 2016, Vol. 36
2016, Vol. 36文章信息
- 李家兵, 张秋婷, 张丽烟, 仝川
- LI Jiabing, ZHANG Qiuting, ZHANG Liyan, TONG Chuan.
- 闽江河口春季互花米草入侵过程对短叶茳芏沼泽土壤碳氮分布特征的影响
- Effect of Spartina alterniflora invasion sequence on soil carbon and nitrogen distribution in a Cyperus malaccensis marsh of the Min River estuary in spring
- 生态学报[J]. 2016, 36(12): 3628-3638
- Acta Ecologica Sinica[J]. 2016, 36(12): 3628-3638
- http://dx.doi.org/10.5846/stxb201510092040
-
文章历史
- 收稿日期: 2015-10-09
- 修订日期: 2016-02-25





2. 福建师范大学环境科学与工程学院, 福州 350007;
3. 福建闽江河口湿地国家级自然保护区管理处, 福州 350200;
4. 福建师范大学亚热带湿地研究中心, 福州 350007;
5. 福建师范大学湿润亚热带生态地理过程教育部重点实验室, 福州 350007;
6. 福建师范大学地理研究所, 福州 350007
2. College of Environmental Science and Engineering, Fujian Normal University, Fuzhou 350007, China;
3. Fujian Min River Estuary Wetland National Nature Reserve, Fuzhou 350200, China;
4. Research Centre of Wetlands in Subtropical Region, Fujian Normal University, Fuzhou 350007, China;
5. Fujian Province and Education of Ministry's Key Laboratory of Humid Sub-tropical Eco-geographical Process, Fujian Normal University, Fuzhou 350007, China;
6. Institute of Geography, Fujian Normal University, Fuzhou 350007, China
河口湿地是河口与海洋相互作用形成的重要生态类型,是海岸带地区响应全球气候变化和人类活动较为敏感的生态系统之一[1]。受河口水动力和海洋潮汐的双重影响,河口湿地的物理、化学及生物过程活跃,生源元素循环过程极为复杂[2]。互花米草(Spartina alterniflora)引入我国滨海湿地后,在抬高滩面、消浪护堤、促淤固沙等方面均发挥了良好作用,但是互花米草在包括长江口、闽江口等河口盐沼-半咸水沼泽湿地的迅速入侵及快速蔓延,已严重威胁到原生河口湿地生态系统的结构、过程及功能。互花米草入侵对河口滨海湿地的影响研究已有较多文献报道,包括互花米草入侵对湿地土壤碳氮养分、土壤有机碳组分及分布[3-10]、植物种类[11]、底栖生物与微生物群落[12-14]及湿地甲烷排放的影响[15-17]等方面,然而,以互花米草入侵斑块及未被入侵的背景沼泽湿地为研究对象,揭示互花米草入侵短期序列下沼泽湿地碳氮养分分布影响的对比研究还比较薄弱。
福建是我国互花米草入侵最为严重的省份之一,目前全省互花米草面积已达4166 hm2,仅次于江苏、浙江和上海[18]。闽江河口湿地是福建省面积最大、生态类型最齐全、冲淤演变最复杂的典型淤泥质海滨湿地,其主要分布的原生湿地沼泽植物主要有芦苇(Phragmites australis)、短叶茳芏(Cyperus malaccensis)和海三棱藨草(Scirpus mariqueter)等。自2002年互花米草开始入侵闽江河口鳝鱼滩湿地,2010年鳝鱼滩湿地互花米草湿地面积已达306.9 hm2 [19]。近年来互花米草开始入侵分布在鳝鱼滩湿地中西部的土著种短叶茳芏沼泽湿地并形成一些大小不一的互花米草入侵斑块。当前,关于互花米草入侵影响下闽江河口湿地的相关研究主要集中于互花米草生物量及不同治理方式对土壤有机碳及养分影响[20-21]、土壤间隙水营养盐分布[22]、植物养分累积[23]、残体分解释放动态[24-25]、温室气体通量及影响因素[26-29]以及入侵裸滩和芦苇湿地有机碳库[30-31]等方面,而关于互花米草入侵土著种短叶茳芏过程中其对不同入侵阶段湿地土壤碳氮分布特征的影响研究还比较薄弱。鉴于此,本研究选择互花米草入侵湿地、入侵边缘湿地以及未入侵短叶茳芏湿地作为研究对象,采用时空互代研究方法,探讨互花米草入侵序列下湿地土壤碳氮空间分布特征的差异。研究结果不但可揭示互花米草不同入侵阶段湿地碳氮养分的变化过程,而且还可为下一步本区互花米草的入侵治理提供重要基础数据。
1 研究区域与研究方法 1.1 研究区概况及样品采集研究区位于闽江河口区域面积最大的鳝鱼滩湿地(119°34′12″—119°41′40″E、26°00′36″—26°03′42″N)。该区域气候暖热潮湿,年均温度19.3 ℃,年降水量1380 mm左右。鳝鱼滩湿地位于琅岐岛与长乐市梅花水道之间,是闽江水流从上游搬运来的泥沙在梅花水道中淤积形成的河口浅滩,正常潮汐情况下显现于水面。湿地土壤为滨海盐土和砂土,主要植物群落为短叶茳芏群落、芦苇群落、海三棱藨草群落以及互花米草群落,其中西部高潮滩潮汐微咸水盐度约为3.79±1.35。
近2年来,在研究区鳝鱼滩中西部大面积短叶茳芏群落中零星分布有遭受互花米草入侵的斑块。为此,2014年4月中旬,选择其中最大的一个地势平坦、远离潮沟的互花米草入侵斑块(面积约900 m2)为研究样地,沿着等高线方向布设短叶茳芏群落、互花米草入侵斑块边缘以及斑块中央3个研究样点。3个样点在高潮时均被潮水淹没,退潮时均裸露,因此潮涨潮落对3个样点土壤各元素分布的影响比较一致。基于时空互代法以及前述短叶茳芏群落(简称入侵前:A),互花米草入侵斑块边缘(简称入侵进程:B)以及互花米草入侵斑块中央(简称入侵后:C)3个样点(图 1)探讨互花米草入侵序列下湿地土壤碳氮空间分布特征的差异。3个样点的土壤田间持水量分别为(44.11±3.65)%,(43.88±8.37)%和(44.98±3.75)%,地上生物量分别为3.99×102、13.45×102 g/m2和24.42×102 g/m2,植株密度分别为(130±11)、(121±10)株/m2和(116±8) 株/m2,平均高度分别为45、100 cm和150 cm。

|
| 图 1 闽江河口鳝鱼滩湿地采样点示意图 Fig. 1 Location of the sampling sits in the Shanyutan wetland of the Min River estuary |
采样时间选择在小潮日,在A、B、C采样点分别用内径为6 cm,长60 cm 的硬质PVC管沿S形分别选择4个点位取样,每个点位采集4个重复柱样(采样深度为30 cm),3个样地共采集48个。柱状样采集后,对每根柱状样按照3 cm一层进行现场分割。分割后的土壤样品立即用自封带封口避光保存。新鲜土样带回实验室挑出可见的动植物残体,石头和其余杂物,自然风干后等分成2份:一份研磨过2 mm和0.149mm土壤筛,装袋备测;另一份土样过2 mm土壤筛,装袋置于4℃冰箱中保存备测。
1.2 样品分析称取10 mg过0.149 mm筛的土壤样品,用碳氮元素分析仪(Vario MAX,德国)测定TC、TN含量。称取5 g过0.149 mm筛的土壤样品,采用25 ml的2 mol/L溶液浸提后用连续流动分析仪(SKALAR SAN++,荷兰)测定NO-3-N和NH+4-N含量。称取5 g过2 mm筛的土壤样品,加入25 mL蒸馏水,振荡30 min后离心静置澄清后用pH计(IQ150,美国)直接测定pH。电导率在采样现场使用便携式电导仪(2265FS,美国)直接测定。
1.3 指标计算土壤剖面第i层平均碳、氮元素密度(Ci)和单位面积一定深度内(j到n 层)碳、氮元素储量(Tc)采用下式计算:
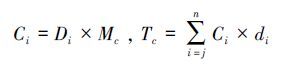
|
式中,Ci为碳、氮元素密度(kg/m3);Tc为碳、氮元素储量(10 g/m2);Di为第i 层土壤容重(g/cm3);Mc为相应的土壤碳、氮元素含量(g/kg);di为第i层厚度(cm);n为土壤剖面分割的层数。
1.4 数据处理利用Origin 8.1软件对数据进行作图和描述性统计分析,运用SPSS 17.1软件对不同样地土壤的TC和TN含量垂直分布进行方差分析,并对pH、粒径、容重、电导以及TC、TN、NH+4-N、NO-3-N含量进行Pearson 相关分析。
2 结果与分析 2.1 入侵前后土壤理化性质的变化从图 2可知,在互花米草入侵短叶茳芏以后,在pH、电导率、容重和粒径分布上均有明显的变化规律,即入侵前后pH从表层到底层均表现为A>B>C,整体上3个采样点从表层到底层pH分别呈现减小趋势,说明互花米草入侵后提高了同层土壤的酸性;电导率在入侵前后表现为B>C>A,整体上3个采样点从表层到底层EC分别呈现增加趋势,说明互花米草入侵后提高了同层土壤的电导率;对于容重来说,入侵前后表现为A>B>C,3个采样点从表层到底层变化幅度较小,说明互花米草入侵后改变了土壤的物理性质,这在入侵前后土壤粒径方面表现的也较为明显,其A和C点土壤粘粒、粉粒和砂粒含量分别为(11.24±1.73)%、(62.00±1.66)%、(26.77±3.39)%和(7.79±0.41)%、(56.21±1.15)%、(35.82±1.57)%,与入侵前相比,入侵后湿地表层土壤的粘粒、粉粒含量显著降低了30.69%和9.34%,而砂粒含量显著增加了33.81%。

|
| 图 2 入侵前后土壤理化性质对比 Fig. 2 Comparison of physical and chemical properties of different soils in the process of Spartina alterniflora invasion |
尽管互花米草不同入侵序列土壤NO-3-N含量的垂直变化特征较为相似,但变化幅度差异较大,其值在A、B和C分别为(0.97±0.88)、(0.32±0.28)mg/kg和(0.61±0.52) mg/kg。整体而言,不同土层的NO-3-N含量整体均表现为A>C>B(P>0.05),说明互花米草入侵可显著降低土壤的NO-3-N含量(图 3)。不同入侵阶段土壤NH+4-N含量的垂直变化特征并不一致,且变化幅度差异不大,其值在入侵前、入侵边缘和入侵后分别为(15.75±3.90)、(19.80±4.32)mg/kg和(18.72±3.45) mg/kg。尽管不同土层的NH+4-N含量在入侵前后未达到显著水平(P>0.05),但互花米草入侵在一定程度上增加了土壤的NH+4-N含量(图 3)。不同入侵阶段土壤TC和TN的垂直变化特征均比较一致,其值在入侵前、入侵边缘和入侵后分别为(15.02±1.45)、(17.18±2.25)、(18.68±4.06) g/kg和(1.60±0.13)、(1.71±0.18)、(1.88±0.34) g/kg(图 4)。比较而言,0—6 cm和15—30 cm土层的TC和TN含量均表现为C>B>A(P<0.01),而在6—15 cm土层,三者均呈显著降低趋势,且碳氮含量较为接近(P>0.05),但整体上互花米草入侵还是增加了土壤的碳氮含量。此外,互花米草入侵前、入侵边缘以及入侵后的C/N比整体也表现为C>B>A(P<0.05),其均值分别为(9.50±0.08)、(9.94±0.11)和(10.24±0.21),说明互花米草入侵整体提高了入侵湿地土壤的C/N比。

|
| 图 3 互花米草入侵过程中土壤硝态氮和铵态氮含量变化 Fig. 3 Variations of nitrate nitrogen and ammonium nitrogen contents in different marsh soils in the process of Spartina alterniflora invasion |

|
| 图 4 互花米草入侵过程中土壤全碳和全氮含量变化 Fig. 4 Variations of total carbon and total nitrogen contents in different marsh soils in the process of Spartina alterniflora invasion |
研究发现,0—6 cm和15—30 cm土层的碳氮储量整体均表现为C>B>A(P<0.01),而在6—15 cm土层,三者的碳氮储量差异不大(P>0.05)(图 5),说明互花米草入侵显著改变了原生湿地环境,从而使得不同土层的碳氮储量整体呈极显著变化(P<0.01)。比较而言,互花米草不同入侵阶段土壤的碳储量呈明显增加趋势,其值在入侵前、入侵边缘和入侵后分别为(646.87±62.53)、(739.91±96.91)g/m2和(804.52±174.98) g/m2。与入侵边缘和入侵前相比,互花米草入侵后的碳储量分别增加了8.73%和24.37%。与之相比,不同入侵阶段土壤的氮储量呈缓慢增加趋势,其值在入侵前、入侵边缘和入侵后分别为(68.72±5.40)、(73.49±7.81) g/m2和(81.00±14.60) g/m2。与入侵边缘和入侵前相比,互花米草入侵后的氮储量分别增加了10.22%和17.87%(图 5)。

|
| 图 5 互花米草入侵过程中土壤碳氮储量变化 Fig. 5 Variations of carbon and nitrogen stocks in different marsh soils in the process of Spartina alterniflora invasion |
对于0—30 cm土层而言,互花米草入侵后的碳总储量为8486.12±21.24 g/m2,相比较入侵边缘的碳储量(7420.31±11.94) g/m2增加了14.36%,比入侵前的碳储量(6542.62±8.37) g/m2增加了29.71%;与之相比,互花米草入侵后的氮储量为(825.67±1.60) t/km2,比入侵边缘的碳储量(745.32±1.00) g/m2增加了10.78%,比入侵前的碳储量(687.95±0.78) g/m2增加了20.02%(图 6)。方差分析表明,互花米草入侵显著增加了短叶茳芏土壤碳氮储量,二者均表现为C>B>A(P <0.01)。

|
| 图 6 互花米草入侵过程中土壤(0—30cm)碳氮总储量变化 Fig. 6 Variations of carbon and nitrogen stocks in marsh soils (0—30 cm) in the process of Spartina alterniflora invasion |
与国内其他湿地的相关研究相似,闽江口互花米草入侵后湿地的碳氮含量均明显高于入侵湿地。不同的是,闽江口互花米草入侵后湿地以及入侵湿地的碳氮含量整体要高于江苏盐城、长江口以及杭州湾湿地的相关结果(表 1)。就闽江口而言,互花米草入侵后湿地的碳含量还与入侵湿地类型有关。本研究中,互花米草入侵短叶茳芏湿地的碳含量要接近于入侵芦苇湿地的碳含量,但明显高于入侵裸滩的相应数值(表 1)。上述对比结果表明,闽江口互花米草入侵对于原生芦苇和短叶茳芏湿地碳氮含量的影响可能更为显著。
| 研究区域Study areas | 碳含量/(g/kg)Carbon content | 氮含量/(g/kg)Total nitrogen content | 被入侵群落Invasion of community | 参考文献References | |||
| 碳形态Carbon forms | 互花米草S. alterniflora | 被入侵群落Invasion of community | 互花米草S. alterniflora | 被入侵群落Invasion of community | |||
| 江苏盐城 | TOC | 6.08 | 4.73 | 0.48 | 0.41 | 盐蒿 | [32] |
| SOC | 0.53—2.03(1.00)(1a) | [33] | |||||
| 0.36—8.93(1.70)(5a) | |||||||
| 0.36—8.93(1.70)(12a) | |||||||
| 0.82—11.33(2.49)(23a) | |||||||
| 长江口 | TOC | 5.57 | 2.40 | 碱蓬 | [34] | ||
| 杭州湾 | SOC | 6.46—6.78 | 4.33—4.48 | 0.69—0.77 | 0.39—0.5 | 海三棱藤草 | [35] |
| 闽江口 | SOC | 4.35 | 4.03 | 裸滩 | [31] | ||
| 17.16 | 15.54 | 芦苇 | [36] | ||||
| TC | 18.68±4.06 | 15.02±1.45 | 1.88±0.34 | 1.60±0.13 | 短叶茳芏 | 本研究 | |
研究表明,互花米草在入侵边缘以及入侵后均明显增加了短叶茳芏湿地土壤的碳氮含量和储量,而这可能与其在改变湿地植被分布格局的同时,亦提高了碳氮在植物-土壤系统中的累积能力有关。互花米草成功入侵后可迅速形成高密度与高生产力的单物种群落[37-38]。首先,与土著种短叶茳芏相比,互花米草入侵边缘和入侵后的地上生物量分别增加了237.09%和512.03%,生物量的大幅增加可明显增加碳的累积量。Liao等[3]的研究表明,与土著植物相比,互花米草生长期较长,且具有叶面积指数、净光合速率和根系生物量高等特征,由此使得其固碳能力很高。本研究亦得到类似结果,互花米草入侵后植被的固碳能力(0.85 kg C/m2)约为入侵边缘植被固碳能力(0.47 kg C/m2)的1.81倍。高建华等[32]的研究表明,互花米草作为研究区潮滩湿地的先锋植物,其生长和发育过程对潮滩淤积发挥着重要控制作用,滩面的沉积速率较高,有利于营养物质的埋藏和保存,增加了整个潮滩湿地每年TC、TN和TP的累积量[39];互花米草对整个潮滩湿地不同植被分布格局改变的同时也增加了TC和TN在整个生态系统中的累积量,而这种生物量、碳氮累积量的增加必然会增加入侵湿地土壤的碳氮含量和储量。其次,互花米草入侵边缘和入侵后凋落物数量、质量及分解速率亦可明显影响入侵湿地土壤的碳氮含量及储量[40]。由于互花米草生物量大且生长期较长,所以其凋落物产生量较短叶茳芏高很多[41]。另外,尽管互花米草入侵导致了植物根际微生物多样性和均匀度降低,使得其凋落物的分解速率略低于短叶茳芏[42],但其较高的凋落物累积量使得碳氮归还土壤的量依然较高。更为主要的是,互花米草根系更为发达,其根系周转速率更快[37],大量死根分解可将碳氮大量释放到不同土层中,进而显著增加入侵过程和入侵后湿地土壤的碳氮含量及储量[43-44]。本研究还表明,互花米草入侵后,6—15 cm土层的TC、TN含量呈现明显降低,原因可能主要与其入侵后中层土壤的砂粒含量明显增加有关。高建华等[32]关于互花米草入侵对苏北潮滩湿地碳、氮和磷分布影响的研究表明,互花米草入侵可导致沉积物中细颗粒比例的增加,进而可为营养物质的储存提供了有利条件。本研究中,互花米草入侵短叶茳芏湿地后,沉积物的容重明显降低而砂粒含量明显增加(图 2),进而不利于养分的存留,导致碳氮含量明显降低。
研究发现,互花米草入侵后湿地土壤的硝态氮整体随土壤深度的增加而增加,铵态氮含量则呈降低趋势(图 2),这主要与入侵后互花米草发达的植物根系促进了表层土壤中硝态氮的深层淋失有关,铵态氮则由于带正电荷,易于被土壤胶体吸附,由此使得其在表层土壤中含量较高,而深层土壤含量较低。另外,互花米草入侵后的硝态氮含量整体呈现降低趋势,而铵态氮呈一定增加趋势。这可能主要与入侵后互花米草根系可促进有机氮矿化及硝化作用有关。已有研究表明,湿地土壤氮的矿化作用与粘粒含量呈负相关,而各类型土壤氮矿化作用的大小一般表现为砂土>壤土>粘土[45]。本研究区的土壤类型为砂壤土,有利于矿化作用的进行。另有研究表明,植物可通过根系分泌的H+和某些具有解胶性的有机酸促进土壤有机氮的矿化,而根际某些生物活性较强微生物的富集也是促进氮矿化的一个重要原因。互花米草入侵后,根系周转能力增强,根际分泌物增多,加上发达根系通气组织可将氧气带至深层土壤,使得土壤矿化能力增强,铵态氮含量呈增加趋势。孙炳寅等[46]对互花米草入侵土壤微生物分布的研究还表明,互花米草的入侵可导致根际细菌、放线菌和真菌数量的增多,且在各生理类群中,氨化细菌和固氮细菌占优势,硝化细菌次之,说明互花米草根系的存在不但有助于土壤的矿化作用,而且硝化作用可能也随之增强。尽管硝化作用产生的硝态氮含量较高,但因硝态氮含量在水分条件较好的研究样地极易淋失,由此导致其含量整体呈降低趋势。另外,伴随着湿地土壤硝化能力的增强,硝化作用导致的氮气态损失可能也随之增加[47-49]。Groffman等[50]的研究还发现,具有较高硝化潜势的生态系统可能具有较高的氮损失量。可能正是由于这种硝化脱氮机制的存在,使得本研究中互花米草在入侵进程和入侵后湿地土壤的硝态氮含量呈降低趋势。
相关性分析表明,湿地土壤的TC与TN含量在互花米草入侵前后均呈极显著性正相关(P <0.01),说明二者在养分供给与循环方面的密切联系。另外,互花米草入侵前土壤的NH+4-N含量与pH呈显著正相关(P <0.05),但入侵后与土壤粘粒和粉粒含量呈显著负相关(P <0.05),与砂粒含量呈显著正相关(P <0.05),而与容重、电导无显著性相关关系(表 2)。已有研究表明,植物通过根系分泌的某些具有解胶性的有机酸和H+可促进土壤有机氮的矿化,同时根际某些生物活性较强微生物的富集亦可促进氮矿化[51-52]。但在本研究中,NH+4-N含量与pH在互花米草入侵后并未呈显著正相关关系(表 2),原因可能主要与其入侵后土壤颗粒组成发生显著变化,进而促进土壤的矿化作用有关。与入侵前相比,入侵后湿地表层土壤的粘粒、粉粒含量显著降低(分别降低了30.69%和9.34%),而砂粒含量显著增加(增加了33.81%),而这在一定程度上增加了土壤的孔隙度,改善了土壤的通气状况,进而使得铵态氮含量有所增加。这一结果正好与前述得出的互花米草入侵后尽管土壤pH明显降低但砂粒含量和NH+4-N含量整体增加的研究结果正好吻合(图 2,图 3)。对于NO-3-N而言,其含量在入侵后与粘粒含量呈显著正相关(P <0.05),而与砂粒含量呈显著负相关(P <0.05)(表 2),而这可能亦主要与互花米草自身的强促淤作用导致土壤颗粒组成发生一定改变有关。入侵后,湿地表层土壤的砂粒含量显著增加,这在一定程度上增加了土壤的孔隙度,改善了土壤的通气状况,而土壤孔隙度的增加尽管有助于土壤硝化作用的进行,但硝态氮的垂直淋失亦同时增加,这一结果也与前述得出的互花米草入侵可导致硝态氮含量降低的结论相吻合。
| 指标Index | pH | 粘粒Clay | 粉粒Silt | 砂粒Sand | 容重 Bulk density | 电导Electrical conductivity | 氨态氮NH+4-N | 硝态氮NO-3-N | 总氮TN | 总碳TC | 碳氮比C/N ratio |
| pH | 1 | B-0.149 | B-0.186 | B0.176 | B0.132 | B-0.177 | B-0.131 | B-0.175 | B-0.412* | B-0.347 | B-0.164 |
| 粘粒Clay | A 0.084 | 1 | B0.885** | B-0.963** | B0.158 | B0.053 | B-0.512* | B0.453* | B-0.195 | B-0.151 | B-0.071 |
| 粉粒Silt | A -0.170 | A0.814** | 1 | B-0.978** | B0.031 | B0.113 | B-0.440* | B0.396 | B0.003 | B0.025 | B0.036 |
| 砂粒Sand | A0.663** | A0.379 | A0.358 | 1 | B-0.086 | B-0.094 | B0.485* | B-0.435* | B0.084 | B0.05 | B0.008 |
| 容重Bulk density | A0.013 | A-0.176 | A-0.107 | A0.101 | 1 | B-0.608** | B-0.094 | B-0.38 | B-0.199 | B-0.217 | B-0.21 |
| 电导Electrical conductivity | A-0.591** | A0.012 | A0.263 | A-0.347 | A-0.314 | 1 | B-0.055 | B0.34 | B0.19 | B0.222 | B0.208 |
| 氨态氮NH+4-N | A0.460* | A0.130 | A0.000 | A0.382 | A-0.068 | A-0.320 | 1 | B-0.137 | B0.365 | B0.166 | B-0.144 |
| 硝态氮NO-3-N | A0.064 | A0.107 | A0.116 | A0.367 | A-0.080 | A-0.348 | A0.147 | 1 | B0.072 | B0.114 | B0.14 |
| 总氮TN | A0.000 | A0.147 | A-0.014 | A-0.023 | A-0.227 | A0.173 | A-0.140 | A0.079 | 1 | B0.921** | B0.555** |
| 总碳TC | A0.173 | A0.263 | A-0.109 | A0.050 | A-0.101 | A-0.028 | A-0.111 | A0.041 | A0.862** | 1 | B0.833** |
| 碳氮比C/N ratio | A0.376 | A0.261 | A-0.197 | A0.167 | A0.206 | A-0.385 | A0.015 | A-0.061 | A-0.033 | A0.477* | 1 |
| *P <0.05;**P <0.01;左上角A、B分别表示入侵前和入侵后 | |||||||||||
互花米草具有高基因渗入能力、高遗传分化能力、高适应性及高抗性,其蔓延的速度相当惊人。互花米草植株高大密集,株高一般为1—3 m,直径在1 cm以上。根系发达,常密布于地下30 cm深的土层内,有时可深达50—100 cm[53]。一方面,互花米草生长年份越久,其向土壤输入的有机质含量就越高;另一方面,互花米草入侵在改变研究区潮滩淤积状况的同时,亦改变了土壤有机质的来源。杨桂山等[54]对苏北潮滩湿地的研究表明,互花米草的多年平均沉积速率达到3 cm/a,且这种淤积改变了土壤有机质的来源。土壤C/N比经常被用于辨别有机质的陆源性和海源性,陆源植物由于富含纤维素,其C/N比往往高于12,而海水中水草富含蛋白质,其C/N比通常介于6—9[55]。本研究中,互花米草入侵前、入侵边缘和入侵后湿地土壤的C/N比分别为(10.24±0.62)、(9.94±0.30)和(9.51±0.29),均小于12,说明研究样地的有机质主要来自海源输人。但从互花米草入侵过程来看,C/N沿入侵方向呈降低趋势,这一递减趋势在一定程度上反映出互花米草入侵正在改变潮滩沉积物中陆源和海源有机质的比例。综上可知,随着互花米草入侵时间的延长,互花米草来源有机质(自源有机质)的比例将呈增加趋势,而海源有机质的比例则呈降低趋势。这一研究结果正好与杭子清[33]对江苏盐城互花米草入侵湿地的研究结果相吻合,即随着入侵年限的延长,来源于互花米草的湿地土壤有机质含量呈增加趋势(表 1)。可见,互花米草在增强湿地淤积能力的同时亦改变了沉积物中有机质的来源比例,使得入侵后湿地土壤养分的自源性增强(提供有机质和养分的能力增强)。本研究从互花米草入侵导致的养分自源性加强以及促淤导致的沉积物养分输入增加角度揭示了其入侵不断增强的关键机制。
互花米草在其不同生长阶段对土壤碳氮含量的确存在一定影响,这就使得开展不同季节互花米草入侵序列对土壤碳氮含量动态变化的影响研究显得尤为必要。但由于闽江口国家级自然保护区的互花米草在春季后常常进行物理割除治理,由此使得开展互花米草不同生长阶段对土壤碳氮含量的影响研究存在一定难度。今后应选择更具典型的替代样地进行分季节采样,并将研究结果与当前数据进行对比分析,以更好的揭示互花米草入侵时间序列对湿地土壤碳氮含量的影响。
4 结论(1) 互花米草入侵显著降低土壤的NO-3-N含量,但整体增加了土壤的NH+4-N含量,这与其入侵后导致湿地土壤颗粒组成发生显著变化,进而促进了土壤的矿化作用和硝化作用,并有助于硝态氮的垂直淋失有关。
(2) 互花米草不同入侵过程土壤TC、TN含量以及C/N比的垂直变化特征均比较一致,入侵整体增加了土壤的碳氮含量和C/N比和土壤的碳氮储量,这主要与其对闽江口湿地植物群落格局、养分生物循环以及强促淤作用引起的土壤颗粒组成等显著改变有关。
(3) 闽江口互花米草入侵对短叶茳芏湿地土壤碳氮含量的影响相对于江苏盐城、长江口以及杭州湾湿地的影响可能更为显著,其互花米草入侵较大改变了土壤中陆源和海源有机质的来源比例,使得入侵后湿地土壤养分的自源性增强。
致谢: 野外采样得到福建师范大学地理科学学院汪旭明、杨平、章文龙等同学的帮助,特此致谢。| [1] | Simas T, Nunes J P, Ferreira J G.Effects of global climate change on coastal salt marshes.Ecological Modelling,2001,139(1): 1–15. |
| [2] | 辛沛, 李凌滨.滨海盐沼孔隙水动力过程研究综述.湿地科学,2007,5(4): 376–384. |
| [3] | Liao C Z, Luo Y Q, Jiang L F, Zhou X H, Wu X W, Fang C M, Chen J K, Li B.Invasion of Spartina alterniflora enhanced ecosystem carbon and nitrogen stocks in the Yangtze Estuary, China.Ecosystems,2007,10(8): 1351–1361. |
| [4] | Mozdzer T J, Zieman J C, McGlathery K J.Nitrogen uptake by native and invasive temperate coastal macrophytes:importance of dissolved organic nitrogen.Estuaries and Coasts,2010,33(3): 784–797. |
| [5] | 张耀鸿, 王艳玲, 李仁英, 朱红霞, 张富存, 黄宾宾, 彭晓丹.互花米草入侵对潮滩土壤活性有机碳组分的影响.土壤通报,2012,43(1): 102–105. |
| [6] | 王刚, 杨文斌, 王国祥, 刘金娥, 杭子清.互花米草海向入侵对土壤有机碳组分、来源和分布的影响.生态学报,2013,33(8): 2474–2483. |
| [7] | 冯振兴, 高建华, 陈莲, 汪亚平, 高建慧, 白凤龙.互花米草生物量变化对盐沼沉积物有机碳的影响.生态学报,2015,35(7): 2038–2047. |
| [8] | Peng R H, Fang C M, Li B, Chen J K.Spartina alterniflora invasion increases soil inorganic nitrogen pools through interactions with tidal subsidies in the Yangtze Estuary, China.Oecologia,2011,165(3): 797–807. |
| [9] | 廖成章. 外来植物入侵对生态系统碳、氮循环的影响:案例研究与整合分析[D]. 上海:复旦大学, 2007 |
| [10] | 彭容豪. 互花米草对河口盐沼生态系统氮循环的影响——上海崇明东滩实例研究[D]. 上海:复旦大学, 2009 |
| [11] | 袁月. 崇明东滩湿地芦苇与互花米草种群间关系格局与影响因素研究[D]. 上海:华东师范大学, 2014. |
| [12] | 曾艳, 田广红, 陈蕾伊, 李静, 安东, 雷振胜, 唐虹, 彭少麟.互花米草入侵对土壤生态系统的影响.生态学杂志,2011,30(9): 2080–2087. |
| [13] | 张骁栋. 互花米草与蟹类扰动对崇明东滩植物种间关系及生地化循环的影响[D]. 上海:复旦大学, 2012 |
| [14] | 沈永明, 杨劲松, 曾华, 周勤.我国对外来物种互花米草的研究进展与展望.海洋环境科学,2008,27(4): 391–396. |
| [15] | 项剑, 刘德燕, 袁俊吉, 丁维新.互花米草入侵对沿海湿地甲烷排放的影响.生态学杂志,2012,31(6): 1361–1366. |
| [16] | 何清华, 黄佳芳, 王维奇, 仝川.刈割治理互花米草对河口沼泽CO2和CH4排放通量的短期效应.湿地科学,2013,11(2): 204–211. |
| [17] | 李杨杰, 陈振楼, 王东启, 胡泓, 王初.长江口盐沼带湿地生态演替过程中甲烷排放研究.环境科学学报,2014,34(8): 2035–2042. |
| [18] | Sun Z G, Sun W G, Tong C, Zeng C S, Yu X, Mou X J.China's coastal wetlands:conservation history, implementation efforts, existing issues and strategies for future improvement.Environment International,2015,79 25–41. |
| [19] | Zhang W L, Zeng C S, Tong C, Zhang Z C, Huang J F.Analysis of the expanding process of the spartina alterniflora salt marsh in shanyutan wetland, minjiang river estuary by remote sensing.Procedia Environmental Sciences,2011,10((Part C)): 2472–2477. |
| [20] | 李丽纯, 林瑞坤, 吴振海, 陈家金, 杨凯, 徐宗焕.闽江口湿地互花米草地上生物量的月动态特征.资源科学,2009,31(11): 1967–1972. |
| [21] | 仝川, 李旭伟, 雍石泉, 杨平, 黄佳芳.闽江口互花米草不同治理方式对土壤有机碳及养分含量的影响.水土保持学报,2012,26(4): 138–142. |
| [22] | 翟水晶, 田琳琳, 仝川.闽江河口短叶茳芏和互花米草沼泽土壤剖面间隙水营养盐含量比较.湿地科学,2013,11(1): 68–74. |
| [23] | 曾从盛, 张林海, 仝川.闽江河口湿地短叶茳芏氮、磷含量与积累量季节变化.生态学杂志,2009,28(5): 788–794. |
| [24] | 曾从盛, 张林海, 王天鹅, 张文娟, 仝川.闽江河口湿地植物枯落物立枯和倒伏分解主要元素动态.生态学报,2012,32(20): 6289–6299. |
| [25] | Zhang L H, Tong C, Marrs R, Wang T E, Zhang W L, Zeng C S.Comparing litter dynamics of Phragmites australis and Spartina alterniflora in a sub-tropical Chinese estuary:contrasts in early and late decomposition.Aquatic Botany,2014,117 1–11. |
| [26] | 仝川, 曾从盛, 王维奇, 闫宗平, 杨红玉.闽江河口芦苇潮汐湿地甲烷通量及主要影响因子.环境科学学报,2009,29(1): 207–216. |
| [27] | 仝川, 鄂焱, 廖稷, 姚顺, 王维奇, 黄佳芳, 张林海, 杨红玉, 曾从盛.闽江河口潮汐沼泽湿地CO2排放通量特征.环境科学学报,2011,31(12): 2830–2840. |
| [28] | 仝川, 姚顺, 王维奇, 黄佳芳, 张林海, 章文龙, 曾从盛.中国东南沿海短叶茳芏潮汐沼泽湿地甲烷动态.中国科学:地球科学,2012,42(5): 723–735. |
| [29] | 张永勋, 曾从盛, 黄佳芳, 王维奇, 仝川.人为干扰对闽江河口短叶茳芏湿地N2O排放的影响.中国环境科学,2013,33(1): 138–146. |
| [30] | 王宝霞, 曾从盛, 陈丹, 王维奇, 张林海.互花米草入侵对闽江河口芦苇湿地土壤有机碳的影响.中国水土保持科学,2010,8(5): 114–118. |
| [31] | 潘婷, 曾六福, 曾从盛, 王维奇.互花米草入侵对闽江河口裸滩湿地土壤有机碳的影响.中国水土保持科学,2015,13(1): 84–89. |
| [32] | 高建华, 杨桂山, 欧维新.互花米草引种对苏北潮滩湿地TOC、TN和TP分布的影响.地理研究,2007,26(4): 799–808. |
| [33] | 杭子清, 王国祥, 刘金娥, 王刚, 王会.互花米草盐沼土壤有机碳库组分及结构特征.生态学报,2014,34(15): 4175–4182. |
| [34] | 张天雨, 葛振鸣, 张利权, 陈怀璞, 严格.崇明东滩湿地植被类型和沉积特征对土壤碳、氮分布的影响.环境科学学报,2015,35(3): 836–843. |
| [35] | 张文敏, 吴明, 王蒙, 邵学新, 姜小三, 周斌.杭州湾湿地不同植被类型下土壤有机碳及其组分分布特征.土壤学报,2014,51(6): 1351–1360. |
| [36] | 王宝霞. 闽江口鳝鱼滩入侵种互花米草与土著种芦苇湿地土壤碳库特征比较[D]. 福州:福建师范大学, 2011. |
| [37] | 邓自发, 安树青, 智颖飙, 周长芳, 陈琳, 赵聪蛟, 方淑波, 李红丽.外来种互花米草入侵模式与爆发机制.生态学报,2006,26(8): 2678–2686. |
| [38] | 沈永明, 王艳芳, 陈寿军, 吴德力, 虞慰岩, 李朝晖.互花米草盐沼湿地大型底栖动物时空分布特征.地理研究,2013,32(4): 638–644. |
| [39] | Mayer L M, Macko S A, Cammen L.Provenance, concentrations and nature of sedimentary organic nitrogen in the gulf of Maine.Marine Chemistry,1988,25(3): 291–304. |
| [40] | Liu J E, Zhou H X, Qin P, Zhou J.Effects of Spartina alterniflora salt marshes on organic carbon acquisition in intertidal zones of Jiangsu Province, China.Ecological Engineering,2007,30(3): 240–249. |
| [41] | 欧阳林梅, 王纯, 王维奇, 仝川.互花米草与短叶茳芏枯落物分解过程中碳氮磷化学计量学特征.生态学报,2013,33(2): 389–394. |
| [42] | 刘白贵, 仝川, 罗榕婷.闽江河口湿地3种主要植物冬春季枯落物分解特征.福建师范大学学报:自然科学版,2008,24(2): 80–85. |
| [43] | France R, Culbert H, Freeborough C, Peters.Leaching and early mass loss of boreal leaves and wood in oligotrophic water.Hydrobiologia,1997,345(2/3): 209–214. |
| [44] | Wang X C, Chen R F, Berry A.Sources and preservation of organic matter in Plum Island salt marsh sediments (MA, USA):long-chain n-alkanes and stable carbon isotope compositions.Estuarine, Coastal and Shelf Science,2003,58(4): 917–928. |
| [45] | 孙志高, 刘景双, 王金达, 李新华, 杨继松.湿地生态系统土壤氮素矿化过程研究动态.土壤通报,2007,38(1): 155–161. |
| [46] | 孙炳寅, 朱长生.互花米草(Spartina alterniflora)草场土壤微生物生态分布及某些酶活性的研究.生态学报,1989,9(3): 240–244. |
| [47] | 孙志高, 刘景双, 杨继松, 李新华, 周旺明.三江平原典型小叶章湿地土壤硝化-反硝化作用与氧化亚氮排放.应用生态学报,2007,18(1): 185–192. |
| [48] | Wrage N, Velthof G L, van Beusichem M L, Oenema O.Van M L.Soil Biology and Biochemistry,2001,33(12-13): 1723–1732. |
| [49] | 孙文广, 孙志高, 甘卓亭, 孙万龙, 王伟.黄河口不同恢复阶段湿地土壤N2O产生的不同过程及贡献.环境科学,2014,35(8): 3110–3119. |
| [50] | Groffman P M, Hanson G C, Erick K, Stevens G.Variation in microbial biomass and activity in four different wetland types.Soil Science Society of America Journal,1996,60(2): 622–629. |
| [51] | ]NartehL T, SahrawatK L.Potentially mineralizable nitrogen in West African lowland rice soils.Geoderma,1997,76(1/2): 145–154. |
| [52] | Fioretto A.Decomposition dynamics of litters of various pine species in Corsican pine forest.Soil Biology and Biochemistry,1998,30(6): 721–727. |
| [53] | 王卿, 安树青, 马志军, 赵斌, 陈家宽, 李博.入侵植物互花米草-生物学、生态学及管理.植物分类学报,2006,44(5): 559–588. |
| [54] | 杨桂山, 施雅风, 秀子修.江苏淤泥质潮滩对海平面变化的形态响应.地理学报,2002,57(1): 76–84. |
| [55] | Cifuentes L A, Coffin R B, Solorzano L, Cardenas W, Espinoza J, Twilley R R.Isotopic and elemental variations of carbon and nitrogen in a mangrove estuary.Estuarine, Coastal and Shelf Science,1996,43(6): 781–800. |