15]和海灰翅夜蛾[26]学习研究中使用过。棉铃虫初孵幼虫仅涉及取食的行为需求,需要以食物本身作为无条件刺激,然而幼虫不能像成虫一样依赖蔗糖溶液就能存活,自然寄主作为无条件刺激又难以避免寄主本身气味的干扰,故采用人工饲料作为无条件刺激,这种饲料的整体挥发物当前尚未报道,但主要原料小麦胚芽粉、黄豆粉、烘焙酵母的主要挥发物成分已有报道。其中,小麦麸皮主要成分为挥发性很弱的脂肪酸类和一些酚酸、黄酮等[29],小麦全粉顶空挥发物成分主要是(E,E)-2,4-癸二烯醛(37.46%)和十二碳-2-烯醛(11.88%),黄豆粉顶空挥发物的主要成分是正己醇(24.77%)、正己醛(25.50%)、1-辛烯-3-醇(19.93%)和E-2-己烯醛(11.38%)[30],酵母浸膏去除作者分析明显杂质后,重新计算的主要物质及相对含量是乙酸(38.66%)、吲哚(16.70%)和2-乙基己醇(12.22%)[31]。这些植物材料均不含有本试验的7种花香挥发物,或者虽然痕量存在但不会显著影响试验结果。苯乙醛条件化组在7次测定中有3次对苯乙醛表现出显著或极显著偏好,芳樟醇条件化组在2次测定中对芳樟醇表现出显著或极显著偏好,苯乙醇条件化组仅1个案例对苯乙醇表现出显著偏好,其他4种挥发物无论训练多长时间嗅觉偏好性也不显著改变,说明棉铃虫在取食过程中并非记忆所有挥发物信息,而是选择性地采集一些关键信息,如各种显花植物的花器中最普遍存在的苯乙醛。此外,棉铃虫低龄幼虫有避开油腺取食的习性,大龄幼虫则偏好到花器上取食[32],暗示着棉铃虫嗅觉偏好性可能随着日龄而变化。以苯乙醛为代表性物质测试了低龄幼虫嗅觉偏好性随着日龄的变化,发现所有日龄的试虫对苯乙醛均没有显著偏好,暗示着生理性成熟不会造成嗅觉偏好性改变,证实这种学习的本质是联系性的,未来需要全程检测整个幼虫发育期气味偏好性的变化情况。
然而,棉铃虫低龄幼虫的联系性学习表现较差,且与训练时间长度关系不大。可能原因如下:(1) 在物种水平上,学习发生在需要持续搜索寄主的虫态[33]。若虫期活跃的黄斑黑蟋蟀Gryllus bimaculatus、马德拉蜚蠊Leucophaea maderae等需要不断探索食物源,具有极强嗅觉学习能力[34-35];相反,鳞翅目初孵幼虫选择取食位置的能力非常有限,很大程度上依赖成虫产卵决策;(2) 棉铃虫成虫采集花蜜时对嗅觉的依赖性远比味觉强,故表现出相对较强的嗅觉学习能力[3-5, 15],而味觉信息对于幼虫食物的指示作用比嗅觉信息更可靠,因为幼虫发育期大部分时间是在植物挥发物相似、但味觉信息差异较大的同一植株不同部位转移为害的,未来需要采用呈味物质作为条件刺激研究其联系性学习来证实。但寻找一种味觉系统能够感知但呈行为学中性的味觉条件刺激并不容易,许多人类基本呈味物质本身对幼虫取食选择有较大的影响[36]。目前鳞翅目幼虫联系性学习仅在苹果蠹蛾[12]和海灰翅夜蛾[11]中报道过,但只有苹果蠹蛾研究的是味觉学习(条件刺激是糖精);(3) 以往昆虫学习的经典条件化步骤中,是将条件刺激与无条件刺激前配对进行若干次训练,训练间隔期被试置于不存在条件刺激和无条件刺激的洁净空间中休息[25, 27, 37]。这种周期性的配对方式可能更容易使条件刺激成为无条件刺激的标志性信息。为了模拟自然生态环境,本试验测试前和训练过程中均没有剥夺试虫的食物,也会潜在削弱花香挥发物的信息指示功能。此前的昆虫嗅觉学习的种间比较表明,食性杂、社会性、寿命长、体型大的种类学习能力更突出[2, 22]。学习的首要功能是使生物更好地适应多变的环境,某些种类尽管具有利用多种寄主植物的潜能,但在个体发育中如果不需要持续开拓食物资源,就不会有很强的学习表现。
| [1] | Dukas R.Evolutionary biology of insect learning.Annual Review of Entomology,2008,53(1): 145–160. |
| [2] | Kandori I, Yamaki T.Reward and non-reward learning of flower colours in the butterfly Byasa alcinous (Lepidoptera:Papilionidae).Naturwissenschaften,2012,99(9): 705–713. |
| [3] | Cunningham J P, West S A, Wright D J.Learning in the nectar foraging behaviour of Helicoverpa armigera.Ecological Entomology,1998,23(4): 363–369. |
| [4] | Cunningham J P, Jallow M F A, Wright D J, Zalucki M P.Learning in host selection in Helicoverpa armigera (Hübner) (Lepidoptera:Noctuidae).Animal Behaviour,1998,55(1): 227–234. |
| [5] | Cunningham J P, Moore C J, Zalucki M P, Cribb B W.Insect odour perception:recognition of odour components by flower foraging moths.Proceedings of the Royal Society Biological Sciences B:biological sciences,2006,273(1597): 2035–2040. |
| [6] | Hartlieb E, Hansson B S, Anderson P.Sex or food? Appetetive learning of sex odors in a male moth.Naturwissenschaften,1999,86(8): 396–399. |
| [7] | Zalucki M P, Clarke A R, Malcolm S B.Ecology and behavior of first instar larval Lepidoptera.Annual Review of Entomology,2002,47(1): 361–393. |
| [8] | Perkins L E, Cribb B W, Hanan J, Glaze E, Beveridge C, Zalucki M P.Where to from here? The mechanisms enabling the movement of first instar caterpillars on whole plants using Helicoverpa armigera (Hübner).Arthropod-Plant Interactions,2008,2(4): 197–207. |
| [9] | Piesik D, Rochat D, van der Pers J, Marion-Poll F.Pulsed odors from maize or spinach elicit orientation in European corn borer neonate larvae.Journal of Chemical Ecology,2009,35(9): 1032–1042. |
| [10] | Sadek M M.Complementary behaviors of maternal and offspring Spodoptera littoralis:oviposition site selection and larval movement together maximize performance.Journal of Insect Behavior,2011,24(1): 67–82. |
| [11] | Salloum A, Colson V, Marion-Poll F.Appetitive and aversive learning in Spodoptera littoralis larvae.Chemical Senses,2011,36(8): 725–731. |
| [12] | Pszczolkowski M A, Brown J J.Single experience learning of host fruit selection by Lepidopteran larvae.Physiology & Behavior,2005,86(1/2): 168–175. |
| [13] | Hendel T, Michels B, Neuser K, Schipanski A, Kaun K, Sokolowski M B, Marohn F, Michel R, Heisenberg M, Gerber B.The carrot, not the stick:appetitive rather than aversive gustatory stimuli support associative olfactory learning in individually assayed Drosophila larvae.Journal of Comparative Physiology A,2005,191(3): 265–279. |
| [14] | Cunningham J P, Zalucki M P, West S A.Learning in Helicoverpa armigera (Lepidoptera:Noctuidae):a new look at the behaviour and control of a polyphagous pest.Bulletin of Entomological Research,1999,89(3): 201–207. |
| [15] | 李为争, 王琼, 李慧玲, 王珏, 李洋洋, 郭线茹, 原国辉.棉铃虫成虫对两种关键花香气味的联系性学习.生态学报,2015,35(11): |
| [16] | Becher P G, Guerin P M.Oriented responses of grapevine moth larvae Lobesia botrana to volatiles from host plants and an artificial diet on a locomotion compensator.Journal of Insect Physiology,2009,55(4): 384–393. |
| [17] | Perkins L E, Cribb B W, Hanan J, Zalucki M P.The role of two plant-derived volatiles in the foraging movement of 1st instar Helicoverpa armigera (Hübner):time to stop and smell the flowers.Arthropod-Plant Interactions,2009,3(3): 173–179. |
| [18] | Cunningham J P, Lange C L, Walter G H, Zalucki M P.Host location behaviour in the desert caterpillar, Heliothis punctifera.Entomologia Experimentalis et Applicata,2011,141(1): 1–7. |
| [19] | 李慧玲, 原国辉, 胡晶晶, 李洋洋, 郭线茹, 李为争.寄主植物轮换饲养和次生代谢物交叉涂布对棉铃虫取食的影响.生态学报,2014,34(24): 7421–7427. |
| [20] | Raguso R A, Levin R A, Foose S E, Holmberg M W, McDade L A.Fragrance chemistry, nocturnal rhythms and pollination "syndromes" in Nicotiana.Phytochemistry,2003,63(3): 265–284. |
| [21] | Sun J G, Huang L Q, Wang C Z.Electrophysiological and behavioral responses of Helicoverpa assulta (Lepidoptera:Noctuidae) to tobacco volatiles.Arthropod-Plant Interactions,2012,6(3): 375–384. |
| [22] | Skiri H T, Stranden M, Sandoz J C, Menzel R, Mustaparta H.Associative learning of plant odorants activating the same or different receptor neurones in the moth Heliothis virescens.The Journal of Experimental Biology,2005,208(4): 787–796. |
| [23] | Daly K C, Carrell L A, Mwilaria E.Characterizing psychophysical measures of discrimination thresholds and the effects of concentration on discrimination learning in the moth Manduca sexta.Chemical Senses,2008,33(1): 95–106. |
| [24] | Stranden M, Røstelien T, Liblikas I, Almaas T J, Borg-Karlson A K, Mustaparta H.Receptor neurones in three heliothine moths responding to floral and inducible plant volatiles.Chemoecology,2003,13(3): 143–154. |
| [25] | Fan R J, Anderson P, Hansson B S. Behavioural analysis of olfactory conditioning in the moth Spodoptera littoralis (Boisd.) (Lepidoptera:Noctuidae). The Journal of Experimental Biology, 1997, 200(23):2969-2976. |
| [26] | Fan R J, Hansson B S.Olfactory discrimination conditioning in the moth Spodoptera littoralis.Physiology & Behavior,2001,72(1/2): 159–165. |
| [27] | Daly K C, Smith B H.Associative olfactory learning in the moth Manduca sexta.The Journal of Experimental Biology,2000,203(13): 2025–2038. |
| [28] | 李为争, 李慧玲, 王珏, 郭线茹, 游秀峰, 原国辉.蛾类花香型广谱引诱剂配方的均匀设计.中国农学通报,2014,30(4): 304–311. |
| [29] | 江生. 小麦麸皮不同提取物的分析和抗氧化活性研究[D]. 镇江:江苏大学, 2009. |
| [30] | 郭钰, 曾玲, 梁广文.长头谷盗对6种寄主谷粉及其挥发物的行为反应.环境昆虫学报,2012,34(4): 432–440. |
| [31] | 耿胜荣, 夏和舟, 鉏晓艳, 陈玉霞, 叶丽秀, 熊光权. 顶空固相微萃取-气相色谱-质谱联用法分析酵母浸膏辐照挥发性成分. 食品科学, 2014, 35(6):55-59. |
| [32] | 郭予元. 棉铃虫的研究. 北京: 中国农业出版社, 1998. |
| [33] | Traynier R M M.Visual learning in assays of sinigrin solution as an oviposition releaser for the cabbage butterfly, Pieris rapae.Entomologia Experimentalis et Applicata,1986,40(1): 25–33. |
| [34] | Matsumoto Y, Mizunami M.Formation of long-term olfactory memory in the cricket Gryllus bimaculatus.Chemical Senses,2005,30(S1): i299–i300. |
| [35] | Decker S, McConnaughey S, Page T L.Circadian regulation of insect olfactory learning.Proceedings of the National Academy of Sciences of the United States of America,2007,104(40): 15905–15910. |
| [36] | 李为争, 付国需, 王英慧, 原国辉, 张元臣, 柴晓乐.棉铃虫幼虫对人类呈味物质的取食反应.生态学报,2010,30(21): 5709–5715. |
| [37] | Hartlieb E.Olfactory conditioning in the moth Heliothis virescens.Naturwissenschaften,1996,83(2): 87–88. |