 2016, Vol. 36
2016, Vol. 36文章信息
- 沈亮, 徐荣, 刘赛, 徐常青, 贺宁, 刘同宁, 陈君
- SHEN Liang, XU Rong, LIU Sai, XU Changqing, HE Ning, LIU Tongning, CHEN Jun.
- 肉苁蓉寄主梭梭根际土壤微生物种类及群落结构特征
- Characteristics of microbial community structure in rhizosphere soil of Haloxylon ammodendron
- 生态学报[J]. 2016, 36(13): 3933-3942
- Acta Ecologica Sinica[J]. 2016, 36(13): 3933-3942
- http://dx.doi.org/10.5846/stxb201410212063
-
文章历史
- 收稿日期: 2014-10-21
- 网络出版日期: 2015-10-30





2. 河南理工大学资源环境学院, 焦作 454003;
3. 宁夏永宁县本草苁蓉种植基地, 银川 750100
2. Institute of Resource and Environment, Henan Polytechnic University, Jiaozuo 454003, China;
3. Ningxia Yongning Plantation of Herba Cistanche, Yinchuan 750100, China
肉苁蓉为列当科肉苁蓉属多年生寄生性草本植物,专性寄生于苋科藜亚科梭梭属植物梭梭Haloxylon ammodendron (C. A. Mey.) Bunge根部[1]。肉苁蓉为我国传统名贵中药材,具有补肾阳、益精血、润肠通便等功效[2],有"沙漠人参"之美誉。随着人们医疗保健意识的增强,补益类中药需求量猛增,其野生资源蕴藏量已不能满足市场需求。国家明令禁止采挖野生肉苁蓉,鼓励通过人工种植满足市场需求[1]。肉苁蓉种子在没有梭梭根系分泌物的作用下,难以萌发及完成寄生生活史[3];目前尚未鉴定出梭梭根系产生的刺激肉苁蓉种子萌发的关键物质[4-5]。由于肉苁蓉和梭梭的寄生机理不清,导致人工种植肉苁蓉的寄生率不高,产量不稳定。近年来,有研究表明寄主植物根系分泌物除了刺激寄生植物种子萌发外,还会促使寄主植物根际土壤微生物繁殖。根际微生物不仅在根系分泌物化学成分转化和分解方面发挥作用;而且在植物根际快速繁殖的AM真菌还增大了寄主与土壤的接触面积,进而促进寄主对养分的吸收和根系分泌物的释放[6-7],最终促使寄生植物寄生率的提高。此外,环境因子中的土壤养分、水分、温度和湿度等因子也可引起根际土壤微生物种类及结构的变化[8]。梭梭根际土壤微生物是否参与了肉苁蓉的寄生过程以及不同季节梭梭根际土壤微生物的变化是否对肉苁蓉的寄生产生影响,目前未见相关报道。
对土壤微生物群落结构的研究通常采用传统的平板培养法,但该方法成本高、工作量大,能够培养分离的微生物只占总数的0.1%-10%,不能够全面反映土壤微生物群落结构的信息。现代研究土壤微生物的方法主要有磷脂脂肪酸法(PLFA)、BIOLOG法、变性梯度凝胶电泳法(DGGE)等[9-10]。其中PLFA是近几年发展起来的研究土壤微生物群落结构的新方法,它能够分析微生物群落的生物量和群落结构变化,主要用于揭示植物栽培及季节变化等诸多因素对土壤微生物群落结构变化的影响[11]。Ramsey等[12]研究表明PLFA也能够表征Biolog和DGGE方法检测出的土壤微生物群落结构变化,在土壤微生物检测领域具有一定优势。因此,本研究拟采用PLFA法及常规AM真菌鉴定法[13],对梭梭5-8月份生长季节的根际土壤微生物种类及群落结构特征进行分析,结合环境因子探讨梭梭根际土壤微生物在肉苁蓉寄生过程中的作用,研究结果将为解析肉苁蓉寄生机理及指导肉苁蓉人工种植提供参考。
1 材料与方法 1.1 梭梭根际土及对照土样采集与处理试验设在宁夏回族自治区永宁县本草苁蓉种植基地,该基地地理坐标为东经106°40'、北纬38°15',海拔1125 m。肉苁蓉种子来自内蒙古阿拉善盟右旗野生肉苁蓉植株。4年生寄主梭梭来自宁夏肉苁蓉种植基地,梭梭高1-2 m,行距3 m,株距1 m。
5-8月份梭梭快速生长期,在种植基地选择一块长势一致、土壤质地均一、近2年内未受干扰的梭梭种植田,面积为20 m×20 m。调查期间该样地杜绝人为干扰,每月初在试验田按照"S"形取样法,收集15株40-50 cm深处的梭梭根际土。采用抖落法收集梭梭毛细根周围0.5 cm内粘连的根际土壤1.00 kg;供试土壤为沙壤土,除去样品中植物残体和碎石,过2 mm筛后分成2部分,一部分土样冷冻干燥后保存在-80℃,用于PLFA分析;另一部分土样风干用于土壤基本理化性状测定。对照样品为5月份采集的试验地50 cm深处的非梭梭根际土1份。
1.2 土壤微生物种类及群落结构分析 1.2.1 PLFA分析PLFA的提取过程参考Frostegrd等方法[14]。提取过程分成4个步骤:脂肪酸释放与甲酯化、溶液中和、脂肪酸萃取、脂肪酸溶解。PLFAs成份测定参考Dong等方法[15]进行,采用美国Agilent 6890气相色谱仪、Agilent 5977质谱联用仪、DB-5 MS色谱柱(30 m×250 μm×0.25 μm)(安捷伦科技有限公司)等仪器进行测试。
PLFA命名及含量计算:采取通用的ω命名系统,前缀a,i和cy分别表示反式,顺式和环状支链构型。以18:2ω6,9为例,18表示主链C原子数目,冒号":"后数字代表不饱和度,ω后数字代表双键所在位置。表征细菌、真菌、放线菌、革兰氏阳性细菌、革兰氏阴性细菌的磷脂脂肪酸标记参考Kimura等方法[16-18]。磷脂脂肪酸含量测定用峰面积法,计算公式为

|
(1) |
式中,PPLFA和PSTD分别是样品和标准物质的峰值面积,S为内标物质浓度(ng/μL),V为样品测试体积(1 μL),R为分取倍数(1/1000),D为稀释倍数(10倍),W为干土质量(g)。
1.2.2 AM真菌分离与鉴定采用湿筛倾注-蔗糖离心法[19]分离土壤孢子,在体视显微镜下观察孢子大小、颜色、菌丝特征、孢子形态等指标。根据《VA菌根真菌鉴定手册》[20]进行分类描述和种属检索和鉴定。不同月份AM真菌孢子数量采用计数法进行。
1.3 肉苁蓉田间接种寄生及种子萌发试验在距梭梭主根40 cm处挖深50 cm、宽20 cm接种坑,每坑接肉苁蓉种子40-60粒,共接种30株梭梭。首次接种时间为5月14日,1个月后调查寄生率,每次调查后重新接种,即分别得到6月15日、7月15日和8月15日的接种寄生率:
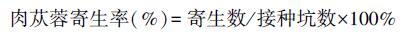
|
(2) |
取每月采集的土样10 g添加50%的甲醇水溶液,超声35 min,温度为47℃,震荡12 h,150r/min,离心,取提取液10 mL,过0.22 μm滤膜,做成0.2 g/m的提取液,稀释1000倍用于肉苁蓉种子萌发试验[21]。

|
(3) |
采自宁夏本草苁蓉种植基地的AR5自动气象站,设定为1 h采集1次气象因子数据,不同月份气象数据为该月气象数据的总平均值。采集的气象因子有:气温(TA)、大气相对湿度(RH)、光照有效辐射(PAR)、50 cm土壤湿度(SM50)、50 cm土壤温度(ST50),降雨量(PT)。
1.4.2 土壤养分测定土壤全氮(TN)采用凯氏定氮仪测定;有效磷(AP)采用钼锑抗比色法测定;速效钾(AK)采用原子吸收分光光度计法测定;有机质(OM)采用重铬酸钾-外加热容重法测定;pH值采用pH 300型便携式pH酸度计法测定。
1.5 数据分析结果为3次重复测定数据的平均值,采用SPSS 20.0统计软件对试验数据进行方差分析、相关分析、主成分分析,用单因素方差分析法(LSD法)检验差异显著性,利用主成分分析法(PCA)进行群落结构比较,结果为所测指标的平均值±标准差。
2 结果与分析 2.1 不同月份梭梭土壤微生物磷脂脂肪酸含量比较由表 1得知,不同月份梭梭根际土壤微生物PLFA种类及含量差异显著。5-8月份土壤微生物总磷脂脂肪酸的含量为6月>7月>5月>8月,说明随着月份变化,梭梭根际土壤微生物的总磷脂脂肪酸含量呈现先上升后下降的趋势。细菌、革兰氏阴性细菌、真菌、放线菌以及需氧菌5-7月份的PLFA含量均高于8月份,且均呈现先上升后下降的趋势。其中真菌的PLFA含量范围为1.70-2.27 nmol/g,以7月份含量最高,8月份含量较低;AM真菌含量范围为0.24-0.33 nmol/g,以6月份含量最高,8月份含量最低。此外,对照土CK土壤微生物的PLFA含量均显著低于不同月份梭梭根际土壤微生物的PLFA值。因此,不同月份环境条件的改变能够引起梭梭根际土壤微生物的结构变化。
| 微生物种群Microbial population | 对照 CK | 5月 May | 6月 June | 7月 July | 8月 August |
| 总磷脂脂肪酸Total PLFA | 3.84±0.42c | 4.90±0.51b | 6.00±0.40a | 5.15±0.40ab | 3.62±0.66c |
| 饱和脂肪酸Normal saturated fatty acids | 2.22±0.12b | 2.53±1.08a | 3.10±0.10a | 2.79±0.62a | 0.40±0.36b |
| 单不饱和脂肪酸Monounsaturated fatty acids | 0.19±0.18b | 0.32±0.15a | 0.26±0.11a | 0.34±0.11a | - |
| 细菌Bacterial biomass | 3.07±1.53a | 4.45±1.23a | 4.44±0.55a | 4.49±0.13a | 4.04±1.53a |
| 革兰氏阳性细菌Gram+ | 0.65±0.16c | 1.15±0.07a | 0.74±0.12bc | 0.95±0.03ab | 0.55±0.16c |
| 革兰氏阴性细菌Gram- | 0.35±0.12b | 0.86±0.31a | 0.45±0.32a | 0.66±0.04a | 0.16±0.02b |
| 革兰氏阳性/阴性细菌Gram+/Gram- | 1.23±0.17a | 1.73±0.68b | 1.35±0.37b | 1.43±0.06b | 3.58±0.07a |
| 真菌Fungi | 1.61±0.16a | 1.78±0.28a | 2.07±0.32a | 2.27±0.13a | 1.70±0.27a |
| AM真菌AM fungi | 0.21±0.04a | 0.32±0.15a | 0.33±0.11a | 0.26±0.06a | 0.24±0.08a |
| 真菌/细菌Fungal/Bacterial biomass | 0.39±0.08a | 0.49±0.06a | 0.54±0.05a | 0.49±0.06a | 0.58±0.04a |
| 放线菌Actinomycetes | 0.44±0.06a | 0.70±0.07a | 0.50±0.29a | 0.64±0.04a | 0.64±0.05a |
| 需氧菌Aerobic bacteria | 1.21±0.04a | 1.86±0.47a | 1.73±0.58a | 2.20±0.14a | 1.51±0.23a |
| 厌氧菌Anaerobic bacteria | - | 0.19±0.06c | 0.22±0.04a | 0.20±0.00b | - |
| 每组同列的不同字母表示差异显著性水平,小写字母为5%差异显著水平 | |||||
对不同月份梭梭根际土壤微生物的磷脂脂肪酸进行主成分分析(图 1a)。结果表明,第一主成分对总PLFA数据变异的贡献率是90.12%,第二主成分对总PLFA数据变异的贡献率是9.21%,累计贡献率达到99.33%。因此,两个主成分可以全面反映研究区域微生物群落的情况。从图 1a可看出,CK及5-8月份的梭梭根际土壤微生物均与第一主成分高度正相关;大部分5月份、7月份及8月份的梭梭根际土壤微生物与第二主成分高度正相关,而大部分的6月份样品与PC2呈负相关。除8月份梭梭根际土壤微生物外,5-7月份梭梭样品相距较近。对照处理CK与6月份的样品距离较近,说明二者结构较为相似。总之,环境因子对不同月份的土壤微生物群落结构变化影响极为明显,5-7月份梭梭根际土壤微生物结构相似,三者均与8月份样品差异显著。

|
| 图 1 梭梭根际土壤微生物群落PLFA主成分分析(a);梭梭根际土壤微生物群落PLFA载荷因子贡献(b) Fig. 1 PCA of PLFA profiles from soil microbial communities of Haloxylon ammodendron (a); Eigenvector loading of PLFA contributing to soil microbial communities ordination pattern of Haloxylon ammodendron (b) |
通过各种土壤微生物在主成分上的载荷因子分析表明(图 1b),代表细菌、厌氧菌、革兰氏阴性细菌和放线菌的磷脂脂肪酸在主成分一(65.75%)上的载荷值较高,主成分一是它们的代表因子,而代表真菌及需氧菌的磷脂脂肪酸在主成分一上均具有较低的载荷值。说明梭梭根际土壤的细菌、厌氧菌、革兰氏阴性细菌和放线菌的数量较多,而真菌及需氧菌的数量相对较少。代表革兰氏阳性细菌及AM真菌的磷脂脂肪酸在主成分二(25.43%)上有较高的载荷值,而代表真菌的磷脂脂肪酸在主成分二上的含量较低。说明革兰氏阴性细菌及AM真菌的数量较多,而真菌的含量较低。综合分析得出,随着月份上升,土壤中的细菌及放线菌类微生物数量不断增多,而真菌类微生物数量较少。
2.3 不同月份梭梭根际AM真菌种类及数量分析由图 2得知,梭梭根际土壤共分离鉴定出4属35种AM真菌。其中球囊霉属(Glomus)22种、无梗囊霉属(Acaulospora)7种、多孢囊霉属(Diversispora)3种、巨孢囊霉属(Gigaspora)3种。黑球囊霉(G. melanosporum)和双网无梗囊霉(A. bireticulata)为其中主要的优势菌种,且在各土壤样品中均有分布。通过计数法分析得知5-8月份梭梭根际土壤AM真菌孢子数量随气温升高而呈现上升趋势。5月份土壤样品中孢子数量(17.5个/g)最低,6月份(31.0个/g)和8月份(35.5个/g)AM真菌孢子密度最高。

|
| 图 2 梭梭根际土AM真菌孢子光镜照片(×400) Fig. 2 AM fungal species under the optical microscope (×400) |
肉苁蓉种子萌发及接种寄生试验表明不同月份的肉苁蓉田间接种寄生率差异较大(表 2)。与对照相比,测试6月份的梭梭根际土壤提取液得到的肉苁蓉种子萌发率最高,而5月份的种子萌发率最低;6月份的肉苁蓉田间接种寄生率最高,而8月份的肉苁蓉田间接种寄生率最低。
| 肉苁蓉Cistanche deserticola | 对照CK | 5月May | 6月June | 7月July | 8月August |
| 种子萌发率Germination rate of seed | 21.65±1.54d | 37.83±8.86c | 65.94±2.77a | 49.61±6.76b | 48.15±7.88b |
| 接种寄生率Parasitism rate | - | - | 59.19±27.04a | 46.75±27.03b | 45.56±24.08b |
本试验基地地处我国温带大陆性气候区,不同月份的气候因子差异较大,且随着气候因子的变化,梭梭根际土壤的基本养分也发生了显著变化(表 3,表 4)。土壤养分及气候因子均可以引起根际土壤微生物种类及结构的变化。相关分析表明(表 5),厌氧菌与气温的相关性达到负显著性水平(P<0.05),需氧菌与50 cm深土壤湿度间的相关性达到正显著性水平(P<0.05),真菌与土壤温度之间的相关性达到正显著性水平(P<0.05)。由此说明,梭梭根际土壤微生物种类及含量主要受温度和湿度影响。梭梭根际微生物与土壤养分间的相关分析表明,革兰氏阳性细菌与各土壤养分的相关性均达到负显著性水平(P<0.05),真菌与土壤全氮、有效磷和有机质呈正相关性,其它微生物与土壤养分各指标不相关。这说明革兰氏阳性细菌的数量随着土壤养分的上升而上升,真菌随着土壤养分的增加而上升。肉苁蓉寄生率与环境因子的相关分析表明,肉苁蓉种子萌发率及田间寄生率与50 cm深土壤温度呈现极显著负相关性(P<0.01),与土壤全氮呈现显著负相关(P<0.05),与有效磷、速效钾和有机质呈现极显著负相关(P<0.01)。这说明随着50 cm深土壤温度的增大,肉苁蓉的寄生率逐渐降低;随着土壤养分的升高,肉苁蓉的寄生率逐渐下降。
| 月份Months | 气温Temperature/℃ | 大气相对湿度Relative humidity of atmosphere /% | 光照有效辐射Effectively of light radiation /lx | 50 cm土壤湿度Soil moisture of 50 cm /% | 50 cm土壤温度Soil temperature of 50 cm /℃ | 降雨量Precipitation/mm |
| 5 | 18.86±1.57b | 29.07±0.96a | 322.50±34.50a | 8.27±8.60ab | 17.20±1.42a | 0.04±0.04a |
| 6 | 22.35±1.47a | 27.12±1.56a | 321.42±41.12a | 17.66±0.96a | 20.65±0.12b | 0.03±0.05a |
| 7 | 23.87±0.36a | 21.87±0.21b | 320.85±19.43a | 9.67±1.80ab | 23.50±0.35c | 0.02±0.02a |
| 8 | 24.16±2.14a | 29.05±3.17a | 320.75±23.42a | 5.97±8.30b | 24.48±0.55c | 0.01±0.01a |
| n=30,每组同列的不同字母表示差异显著性水平;小写字母为5%差异显著水平 | ||||||
| 月份Months | 全氮Total N/(g/kg) | 有效磷Available P/(mg/kg) | 速效钾Readily available K/ (mg/kg) | 有机质Organic matter/ (g/kg) | pH |
| 5 | 0.37±0.02c | 7.90±0.54c | 36.10±2.12c | 3.91±0.23c | 8.76±0.79d |
| 6 | 0.35±0.07d | 7.10±0.63d | 35.30±1.27d | 3.24±0.26d | 9.26±0.83b |
| 7 | 0.40±0.04b | 9.70±0.82a | 64.40±4.37b | 4.05±0.28a | 9.13±0.61c |
| 8 | 0.46±0.05a | 8.60±0.73b | 66.20±4.72a | 3.95±0.31b | 9.32±0.52a |
| 每组同列的不同字母表示差异显著性水平;小写字母为5%差异显著水平 | |||||
| 微生物种群Microbial population | TA | RH | PAR | SM50 | ST50 | PT | TN | AP | AK | OM | pH |
| 细菌Total bacteria | -0.75 | 0.58 | -0.59 | -0.52 | -0.68 | 0.61 | -0.72 | -0.68 | -0.66 | -0.69 | -0.65 |
| 革兰氏阳性细菌Gram+ | -0.67 | 0.54 | 0.03 | -0.53 | -0.69 | -0.23 | -0.88* | -0.85* | -0.84* | -0.86* | -0.83* |
| 革兰氏阳性细菌Gram- | 0.40 | -0.36 | 0.80 | -0.10 | 0.31 | -0.63 | -0.23 | -0.28 | -0.30 | -0.27 | -0.31 |
| 需氧菌Aerobic bacteria | 0.62 | -0.41 | 0.33 | 0.85* | 0.60 | -0.16 | 0.52 | 0.47 | 0.45 | 0.48 | 0.44 |
| 厌氧菌Anaerobic bacteria | -0.81* | 0.64 | -0.51 | -0.66 | -0.78 | 0.26 | -0.59 | -0.54 | -0.52 | -0.55 | -0.51 |
| 真菌Total fungi | 0.80 | -0.70 | 0.17 | 0.61 | 0.83* | -0.10 | 0.83* | 0.80* | 0.79 | 0.81* | 0.78 |
| AM真菌AM fungi | -0.12 | 0.34 | 0.33 | 0.64 | -0.17 | -0.42 | -0.40 | -0.35 | -0.32 | -0.36 | -0.31 |
| 放线菌Actinomycetes | -0.04 | 0.07 | 0.50 | -0.68 | -0.18 | -0.37 | -0.27 | -0.32 | -0.34 | -0.31 | -0.35 |
| 肉苁蓉种子萌发率Germination | -0.33 | 0.49 | 0.29 | 0.75 | -0.98** | 0.41 | -0.85* | -0.92** | -0.99** | -0.98** | 0.46 |
| 肉苁蓉寄生率Parasitism rate | -0.28 | 0.54 | 0.37 | 0.75 | -0.99** | 0.39 | -0.85* | -0.93** | -0.99** | -0.99** | 0.48 |
| *表示P<0.05水平显著相关,**表示P<0.01水平极显著相关;TA:气温 air temperature;RH:大气相对湿度 relative humidity;PAR:光照有效辐射 effective radiation of light;SM50:50 cm土壤湿度 soil moisture;ST50:50 cm土壤温度 soil temperature;PT:降雨量 precipitation;TN:全氮 total nitrogen;AP:有效磷 available phosphorus;AK:速效钾 available potassium;OM:有机质 organic matter | |||||||||||
土壤微生物主要包括土壤中的细菌、真菌、放线菌及藻类。本研究中除对照CK及8月份的样品没有检测到厌氧菌外,其他不同月份的梭梭根际土壤中均检测到了细菌、真菌、AM真菌及放线菌。其中以表征细菌的PLFA所占的比例最高,其次是真菌、放线菌占的比例最低,这与以往的研究结果相似[22]。一般来说,土壤中细菌数量最多,其次为真菌,放线菌数量最少。随着环境变化,土壤微生物区系种类也会发生显著改变[23]。细菌含量较高因其结构简单、繁殖快、抗逆性强、可以生存于湿润及透气性不好的环境[24];真菌含量相对较少是因其只适宜于生长在通气良好的环境中;放线菌喜热、耐旱,但其竞争力较弱[25-26],种群较小。梭梭根际土壤微生物的PLFA含量显著高于对照土CK,说明梭梭根系分泌物可以促进其根际土壤微生物生长。罗倩等[27]采用平板培养法研究了梭梭根际和非根际土壤微生物的数量及群落结构特征,结果表明梭梭具有较强的根际正效应,且根际微生物也显著促进梭梭的生长及其对土壤养分的吸收。通常植物光合产物有4%-7%通过分泌作用进入土壤,在根系分泌物的作用下,根际微生物得到快速生长,其种类及数量远高于非根际土壤微生物[28]。
3.2 环境因子对梭梭根际土壤微生物的影响研究表明梭梭在不同生长时期,其根际土壤微生物的数量、种类及优势种等均存在一定差异,这与不同环境条件下的梭梭根系分泌物种类和含量有关[29]。梭梭一般为4月底抽出新芽,5月中旬开花,6月初进入生长旺盛期,7-8月发育中的种子进入休眠期,9-10月种子进入成熟期。研究表明5-7月份的梭梭根系微生物种类及含量高于8月份样品。这与5-7月气温及湿度适宜,梭梭生长活跃,大量产生根系分泌物促进其根际土壤微生物的生长有关;8月份天气炎热,梭梭生长缓慢进入夏季休眠阶段导致其根系分泌物减少,使得其根际土壤微生物的活动减弱。陈子英等[30]研究表明冬小麦由幼苗期到返青期间的根际微生物的数量最高,而返青后迅速减少,这与小麦返青期间产生的大量根系分泌物有关。
相关分析表明真菌与土壤温度间的相关性达到正显著性水平,说明梭梭根际真菌的数量随着温度上升逐渐增大。5-7月份的梭梭根际细菌、真菌、放线菌等微生物PLFA含量显著高于8月份,可能与5-7月份采样点降雨量、大气相对湿度和土壤湿度等条件较好,使得土壤微生物快速生长有关;土壤温湿度较高也促使梭梭毛细根生长较快,使其根系分泌物增加,进而促进了土壤微生物种类及数量的增加。肉苁蓉寄生率与50 cm深土壤温度呈显著负相关,说明随着土壤温度升高,肉苁蓉的寄生率逐渐降低。因此,结合真菌与土壤温度的相关分析,得出肉苁蓉的最适寄生温度应该在6月份。肉苁蓉的寄生率与土壤养分显著负相关,说明在土壤养分含量较低的时候肉苁蓉寄生率较高,这可能与土壤贫瘠引起其根际土壤微生物的大量繁殖,促进了肉苁蓉的寄生有关。该试验基地的平均pH值为9.12,较高的pH值与该基地位于偏碱性水质的黄河附近有关,这与梭梭耐盐碱的生长特性也一致。大多数微生物适宜在中性或微碱性环境中生存,土壤pH值降低会减弱微生物的活动并影响到其生长状况[31],该基地6月份较高的pH值及适宜的气候可能促进梭梭根际土壤微生物大量繁殖,这对梭梭根系分泌物的释放具有一定的促进作用。
3.3 梭梭根际土壤微生物变化对肉苁蓉寄生的影响近年来,肉苁蓉与梭梭的寄生机理已成为肉苁蓉研究领域的热点。李天然等[3]研究表明肉苁蓉种子只有在梭梭根系分泌物存在的情况下才能萌发,且只在多年生的梭梭根际才能完成寄生生活史。梭梭是多年生旱生小乔木,一些较为成熟的寄生性草本植物根系分泌物研究方法,如独角金(Strigaasiatica)、百脉根(Lotusjaponicus)和菟丝子(Cuscuta chinensis)等水溶液培养方法很难用于梭梭根系分泌物的研究,且多年生、沙生性的生物特征也使得梭梭很难在实验室开展研究[7, 32-33];田间收集的梭梭根系分泌物因受土壤微生物、土壤小动物及植物残体等因素影响难以有效控制。因此,目前还未见有关从梭梭根系分泌物中得到刺激肉苁蓉种子萌发的相关物质报道。有研究表明植物根际土壤微生物可以通过改变寄主植物根际营养状况和植物激素含量来改变植物体内生理生化过程,从而影响寄主根系分泌物种类和数量[34]。本研究首次对不同季节的梭梭根际土壤微生物种类及群落结构特征进行分析,结果表明肉苁蓉田间寄生率与其根际土壤微生物的结构变化有一定相关性,二者的最高值均出现在6月份。Akiyama等[7]研究表明,百脉根根系分泌物中含有促进产生菌丝分支的化学物质5-deoxystrigol,在很低的浓度时就可以刺激其根系周围菌根真菌产生大量分支。因此,寄生植物在寄生过程中与其寄主根际土壤微生物的关系密切。有研究表明珠状巨孢囊霉(G. margarita)通过在寄主百脉根毛细根处定植及产生分支,增大百脉根与土壤的接触面积,从而促进其对土壤养分的吸收和寄生关键物质的分泌,本研究也从梭梭根际土壤中发现了可能参与寄生的巨孢囊霉属AM真菌[7]。因此,梭梭根际土壤微生物可能参与肉苁蓉的寄生过程。
由于田间试验影响因素较多,梭梭根际微生物是否参与肉苁蓉的寄生,还需要进一步证实。根据梭梭的生长特性及前人研究方法,已经初步摸索出了一套严谨的田间梭梭根系分泌物和根际土壤微生物收集及分析方法,相关研究工作正在进行中。由于磷脂不能作为细胞的贮存物质,一旦生物细胞死亡,其中的磷脂化合物就会迅速分解,使得PLFA的测试结果不能准确的代表土壤微生物的真实信息。此外,该方法只能从土壤微生物属的水平上进行鉴定,不能到达种的水平。因此,后续用于梭梭根际特定微生物类群的分析还需要利用PCR-DGGE等方法进行。
4 结论(1) 5-7月份梭梭根际土壤微生物种类及PLFA含量显著高于8月份,以6月份总PLFA及AM真菌PLFA含量最高;采用湿筛倾注-蔗糖离心法,从梭梭根际土壤共鉴定出4属35种AM真菌。它们分别为:球囊霉属22种、无梗囊霉属7种、多孢囊霉属3种、巨孢囊霉属3种。其中以黑球囊霉和双网无梗囊霉为优势种群,以6月份和8月份的梭梭根际土壤AM真菌孢子数量最高,且发现了可能参与寄生的巨孢囊霉属AM真菌。
(2) 梭梭根际土壤微生物群落结构的变化受气候因子及土壤养分的显著影响;与寄生有关的真菌数量与50 cm深土壤温度呈显著正相关;肉苁蓉寄生率与50 cm深土壤温度及土壤养分呈显著负相关。
(3) 6月份梭梭根际土壤提取液得到的肉苁蓉种子萌发率和肉苁蓉田间接种寄生率在5-8月份中均最高,这与6月份梭梭根际土壤微生物的种类及PLFA含量较高显著相关。根据以上结果,本研究推测梭梭根际土壤微生物可能参与了肉苁蓉的寄生过程。
致谢: 孔垂华教授、董红云博士、夏志超博士、刘源经理及常玉成农艺师在实验材料收集、样品测试及分析工作中给予帮助,特此致谢。| [1] | 贾志清, 鲁琦. 梭梭. 北京: 中国环境科学出版社, 20051–30. |
| [2] | 国家药典委员会. 中华人民共和国药典(I部). 北京: 中国医药科技出版社, 201025–25. |
| [3] | 李天然, 许月英, 戈建新, 徐梅英.肉苁蓉(Cistanche deserticola Ma)种子的萌发及与寄主梭梭(Haloxylon ammodendron Bunge)的关系.内蒙古大学学报:自然科学版,1989,20(3): 395–400. |
| [4] | 张汝民, 张丹, 白静, 陈宏伟, 高岩.不同苗龄梭梭根系分泌物组分分析.西北植物学报,2006,26(10): 2150–2154. |
| [5] | 陈虞超, 李苗, 陈晓军, 张丽, 宋玉霞.外源信号物质对肉苁蓉种子萌发与吸器形成的影响.植物生理学报,2012,48(3): 260–264. |
| [6] | Akiyama K, Hayashi H.Strigolactones:Chemical signals for fungal symbionts and parasitic weeds in plant roots.Annals of Botany,2006,97(6): 925–931. |
| [7] | Akiyama K, Matsuzaki K, Hayashi H.Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi.Nature,2005,435(7043): 824–827. |
| [8] | 展小云, 吴冬秀, 张琳, 张灿娟, 周双喜, 杨云霞.小叶锦鸡儿根际微生物群落功能多样性对环境变化的响应.生态学报,2010,30(12): 3087–3097. |
| [9] | 王玲, 黄世文, 刘连盟, 黄雯雯.三种研究农田土壤微生物多样性方法的比较.科技通报,2009,25(5): 588–592. |
| [10] | Insam H.Developments in soil microbiology since the mid 1960s.Geoderma,2001,100(3/4): 389–402. |
| [11] | 颜慧, 蔡祖聪, 钟文辉.磷脂脂肪酸分析方法及其在土壤微生物多样性研究中的应用.土壤学报,2006,43(5): 851–859. |
| [12] | Ramsey P W, Matthias C R, Kevin P F, Holben W E, Gannon J E.Choice of methods for soil microbial community analysis:PLFA maximizes power compared to CLPP and PCR-based approaches.Pedobiologia,2006,50(3): 275–280. |
| [13] | Smith S E, Read D J. Mycorrhizal Symbiosis.3rd ed. Cambridge UK: Academic Press, 2008. |
| [14] | Frostegård A, Tunlid A, Bååth E.Phospholipid fatty acid composition, biomass, and activity of microbial communities from two soil types experimentally exposed to different heavy metals.Applied and Environmental Microbiology,1993,59(11): 3605–3617. |
| [15] | Dong H Y, Kong C H, Wang P, Huang Q L.Temporal variation of soil friedelin and microbial community under different land uses in a long-term agroecosystem.Soil Biology and Biochemistry,2014,69 275–281. |
| [16] | Kimura M, Asakawa S.Comparison of community structures of microbiota at main habitats in rice field ecosystems based on phospholipid fatty acid analysis.Biology and Fertility of Soils,2006,43(1): 20–29. |
| [17] | Zelles L, Bai Q Y, Rackwitz R, Chadwick D, Beese F.Determination of phospholipid and lipopolysaccharide-derived fatty acids as an estimate of microbial biomass and community structures in soils.Biology and Fertility of Soils,1995,19(2/3): 115–123. |
| [18] | 顾美英, 徐万里, 茆军, 张志东, 唐光木, 葛春辉.新疆绿洲农田不同连作年限棉花根际土壤微生物群落多样性.生态学报,2012,32(10): 3031–3040. |
| [19] | Daniels B A, Skipper H D. Methods for the recovery and quantitative estimation of propagules from soil. USA: The American Phytopathology Society Press, 1982. |
| [20] | 刘润进, 李晓林. 丛枝菌根及其应用. 北京: 科学出版社, 2000190–194. |
| [21] | 于晶, 周峰, 徐荣, 陈君, 马维思, 徐江, 刘同宁.赤霉素对肉苁蓉种子萌发的影响研究.中国种业,2010,10 59–61. |
| [22] | 胡婵娟, 郭雷, 刘国华. 黄土丘陵沟壑区不同植被恢复格局下土壤微生物群落结构. 生态学报, 2014, 34(11):2986-2995. |
| [23] | 李元, 杨济龙, 王勋陵, 胡之德.紫外辐射增加对春小麦根际土壤微生物种群数量的影响.中国环境科学,1999,19(2): 157–160. |
| [24] | Herndl G J, Müller-Niklas G, Frick J.Major role of ultraviolet-B in controlling bacterioplankton growth in the surface layer of the ocean.Nature,1993,361(6414): 717–719. |
| [25] | Amann R, Ludwig W.Ribosomal RNA-targeted nucleic acid probes for studies in microbial ecology.FEMS Microbiology Review,2000,24(5): 555–565. |
| [26] | Tabacchioni S, Chiarini L, Bevivino A, Cantale C, Dalmastri C.Bias caused by using different isolation media for assessing the genetic diversity of a natural microbial population.Microbial Ecology,2000,40(3): 169–176. |
| [27] | 罗倩, 黄宝灵, 唐治喜, 来利明, 魏伟, 郑元润.新疆盐渍土3种植被类型土壤微生物碳源利用.应用与环境生物学报,2013,19(1): 96–104. |
| [28] | Marschner P, Rengel Z. Contributions of rhizosphere interactions to soil biological fertility//Soil Biological Fertility. Netherlands: Springer, 200781–98. |
| [29] | 张汝民, 张丹, 白静, 陈宏伟, 高岩.不同苗龄梭梭根系分泌物组分分析.西北植物学报,2006,26(10): 2150–2154. |
| [30] | 陈子英, 吴观以, 王涌清, 吴瑞武.植物根际微生物与根部分泌物的研究I. 小麦、玉米、绿豆等作物的根际微生物数量与根部分泌物中的可溶性糖和氨基酸.微生物学报,1964,10(2): 212–221. |
| [31] | Rousk J, Brookes P C, Bååth E.Contrasting soil pH effects on fungal and bacterial growth suggest functional redundancy in carbon mineralization.Applied Environmental Microbiology,2009,75(6): 1589–1596. |
| [32] | Besserer A, Puech-Pages V, Kiefer P, Gomez-Roldan V, Jauneau A, Roy S, Portais J C, Roux C, Becard G, Sejalon-Delmas N.Strigolactones stimulate arbuscular mycorrhizal fungi by activating mitochondria.PLoS Biology,2006,4(7): e226 |
| [33] | Bouwmeester H J, Matusova R, Sun Z K, Beale M H.Secondary metabolite signaling in host-parasitic plant interactions.Current Opinion in Plant Biology,2003,6(4): 358–364. |
| [34] | Graham R D.Genotypic differences in tolerance to manganese deficiency//Manganese in Soils and Plants.Netherlands:Springer,1988,33 261–276. |