 2016, Vol. 36
2016, Vol. 36文章信息
- 颉洪涛, 何兴东, 尤万学, 余殿, 刘惠芬, 王金龙, 古松, 聂庆华, 梁玉婷, 张京磊
- XIE Hongtao, HE Xingdong, YOU Wanxue, YU Dian, LIU Huifen, WANG Jinlong, GU Song, NIE Qinghua, LIANG Yuting, ZHANG Jinglei.
- 哈巴湖国家级自然保护区油蒿群落生态化学计量特征对群落生物量和物种多样性的影响
- Effects of ecological stoichiometry on biomass and species diversity of the Artemisia ordosica community in Habahu National Nature Reserve
- 生态学报[J]. 2016, 36(12): 3621-3627
- Acta Ecologica Sinica[J]. 2016, 36(12): 3621-3627
- http://dx.doi.org/10.5846/stxb201506171229
-
文章历史
- 收稿日期: 2015-06-17
- 修订日期: 2016-01-18





2. 宁夏哈巴湖国家级自然保护区管理局, 宁夏 751501;
3. 天津农学院, 天津 300384;
4. 南开大学环境科学与工程学院, 天津 300071
2. Ningxia Habahu National Nature Reserve Administration, Ningxia 751501, China;
3. Tianjin Agricultural University, Tianjin 300384, China;
4. College of Environmental Science and Engineering, Nankai University, Tianjin 300071, China
植物营养元素及元素化学计量特征是现代生态学的重要研究内容[1]。生态化学计量学(ecological stoichiometry)则是研究生态过程中多种化学元素(主要是C、N、P)平衡关系的学科,并试图通过元素比率把这些小到分子、细胞,大到群落、生态系统的研究尺度联系起来,在一定程度上为生态学的发展构建了一个统一的理论框架[2-3]。生态化学计量特征不仅影响生物的生长率和体内营养物质的合成,而且影响群落结构和动态变化、营养级动态,还影响共生物种的选择性、养分限制、生态系统养分循环和供求平衡等[3-4]。生态化学计量特征可用来判断植物养分限制状况、揭示植物对营养元素的利用策略等[1, 5]。
随着全球环境的变化,陆地生态系统的结构、过程、功能、植被分布格局和演替过程也在发生显著的变化[6],其中,植物群落生态化学计量特征变化的作用越来越受到国内外学者的关注。研究发现,不同地带植物叶N∶P不同[7-8],生长季植物叶C∶N比、C∶P比和N∶P比变化趋势不同[9],植物叶C∶N比和N∶P比与草原退化程度相关[10],这说明不同植物群落的植物生长与生态化学计量特征变化有关,事实上,生态化学计量特征影响着植物生长率[3, 11]。研究表明,N∶P比生态化学计量特征的变化会对植物生长构成限制[12-13],分为氮限制、磷限制和氮磷共同限制,进而影响群落生产力和物种多样性[14-15]。资源平衡假说[16]表明,物种丰富度是由潜在限制养分的平衡供给所孕育,即在氮磷供给平衡(N∶P比为10—15,氮磷共同限制)时,植被物种丰富度最大。对于沙地生态系统而言,由于沙土中氮磷含量较低,植物N∶P比生态化学计量特征呈现什么样的状态以及它对植物群落生产力和物种多样性会产生怎样的影响是一个让人感兴趣的问题。为此,本研究选择哈巴湖国家级自然保护区内沙地生态系统中的油蒿群落为研究对象,测定分析了不同发育阶段油蒿群落植物C、N、P生态化学计量特征,探讨了油蒿群落植物生态化学计量特征对群落生物量、物种多样性的影响。
1 研究区概况宁夏哈巴湖国家级自然保护区处于黄土高原向鄂尔多斯台地过渡、半干旱区向干旱区过渡、干草原向荒漠草原过渡、灰钙土向棕钙土过渡、农区向牧区过渡的交错地带,境内生态系统相互交错、类型多样,有灌丛、草原、草甸、荒漠、湿地和沙地等生态系统。
研究区位于宁夏回族自治区盐池县中北部的哈巴湖国家级自然保护区内(37°38′36″—38°02′00″N,106°53′10″—107°38′10″E之间),海拔1300—1622 m,属于干旱、半干旱区及草原、荒漠草原区过渡带。气候干燥,雨量少而集中,蒸发强烈,温差大,日照长。年均气温7.7℃,年均日照2867.9 h,平均无霜期128 d,年均降雨量270 mm,80%多集中在7—9月份。年均蒸发量2727.24 mm,是全年降水量的9.6倍。土壤主要有灰钙土、风沙土、潮土、盐土、新积土、棕钙土和黄绵土等[17]。植被以沙生植被和盐碱地植被为主,主要植物有油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)、长芒草(Stipa bungeana)、猫头刺(Oxytropis aciphylla)、草木犀状黄芪(Astragalus melilotoides)、达乌里胡枝子(Lespedeza davurica)、白刺(Nitraria tangutorum)、红砂(Reaumuria soongorica)、细枝盐爪爪(Kalidium gracile)、甘草(Glycyrrhiza uralensis)、苦豆子(Sophora alopecuroides)和芨芨草(Achnatherum splendens)、白草(Pennisetum centrasiaticum)、黄蒿(A. scoparia)、赖草(Aneurolepidium dasystachys)、假苇拂子茅(Calamagrostis pseudophragmites)、绳虫实(Corispermum deslinatum)、猪毛菜(Salsola Pall.)、二裂委陵菜(Potentilla bifurca)、鹅绒藤(Cynanchum chinensis)、菟丝子(Cuscuta chinensis)等。
2 研究方法 2.1 野外调查和取样2014年7月中下旬,在油蒿群落中,选取油蒿不同发育时期的6个典型群落,即①油蒿-虫实+猪毛菜群落、②油蒿-假苇拂子茅群落、③油蒿-白草群落、④油蒿-白草+赖草群落、⑤油蒿-苦豆子群落、⑥油蒿-黄蒿-苦豆子群落,其中,①和②为油蒿群落发育的早期阶段,③和④为中期阶段,⑤和⑥为后期阶段。在每个群落中,分别随机设置一条50 m样线,每隔10 m设置一个4 m × 4 m样方,调查样方内的油蒿密度和盖度;然后,在4 m × 4 m样方中,再随机设置一个1 m × 1 m小样方,调查草本植物的种类、密度、盖度,然后切割地上部,称量鲜重,带回实验室后,用烘箱烘干(80℃烘24 h),称量干重。在群落调查的同时,足量采集样方中植物新鲜叶片封存于信封袋中,贴签带回实验室用于C、N、P含量测定。
2.2 样品处理及测定将植物样品在信封内自然风干后打磨成粉末状,存于信封中用于实验分析。植物叶碳含量用重铬酸钾容量法-外加热法[18]测定,氮含量用浓H2SO4-H2O2消煮-半微量凯氏定氮法[18]测定,磷含量用浓H2SO4-H2O2消煮-钼锑抗比色法[18]测定。
2.3 数据分析油蒿群落植物叶C∶N∶P比为样方内所有植物叶C∶N∶P比的加权平均值[12, 19],每个物种的生物量为加权因子,计算时,先计算加权的C、N、P含量,再计算C∶N∶P比。物种多样性为Shannon-Wiener指数, H′=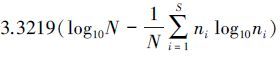
用Excel 2010进行数据整理,并计算植物叶C∶N∶P比以及群落生物量和物种多样性的平均值和标准差。然后,用SPSS 20.0进行单因素方差分析(One-way ANOVA),首先对数据进行方差齐性检验,方差齐性时,用Duncan进行组间差异显著性检验分析;当方差非齐性时,用Tamhane进行组间差异显著性检验分析。在Excel 2010中进行油蒿群落植物叶C∶N∶P比群落生物量和物种多样性的回归与作图。
3 结果与分析 3.1 油蒿群落植物叶C、N、P含量及其化学计量特征结果表明,从油蒿群落早期、中期到后期,植物叶C、N、P营养状况差异明显(表 1),限制元素的作用程度也在逐渐发生变化。分析表明,6个油蒿群落平均叶C∶N比为20.92 ± 8.14,N∶P比为22.18 ± 13.99,C∶P比为410.80 ± 217.36。从C∶N∶P比化学计量特征来看,植物叶C∶N比的变化范围为15.86—25.42,N∶P比的变化范围为10.65—33.70,C∶P比的变化范围为261.51—539.74(表 2)。C∶N比从油蒿早期到中期再到后期呈现逐渐变小的趋势,其中从油蒿早期到油蒿中期以及从油蒿中期到油蒿后期,植物叶C∶N比均显著减小(P <0.05)。植物叶N∶P比从油蒿早期到中期再到后期呈现逐渐变大的趋势,且各个时期差异显著(P <0.05),从早期的10.65 ± 1.00增加到后期的33.70 ± 4.83,增长幅度相对较大。植物叶C∶P比从油蒿早期到油蒿中期以及从中期到后期均有增大的趋势,但差异均不显著。
| 发展阶段Stage | 碳含量/(g/kg) C content | 全氮含量/(g/kg) Total N concentration | 全磷含量/(g/kg) Total P concentration |
| 油蒿早期Early | 496.44 ± 9.79 a | 22.27 ± 2.27 | 2.339 ± 0.319 a |
| 油蒿中期Middle | 443.59 ± 7.62 b | 24.14 ± 2.84 | 1.441 ± 0.175 b |
| 油蒿后期Final | 442.75 ± 6.56 b | 28.85 ± 1.27 | 1.266 ± 0.175 b |
| 数据为平均值士标准误(Data are mean±SE),同列不同小写字母表示在0. 05水平差异显著 | |||
| 发展阶段Stage | C∶N比 C∶N ratio | N∶P比 N∶P ratio | C∶P比 C∶P ratio |
| 油蒿早期Early | 25.42 ± 2.96 a | 10.65 ± 1.00 c | 261.51 ± 29.88 b |
| 油蒿中期Middle | 21.45 ± 2.58 ab | 22.16 ± 3.04 b | 431.13 ± 37.33 ab |
| 油蒿后期Final | 15.86 ± 0.86 b | 33.70 ± 4.83 a | 539.74 ± 92.55 a |
| 数据为平均值士标准误(Data are mean土SE),同列不同小写字母表示在0. 05水平差异显著 | |||
油蒿-虫实群落、油蒿-假苇拂子茅群落、油蒿-白草群落、油蒿-白草-赖草群落、油蒿-苦豆子群落和油蒿-黄蒿-苦豆子群落,这6个群落中植物叶的C∶N比、N∶P比、物种多样性以及群落生物量具有显著的差异(图 1)。分析表明,油蒿群落发育后期阶段的植物(⑤和⑥)具有较低的C∶N比和较高的N∶P比,而油蒿群落发育早期阶段的植物(①和②)具有较高的C∶N比和较低的N∶P比。

|
| 图 1 油蒿群落不同发育阶段植物叶碳氮比、氮磷比、物种多样性和生物量变化 Fig. 1 Plant foliar C∶N ratio,N∶P ratio,species diversity and biomass in different stages of A. ordosica community |
进一步回归结果表明,油蒿群落植物物种多样性和生物量与叶C∶N比和N∶P比之间具有显著的回归关系(图 2),随着叶C∶N比的增大,油蒿群落生物量呈指数函数显著降低(P <0.05),物种多样性呈对数函数降低的趋势,但不显著;而随着叶N∶P比的增大,油蒿群落生物量和物种多样性均呈幂函数显著增加(P <0.05)。

|
| 图 2 油蒿群落植物叶碳氮比、氮磷比、物种多样性和生产力之间回归关系 Fig. 2 Regression relationships among plant foliar C∶N ratio,N∶P ratio,species diversity and biomass in A. ordosica community |
本研究结果表明,6个油蒿群落植物叶C含量低于全球 492 种陆生植物叶C平均含量[20],但叶N含量高于中国陆地植物平均水平,叶P含量也高于中国陆地植物平均水平[21]。植物叶C、N和P含量变化说明了植物在不同生存环境中的元素利用策略[22]。本研究中,油蒿群落生长在相对干旱的环境中,植物体内保持较高的叶N、P含量可能有利于适应高温干旱环境生存,这一结果证实了干旱环境植物平均叶N含量相对较高的结论[23-25]。
在研究区,油蒿作为沙地中最重要的固沙植物,具有广泛的分布范围和绝对的主导地位,随着伴生种的变化而形成一个连续的油蒿群落发展过程。在这个发展过程中,油蒿可存活30 a左右,可以将油蒿的发展过程明显地分为油蒿早期、中期和后期3个发育时期。用空间代替时间的方法,分析了油蒿发展过程中各个时期的植物叶C∶N∶P比化学计量特征。根据Koerselam和Meuleman的研究结果,当N∶P<14时,群落水平上的植物生长主要受N限制;而N∶P>16时,植物生长主要受P限制[12],据此分析,油蒿群落发育前期,植物生长主要是受N元素限制,发育中后期,植物生长主要是受P元素限制。
具体来说,从油蒿群落早期到后期的发展过程中,植物叶C∶N比总体呈现逐步变小的趋势,且油蒿早期和中期的变化差异显著,中期到后期的变化差异不显著。在这一过程中,植物叶的C含量是总体呈变小趋势的,而氮含量总体上是呈现增加趋势的,因此C∶N比呈现出变小的趋势。植物叶N∶P从油蒿早期到中期再到后期呈增加趋势,且各个时期的差异显著。在油蒿早期,植物叶N∶P比为11,说明在这一时期,植物生长主要受N元素的限制;到了油蒿中期和后期,N∶P比分别变为18和33,说明在油蒿中期和后期,植物的营养状况发生了明显的变化,由早期的受N元素限制转变成为受P元素限制,这与单独测定的植物体内磷含量的变化情况是完全符合的。植物叶C∶P比从油蒿早期到中期,有减小趋势但不显著,从油蒿中期到后期,植物叶C∶P比显著增大,在这一时期,虽然植物叶C和P含量都是减小的,但二者的变化速度不同,植物叶的碳含量减小速度明显低于同时期内磷含量的减小速度。从植物叶C∶N∶P比的总体变化情况来看,在油蒿群落从前期到后期的发展过程中,植物的生长逐渐由受N元素限制转变成为受P元素限制,这可能与竞争及生物因子综合作用有关[12]。进一步分析表明,油蒿群落发育早期阶段的植物具有较高的叶C∶N比和较低的叶N∶P比,而油蒿群落发育后期阶段的植物具有较低的叶C∶N比和较高的叶N∶P比,这符合生长率假说[3-4]。
4.2 油蒿群落植物叶C∶N∶P化学计量特征与物种多样性和群落生物量的关系植物物种多样性和群落生产力的变化是植物与植物之间以及植物与环境之间长期作用的结果[26]。植物与植物之间的竞争、光照、水分和养分状况是最重要的影响因素[27]。就植物养分而言,氮和磷的不足或者相对比率变化范围超过一定限度会限制植物的生长,植物的生长速率会发生变化,进而影响植物密度[28],进一步可能会对植物物种多样性和群落生产力产生相应的影响。本文以研究区油蒿不同发育时期6个群落为研究对象,来探讨植物叶C∶N∶P比特征与植物群落生物量和物种多样性之间的关系。
本研究结果表明,随着油蒿群落植物叶C∶N比的增大,群落生物量和物种多样性逐渐降低,而随着叶N∶P比的增大,群落生产力和物种多样性均显著提高,说明植物叶C∶N∶P比特征对植物群落生物量和物种多样性有显著影响。由于C在植物体内的含量很高且较为稳定[29-30],因此N含量变化是影响C∶N比的主要因素,C∶N比的增大主要是由于植物叶中N含量的减少造成的,而N含量的不足会影响植物的生长率,从而降低物种多样性和生产力,因此叶C∶N比的增大降低了群落生物量和物种多样性。而就随着油蒿群落植物叶N∶P比的增大,群落生物量增大,这一现象看起来与生长率假说不相吻合,分析原因可能是由于在沙地上生长的油蒿群落处于群落演替的早期阶段,就整个油蒿群落而言,在演替系列中,油蒿群落处于植物密度和生物量上升阶段。由本文前面的分析可知,油蒿群落植物叶P含量高于中国陆地植物平均水平,植物的生长主要受N限制,受P限制作用小于受N限制作用,因此N含量增加对研究区油蒿群落生产力和物种多样性的贡献更大,本研究还发现,植物叶P含量的稳定性强于N元素,变化范围相对较小,这也从侧面突出显示了植物叶N元素含量变化的重要影响。也可能基于同样的原因,随着油蒿群落植物叶N∶P比的增大,物种多样性增大。因此,N含量的变化是造成油蒿群落植物叶C∶N比和N∶P比变化的主要因素,在油蒿群落水平上表现为物种多样性和群落生物量产生相应变化。
陈军强等研究了祁连山东部亚高山草甸高产草地和低产草地植物群落物种多样性与植物群落C、N、P生态化学计量特征的关系[31],结果表明,高产样地上植物群落的物种数与N∶P呈显著正相关关系,与C∶N呈显著负相关关系;低产样地上植物群落的物种数与C∶N呈显著正相关关系,但和N∶P无明显相关关系。本研究结果与他们的结果有一定的共同性。生态化学计量特征之所以能够影响群落生物量和物种多样性是由于群落存在N限制或者P限制[13-14],有关这一问题的发生机制还有待深入研究。
| [1] | 贺金生, 韩兴国.生态化学计量学:探索从个体到生态系统的统一化理论.植物生态学报,2010,34(1): 2–6. |
| [2] | 王绍强, 于贵瑞.生态系统碳氮磷元素的生态化学计量学特征.生态学报,2008,28(8): 3937–3947. |
| [3] | Elser J J, Sterner R W, Gorokhova E, Fagan W F, Markow T A, Cotner J B, Harrison J F, Hobbie S E, Odell G M, Weider L W.Biological stoichimometry from genes to ecosystems.Ecology letters,2000,3(6): 540–550. |
| [4] | Andersen T, Elser J J, Hessen D O.Stoichiometry and population dynamics.Ecology Letters,2004,7(9): 884–900. |
| [5] | He J S, Fang J Y, Wang Z H, Guo D L, Flynn D F B, Gengrg Z.Stoichiometry and large scale patterns of leaf carbon and nitrogen in the grassland biomes of China.Oecologia,2006,149(1): 115–122. |
| [6] | 潘复静, 张伟, 王克林, 何寻阳, 梁士楚, 韦国富.典型喀斯特峰丛洼地植被群落凋落物C:N:P生态化学计量特征.生态学报,2011,31(2): 335–343. |
| [7] | 张文彦, 樊江文, 钟华平, 胡中民, 宋璐璐, 王宁.中国典型草原优势植物功能群氮磷化学计量学特征研究.草地学报,2010,18(4): 503–509. |
| [8] | 仁书杰, 于贵瑞, 姜春明, 方华军, 孙晓敏.中国东部南北样带森林生态系统102个优势种叶片碳氮磷化学计量学统计特征.应用生态学报,2012,23(3): 581–586. |
| [9] | 牛得草, 李茜, 江世高, 常佩静, 傅华.阿拉善荒漠区6种主要灌木植物叶片C:N:P化学计量比的季节变化.植物生态学报,2013,37(4): 317–325. |
| [10] | 银晓瑞, 梁存柱, 王立新, 王炜, 刘钟龄, 刘小平.内蒙古典型草原不同恢复演替阶段植物养分化学计量学.植物生态学报,2010,34(1): 39–47. |
| [11] | 严正兵, 金南瑛, 韩廷申, 方精云, 韩文轩.氮磷施肥对拟南芥叶片碳氮磷化学计量特征的影响.植物生态学报,2013,37(6): 551–557. |
| [12] | Koerselman W, Meuleman A F M.The vegetation N:P ratio:a new tool to detect the nature of nutrient limitation.Journal of Applied Ecology,1996,33(6): 1441–1450. |
| [13] | Aerts R, de Caluwe H, Beltman B.Is the relation between nutrient supply and biodiversity co-determined by type of nutrient limitation? Oikos, 2003, 101(3):489-498.Oikos,2003,101(3): 489–498. |
| [14] | Güsewell, S, Bailey, K M, Ro em, W J, Bedford, B.L.Nutrient limitation and botanical diversity in wetlands:can fertilisation raise species richness? Oikos,2005,109(1): 71–80. |
| [15] | Venterink H O, Wassen M J, Verkroost A W M, de Ruiter P C.Species richness-productivity patterns differ between N-, P-, and K-limited wetlands.Ecology,2003,84(8): 2191–2199. |
| [16] | Braakhekke W G, Hooftman D A P.The resource balance hypothesis of plant species diversity in grassland.Journal of Vegetation Science,1999,10(2): 187–200. |
| [17] | 李瑞, 张克斌, 边振, 尤万学.宁夏哈巴湖自然保护区湿地景观多样性分析.干旱区资源与环境,2010,24(11): 139–143. |
| [18] | 鲍士旦. 土壤农化分析(.(第二版). 北京: 中国农业出版社, 2000. |
| [19] | 阎恩荣, 王希华, 周武.天童常绿阔叶林演替系列植物群落的N:P化学计量特征.植物生态学报,2008,32(1): 13–22. |
| [20] | Han W X, Fang J Y, Guo D L, Zhang Y.Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China.New Phytologist,2005,168(2): 377–385. |
| [21] | 曾冬萍, 蒋利玲, 曾从盛, 王维奇, 王纯.生态化学计量学特征及其应用研究进展.生态学报,2013,33(18): 5484–5492. |
| [22] | Tripathi N, Singh R S.Influence of different land uses on soil nitrogen transformations after conversion from an Indian dry tropical forest.Catena,2009,77(3): 216–223. |
| [23] | 刘万德, 苏建荣, 李帅锋, 张志钧, 李忠文.云南普洱季风常绿阔叶林演替系列植物和土壤C、N、P化学计量特征.生态学报,2010,30(23): 6581–6590. |
| [24] | 李征, 韩琳, 刘玉虹, 安树青, 冷欣.滨海盐地碱蓬不同生长阶段叶片C、N、P化学计量特征.植物生态学报,2012,36(10): 1054–1061. |
| [25] | 李玉霖, 毛伟, 赵学勇, 张铜会.北方典型荒漠及荒漠化地区植物叶片氮磷化学计量特征研究.环境科学,2010,31(8): 1716–1725. |
| [26] | 罗亚勇, 张宇, 张静辉, 卡召加, 尚伦宇, 王少影.不同退化阶段高寒草甸土壤化学计量特征.生态学杂志,2012,31(2): 254–260. |
| [27] | 张继义, 赵哈林, 张铜会, 赵学勇.科尔沁沙地植被恢复系列上群落演替与物种多样性的恢复动态.植物生态学报,2004,28(1): 86–92. |
| [28] | 赵雪莱, 何兴东, 薛苹苹, 张宁, 邬畏, 李荣, 慈华聪, 徐静静, 高玉葆, 赵哈林.土壤碳酸钙/有效磷比化学计量特征对油蒿群落植物密度的影响.科学通报,2012,57(1): 80–87. |
| [29] | Reich P B, Oleksyn J.Global patterns of plant leaf N and P in relation to temperature and latitude.Proceedings of the National Academy of Sciences of the United States of America,2004,101(30): 11001–11006. |
| [30] | Dowing J A.Marine Nitrogen:phosphorus stoichiometry and the global N:P cycle.Biogeochemistry,1997,37(3): 237–252. |
| [31] | 陈军强, 张蕊, 侯尧宸, 马丽娜, 丁路明, 龙瑞军, 尚占环.亚高山草甸植物群落物种多样性与群落C、N、P生态化学计量的关系.植物生态学报,2013,37(11): 979–987. |