 2016, Vol. 36
2016, Vol. 36文章信息
- 张雪妮, 杨晓东, 吕光辉
- ZHANG Xueni, YANG Xiaodong, LÜ Guanghui.
- 水盐梯度下荒漠植物多样性格局及其与土壤环境的关系
- Diversity patterns and response mechanisms of desert plants to the soil environment along soil water and salinity gradients
- 生态学报[J]. 2016, 36(11): 3206-3215
- Acta Ecologica Sinica[J]. 2016, 36(11): 3206-3215
- http://dx.doi.org/10.5846/stxb201505291082
-
文章历史
- 收稿日期: 2015-05-29
- 修订日期: 2015-10-22





2. 绿洲生态教育部重点实验室, 乌鲁木齐 830046;
3. 新疆大学资源与环境科学学院, 乌鲁木齐 830046
2. Key Laboratory of Oasis Ecology, Education Ministry, Urumqi 830046, China;
3. College of Resources and Environment Science, Xinjiang University, Urumqi 830046, China
植物多样性格局及其成因一直以来都是群落生态学研究的重点问题之一[1-4]。物种多度分布和多样性指数是研究群落多样性问题的基本工具,在某种程度上多度分布反映了物种对有限资源成功竞争的结果,能够阐明一定环境下决定集聚的生物多样性过程[5],而多样性指数是群落多样性的数量化表征,二者相辅相成,可综合反映群落结构和多样性格局。植物多样性格局的变化是多因素共同作用的过程,其中土壤环境是最直接、重要的因素之一,特别是在遭受旱胁迫、盐胁迫的干旱地区,土壤水分和盐分(水盐)是植物生长的主要制约因素,直接影响植被发生、多样性维持过程和群落演替等[6-7],决定了该区域植物多样性格局及其土壤环境影响的复杂性和特殊性[8]。因此,剖析干旱区荒漠植物多样性格局及其土壤环境影响规律,有助于阐明荒漠植物群落多样性过程机制,以及种群与环境相互作用的过程,对揭示干旱区植被资源恢复机理具有重要意义。
近年来,生物多样性锐减吸引了学界对多样性研究的高度关注[9],关于多样性格局及其影响机制亦积累了丰富成果。植物多样性格局和过程对环境因素的响应机制随着区域间气候、地形或土壤类型等而不同。目前,相关研究针对森林、草原等植被类型[10],从海拔高度[11]、纬度[12]以及尺度[13]等视角对不同植被类型的多样性格局响应机制展开了广泛分析,不同植被类型的多样性格局对各种梯度的响应不尽相同,未来有必要开展植物多样性格局和形成机制的深入和长期研究[11]。在干旱区,有关研究主要关注某一群落特征对单一环境梯度的响应,如水分或盐分梯度下植物群落组成[14-15],丰富度、多样性[16-17]的变化等,对水分和盐分双因子梯度下荒漠植物多度格局的响应机制尚缺乏深入探讨。干旱区土壤水盐关系密切[18],其动态直接影响土壤营养循环和生物学特征[19],进而影响植被结构和特征。因此,有必要对荒漠区水盐梯度下植物多样性格局的响应规律进行剖析,探讨物种在不同水盐梯度下与土壤环境间的作用机制,以期揭示荒漠区植物多样性的变化规律。
新疆艾比湖国家级湿地自然保护区属于典型的干旱荒漠区,位于保护区内的阿其克苏河河岸带土壤水盐含量较高,并随着离河道距离增大而降低[20]。另外,已有研究证明在干旱区随着离河道距离增加,植物种类分布和更替表现出一定的响应规律[21-22]。因此,在艾比湖保护区垂直阿其克苏河方向可开展土壤水盐梯度下荒漠植物多样性格局及影响机制研究,以期通过该研究回答以下科学问题:在自然土壤水盐双梯度下,(1) 荒漠植物多样性格局如何响应?(2) 土壤环境因子如何作用于植物组成和多样性变化,并最终形成特定水盐梯度下的多样性格局?解答上述问题,有助于了解荒漠植物群落结构沿土壤水盐梯度的变化规律及生物多样性维持机制,为揭示荒漠区植物多样性过程,了解干旱区生物多样性丧失机理[1]、恢复和管理植被资源、治理和修复土壤盐渍化等提供科学参考。
1 材料和方法 1.1 研究区概况艾比湖湿地国家级自然保护区(44°30′ —45°09′ N,82°36′ —83°50′ E)位于新疆精河县西北、准噶尔盆地西南,是准噶尔盆地西南缘最低洼地和水盐汇集中心。本区为典型大陆性气候,干燥少雨多风,年平均气温5℃。降水量年内分配不均,蒸发强烈,多年平均降水量和蒸发量分别为105.17 mm和1 315 mm。艾比湖特殊的湿地生态环境有其独特的生物资源多样性,主要植物种类包括近河岸的胡杨(Populus euphratica Oliv.)、柽柳(Tamarix ramosissima Ldb.)、梭梭(Haloxylon ammodendron (C. A. M.) Bge.)、盐豆木(Halimodendron halodendron (Pall.) Voss.)和芦苇(Phragmites australis (Cav.)Trin. ex Steud.),湖滨盐沼地的盐穗木(Halostachys caspica (M. B.) C. A. M.)、盐节木(Halocnemum strobilaceum (Pall.) M. B.)、碱蓬(Suaeda glauca (Bge.) Bge.)和盐爪爪(Kalidium foliatum (Pall.) Moq.),平原低地还分布有甘草(Glycyrrhiza uralensis Fisch.)、黑果枸杞(Lycium ruthenicum Murr.)、小獐毛(Aeluropus pungens (M. B.)C. Koch.),山前冲积洪扇有琵琶柴(Reaumuria soongorica (Pall.)Maxim.)等[23]。该区域分布有灰漠土、灰棕漠土和风沙土等典型地带性土壤,以及盐(盐渍化)土、草甸土和沼泽土等隐域性土壤。因干旱和盐碱作用,保护区土壤盐分高,碱性强,土壤质地较差,浅层(0—10 cm)土壤的平均电导率和pH值分别为5.41 ms/cm和8.77,平均容重约1.38 g/cm3,含水量平均为7.19%[24]。阿其克苏河位于保护区内湖区东侧,是艾比湖的水源之一,河岸及近处的古河道潜水溢出带分布有大量胡杨、柽柳、梭梭、盐豆木等典型植物,近年来,由于人类活动和气候变化的影响,阿其克苏河几近干涸,河岸植被面临退化威胁。
1.2 研究方法 1.2.1 样方调查、土壤采集与分析按土壤水盐可能沿离河距离的变化梯度,同时避开河道南岸的农业活动影响,选择在艾比湖湿地自然保护区阿其克苏河北侧垂直河道方向设置间隔约5 km的3条样带,样带涵盖区域可反映研究区局地尺度的水盐变化。各样带按离河距离增加向北部调查,各样带每隔500 m,设置1个10 m×10 m样方,样带和样方分布见图 1,试验中共调查样方32个。调查过程中记录样方位置和海拔,调查指标主要为物种数与个体数。同时,在调查样方内,随机采集5个裸地的0—15 cm土壤约500g,充分混匀后用四分法取土样约500g,同步利用TDR (Field ScoutTM TDR 300,Spectrum Technologies,Inc.,Plainifield,USA)在取土点附近测定土壤容积含水量;土样采集后置入自封袋,带回实验室自然风干后用于实验分析。室内分析的土壤指标和方法分别为:土壤pH值用酸度计法测定(PHS-3C,上海仪电科学仪器股份有限公司,上海,中国)、电导率采用电导率仪(DDS-307,上海仪电科学仪器股份有限公司,上海,中国)测定、有机碳(SOC)用重铬酸钾容量法、全氮(TN)用凯氏定氮法、全磷(TP)用HClO4-H2SO4消解-钼锑抗比浊法,全硫(TS)用Mg(NO3)2氧化-BaSO4比浊法,各指标测定方法参考文献[25]。

|
| 图 1 调查样地和采样点示意图 Fig. 1 Investigating plots and sampling sites |
土壤水分和盐分梯度划分通过聚类分析确定,在比较不同土壤水盐梯度下土壤各指标差异的基础上,分别分析各水盐梯度下植物种的多度分布、多样性变化,最后利用排序方法和逐步回归法分别分析物种和多样性指数与土壤因素间关系。上述聚类分析(类平均法)、方差分析(GLM,Duncan法多重比较)和逐步回归分析在SAS 8.0中完成;排序法首先用DCA分析选择排序方法,当排序轴梯度长度(Lengths of gradient,LGA)大于4时适合非线性模型,当LGA小于3时适合线性模型,LGA介于3和4之间时两种模型均适合[26]。SW1和SW2样地的物种DCA第一排序轴长度分别为4.765(>4) 和2.73(<3),因此分别采用CCA (单峰模型)和RDA(线性模型)分析两样地物种与土壤因子间关系,多样性指数的DCA分析显示LGA均小于3,因此采用RDA分析其与土壤环境间关系,所有排序分析在CANOCO 4.5中完成。植物多度分布拟合及α多样性计算利用R语言的vegan包完成,多样性指数的具体计算过程如下:
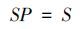
|
(1) |
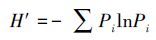
|
(2) |
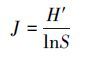
|
(3) |

|
(4) |
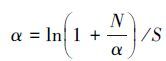
|
(5) |
式中,S为样方中的物种数,Pi为第i个种的多度占总多度的比例,N为总多度。
2 结果与分析 2.1 水盐梯度下植物组成和土壤因子差异32个样方划分至近河岸的高水盐样地(SW1)和距河岸2 km以上的低水盐样地(SW2)(图 1,表 1);两样地共记录植物26种,分属11科20属,SW1和SW2样地均出现20种植物,共有种14种 (表 2),不同样地的植物生活型组成存在差异(表 1),其中SW1样地灌木比例较高(12种),SW2样地灌木和草本比例相当(8、9种);与植物组成差异对应,SW1样地的土壤酸碱度 (pH)、电导率(EC)和容积含水量(SVWC)均极显著高于SW2样地(P<0.01) (表 1),反映了近河岸区域较好的土壤水分条件和较强的盐分胁迫;两样地土壤营养元素除全P外均有显著差异。
| 样地Plots | 乔,灌,草物种数量 Number of Tree, Shrub, Grass species | pH | 电导率 Electrical conductivity/(mS/cm) | 容积含水量 Soil volume water content/% | 有机碳 Soil organic carbon/ (g/kg) | 全氮 Total nitrogen/ (g/kg) | 全磷 Total phosphorus/ (g/kg) | 全硫 Total sulfur /(g/kg) |
| SW1(n=13) | 3,12,5 | 8.64a±0.38 | 12.02a±4.93 | 16.65a±2.89 | 4.45a±2.29 | 0.46a±0.18 | 0.46a±0.26 | 42.22a±11.34 |
| SW2(n=19) | 3,8,9 | 8.04b±0.36 | 1.91b±1.25 | 2.63b±2.32 | 1.19b±0.96 | 0.17b±0.10 | 0.40a±0.16 | 11.34b±12.28 |
| 同一指标数据的不同字母表示差异极显著,显著性水平P<0.01 | ||||||||
| 序号 Id | 植物种名 Plant Species name | 科属 Family and genera | 所属样地 Plot belongs to | ||||
| 1 | 梭梭 Haloxylon ammodendron | 藜科 梭梭属 | + | ||||
| 2 | 胡杨 Populus euphratica | 杨柳 科杨属 | + | ||||
| 3 | 骆驼刺 Alhagi sparsifolia | 豆科 骆驼刺属 | + | ||||
| 4 | 白刺 Nitraria schoberi | 蒺藜科 白刺属 | + | ||||
| 5 | 琵琶柴 Reaumuria soongorica | 柽柳科 琵琶柴属 | + | ||||
| 6 | 猪毛菜 Salsola collina | 藜科 猪毛菜属 | + | ||||
| 7 | 木碱蓬 Suaeda dendroides | 藜科 碱蓬属 | + | ||||
| 8 | 罗布麻 Apocynum venetum | 夹竹桃科 罗布麻属 | + | ||||
| 9 | 芦苇 Phragmites australis | 禾本科 芦苇属 | + | ||||
| 10 | 盐爪爪 Kalidium foliatum | 藜科 盐爪爪属 | + | ||||
| 11 | 花花柴 Karelinia caspica | 菊科 花花柴属 | + | ||||
| 12 | 柽柳 Tamarix ramosissima | 柽柳科 柽柳属 | + | ||||
| 13 | 盐豆木 Halimodendron halodendron | 豆科 盐豆木属 | + | ||||
| 14 | 盐节木 Halocnemum strobilaceum | 藜科 盐节木属 | + | ||||
| 15 | 甘草 Glycyrrhiza uralensis | 蝶形花科 甘草属 | SW1 | ||||
| 16 | 盐穗木 Halostachys caspica | 藜科 盐穗木属 | SW1 | ||||
| 17 | 盐地碱蓬 Suaeda salsa | 藜科 碱蓬属 | SW1 | ||||
| 18 | 平卧碱蓬 Suaeda prostrata | 藜科 碱蓬属 | SW1 | ||||
| 19 | 黑果枸杞 Lycium ruthenicum | 茄科 枸杞属 | SW1 | ||||
| 20 | 小叶碱蓬 Suaeda microphylla | 藜科 碱蓬属 | SW1 | ||||
| 21 | 刺沙蓬 Salsola ruthenica | 藜科 猪毛菜属 | SW2 | ||||
| 22 | 沙蓬 Agriophyllum squarrosum | 藜科 猪毛菜属 | SW2 | ||||
| 23 | 沙拐枣 Calligonum ebinuricum | 蓼科 沙拐枣属 | SW2 | ||||
| 24 | 对节刺 Horaninowia ulicina | 藜科 对节刺属 | SW2 | ||||
| 25 | 露果猪毛菜 Salsola aperta | 碱蓬科 猪毛菜属 | SW2 | ||||
| 26 | 绢蒿 Seriphidium santolinum | 菊科 绢蒿属 | SW2 | ||||
| 所属样地列中“+”号表示该物种在两个样地均出现 | |||||||
根据赤池信息准则(Akaike information criterion,AIC)越小拟合越佳的准则,SW1样地植物多度符合对数正态分布(生物统计模型)(图 2),富集种和稀有种分别3—4种,大多数种多度适中,反映了多数物种对高水盐生境的普遍适应性;SW2样地服从Zipf模型(生态位模型)分布(图 2),说明物种先后进入该生境的条件不同,后进入生境的物种需要付出较高代价才能侵入[27],可能属于逆境生存能力较强的种类;上述植物多度格局对不同水盐梯度的响应反映了植物多样性过程中的物种组成调整机制。

|
| 图 2 >SW1与SW2样地物种多度拟合曲线 Fig. 2 Plant abundance fitting curve of SW1 and SW2 plot |
两样地物种多样性差异总体显著(P<0.05)(图 3)。除Shannon-Wiener指数外,SW1样地多样性和均匀度均显著高于SW2样地(P<0.05);由于Shannon-Wiener和Simpson指数对常见种敏感,侧重群落优势度,Fisher-α参数对稀疏种敏感,侧重物种多度[28],因此上述差异反映出两样地常见种的优势度差异较小,而稀疏种的多度差异较大,这与两样地物种等级为5—15及两端时的多度特征相符,说明两类样地的多样性动态可能主要由各生境内的优势种和稀疏种主导。

|
| 图 3 SW1与SW2样地植物多样性差异 Fig. 3 Difference of plant diversity between SW1 and SW2 plots |
SW1样地CCA分析显示,第一、二轴特征值分别达到0.89和0.44,各排序轴上物种与土壤因子间相关性较高(>0.95),前两轴对物种-环境关系的累计方差贡献率达到62.5%; SW2样地RDA排序的前两轴特征值分别为0.62和0.05,物种与土壤环境间相关性达到0.73以上,第一、二排序轴对物种-环境间关系的累计方差贡献率达到94.7%以上(表 3)。上述说明土壤环境变量与物种间关系密切,土壤因素能够充分解释植物群落结构的变化。
| 项目 Item | 第一轴 Axis 1 | 第二轴 Axis 2 | 第一轴F检验 Axis 1 F test | |
| SW1 | 特征值 Eigenvalues | 0.89 | 0.44 | F=1.785 P=0.01 |
| CCA | 物种-环境相关性 Species-environment correlations | 0.98 | 0.95 | |
| 物种-环境关系的累计方差贡献率 Cumulative percentage variance of species-environment relation | 41.7 | 62.5 | ||
| SW2 | 特征值 Eigenvalues | 0.62 | 0.05 | F=18.02 P=0.01 |
| RDA | 物种-环境相关性 Species-environment correlations | 0.89 | 0.73 | |
| 物种-环境关系的累计方差贡献率 Cumulativepercentage variance of species-environment relation | 87.9 | 94.7 |
SW1样地第一、二排序轴分别与土壤S、pH、SVWC和P、C、N间有相对小的夹角,说明第一、二轴分别代表土壤酸碱度和水分以及土壤元素的变化梯度。植物种多集中于排序图中心,即高水盐生境下植物种普遍分布于土壤水盐和营养含量较低的区域;从物种水平看,盐地碱蓬(Suaeda salsa (L.)Pall.)、盐豆木、芦苇和琵琶柴等与土壤C、N、P营养间正相关,小叶碱蓬(Suaeda microphylla Pall.)和甘草与C、P营养间,以及盐爪爪、盐节木和猪毛菜(Salsola collina Pall.)与土壤水盐和营养间均为负相关(图 4)。SW2样地土壤SVWC与第一轴呈正相关,土壤C、P、N和pH与第二轴正相关,第一、二轴分别反映土壤水分和营养的变化。盐爪爪、盐节木、白刺(Nitraria schoberi L.)、骆驼刺(Alhagi sparsifolia (B. Keller et Shap.)Shap)和琵琶柴等分布于土壤水盐和C、P营养较高的区域;沙蓬(Agriophyllum squarrosum (L.)Moq.)、刺沙蓬(Salsola ruthenica Iljin.)、芦苇、花花柴(Karelinia caspica (Pall.) Less.)和盐豆木等分布于水盐偏低、营养适中但低S的土壤上;胡杨、猪毛菜等生长于土壤水盐和C、P略低的生境;而木碱蓬(Suaeda dendroides (C. A. M.)Moq.)、沙拐枣(Calligonum ebinuricum Ivanova.)、对节刺(Horaninowia ulicina Fisch. et Mey.)、露果猪毛菜(Salsola aperta Pauls.)、梭梭和绢蒿(Seriphidium santolinum (Schrenk)Poljak.)等则分布于土壤水盐和营养均较低的区域(图 4)。

|
| 图 4 SW1和SW2样地物种与土壤因子的CCA和RDA分析 Fig. 4 Ordination analysis on plant species and soil factors of SW1 (CCA) and SW2 (RDA) |
从多样性指数来看,土壤pH对高、低水盐生境下的植物多样性动态均有极显著影响,特别是对样地内种丰富度变化的调控作用显著;土壤SVWC和S营养仅在水盐较低生境下对植物多样性的变化有微弱的极显著影响;土壤EC及其他营养元素对研究区植物多样性的变化未表现出显著影响(表 4)。
| 指标 Item | 土壤pH pH | 土壤容积含水量 Soil volume water content /% | 土壤全硫 Total sulfur/ (g/kg) | R2 |
| 丰富度指数SP | 0.85**/0.67** | —/— | —/0.11** | 0.97/0.97 |
| Shannon-Wiener指数Shannon-Wiener | 0.15**/0.10** | —/-0.15** | —/0.03** | 0.94/0.86 |
| Pielou指数Pielou | —/0.06** | 0.04**/-0.08** | —/0.01** | 0.94/0.86 |
| Simpson指数Simpson | 0.07**/0.05** | —/-0.08** | —/0.01** | 0.94/0.84 |
| Fisher-α指数Fisher-α | 0.24**/0.11** | —/— | —/0.02** | 0.90/0.95 |
| “/” 前后分别表示SW1和SW2生境下的回归系数;**表示该指标在回归模型中极显著P<0.01; “—”表示无数据 | ||||
多样性指数与土壤因子间的RDA分析显示(图 5),SW1样地植物多样性指数与土壤pH和SVWC间多为正相关(除SP与SVWC外),物种丰富度SP与土壤S、P、EC间负相关,与土壤C、N间正相关;沿RDA第一排序轴从左到右,随着土壤S的降低和C、N的增加,物种丰富度增加;沿RDA第二排序轴从下到上,随着土壤pH、SVWC含量的降低,多样性指数逐渐降低;综合来看,土壤pH和SVWC是影响SW1 样地植物多样性变化的主要正向因子,抑制多样性增加的主要因子为土壤S。SW2样地植物多样性指数与土壤S间正相关,Shannon-Wiener指数、Simpson指数和Pielou指数与土壤pH间正相关;沿RDA第一排序轴从左至右,随着土壤S含量增加,多样性指数逐渐增大;沿RDA第二排序轴,从上至下随着pH的降低和土壤SVWC、EC、C、P等的增加,多样性指数下降;总体看来,土壤S、pH和SVWC是影响SW2样地多样性的决定性因子。

|
| 图 5 SW1和SW2样地物种多样性指数与土壤因子的RDA分析 Fig. 5 RDA analysis on plant species indices and soil factors of SW1 and SW2 |
物种多度分布模型的变化预示着生态学过程的转变[1]。研究区植物多度分布模型在高、低土壤水盐梯度间由对数正态分布模型转变为Zipf模型的变化说明,群落的组成结构对不同水盐条件产生响应。高水盐梯度下,物种多度呈对数正态分布模型表明,植物种丰度和多度结构可能受多种因素随机变化的影响[5],即在水分和盐分相对较高的生境,其他土壤因素对植物分布的决定作用同样不容忽视。低水盐梯度下物种多度分布符合Zipf模型,反映环境中多种因素的影响是依次作用于物种的,后来种的生存需求更多,因此比最初到达的种要少[5]。对于研究区的低水盐梯度,可以推测随着生境条件的愈加恶劣,植物间对资源的竞争逐渐凸显,具有不同生态位宽度的种规模产生分异,进而形成低水盐梯度下多度符合Zipf模型分布。从具体的多度和丰度变化来看,两样地多度较高(SW1>100,SW2>360)的植物种主要为草本,而部分草本以及灌木和乔木的多度较低(SW1和SW2<10)。同时,两样地总丰富度相等,但不同水盐梯度下灌木和草本植物丰富度的变化较大。
对于植物多样性,通常在一定范围内,其随着水分增加或盐分降低而提高[7, 16],但研究区高水盐生境下的植物多样性指数显著高于低水盐生境。这可能与植物对干旱和盐分胁迫的生态适应性有关。高水盐生境下,主要分布耐盐灌木或草本植物(如盐地碱蓬、小叶碱蓬、甘草、盐穗木等),而在土壤水分极度限制的低水盐生境下,仅分布有少量的耐旱草本或灌木(刺沙蓬、对节刺、沙拐枣、沙蓬等)。因此,旱胁迫加剧可能是导致两种生境下植物多样性指数差异显著的主要原因。
上述说明,土壤水盐及其他土壤因子对群落结构的影响主要通过草本植物及其多度调整来实现,研究区草本植物对浅层土水盐变化的敏感性较高,可在一定程度上指示土壤水盐梯度,为荒漠植被恢复和土壤盐渍化治理提供参考。在低水盐生境下,干旱胁迫是导致植物多样性显著降低的主要原因。
3.2 荒漠植物多样性格局与土壤因子间的关系不同水盐梯度下物种组成与土壤因子间关系的变化反映了群落结构的调整及其对生境变化的适应策略。在高水盐梯度下,仅在该样地分布的植物整体随着土壤S和pH、SVWC和EC的提高而增加,仅在SW2样地分布的物种则整体随着土壤因素的降低而增加,而对于两样地均出现的物种,其与土壤C、P、EC、SVWC的关系则在SW2样地更为密切。上述说明,群落通过耐盐或耐旱种的特殊分布来调整其结构,进而维持不同生境下群落的延续。同时,随着水盐的降低,SW2样地物种受土壤水分的限制更为突出。
研究区土壤C、N、P对不同水盐梯度下的植物多样性动态均未表现出显著影响,但土壤S元素在低水盐生境下对植物多样性动态的影响达到极显著水平。虽然在前人的研究中,有结论认为土壤营养对生物多样性没有影响[29],然而,艾比湖保护区位于典型的荒漠区,土壤贫瘠干旱,营养循环缓慢,利用率低[30],植物生长和分布普遍遭受N、P营养的限制[31],N、P对植物多样性无显著影响说明,一方面,干旱可能影响植物吸收P的能力[30],并且N限制随着P限制而加剧[32],研究区植物承受的干旱胁迫影响可能大于N、P限制的效应。另一方面,在植物演替的稳定时期,其对氮和磷的需求较低[30],研究区植物群落组成相对稳定(共有种14种),这一时期植物对氮、磷的需求也可能稳定在较低水平。S对研究区植物多样性的显著影响可能是由于其在植物的生长凋节、解毒、防卫和抗逆等过程中起重要作用[33];干旱区含盐分、石膏和石灰较高的荒漠土壤S含量亦较高[34],有研究表明钙质土施S可显著提高土壤中P的有效性,同时S还可以促进N 的利用效率[35]。因此,土壤S对提高研究区植物多样性可能具有一定的关键作用,特别是在低水盐生境下土壤S与部分特有植物关系紧密,说明土壤S对某些荒漠耐旱植物种的生长影响较大,其作用应在荒漠区生物多样性保护中予以重视。
| [1] | 马克明.物种多度格局研究进展.植物生态学报,2003,27(3): 412–426. |
| [2] | 韩大勇, 杨允菲.松嫩草地植物群落物种多度-分布关系及其解释.生物多样性,2014,22(3): 348–357. |
| [3] | Lefcheck J S, Byrnes J E, Isbell F, Gamfeldt L, Grifın J N, Eisenhauer N, Hensel M J S, Hector A, Cardinale B J, Duffy J E.Biodiversity enhances ecosystem multifunctionality across trophic levels and habitats.Nature Communications,2015,6 6936 |
| [4] | 李旭华, 邓永利, 张峰, 董刚, 李世广.山西庞泉沟自然保护区森林群落物种多样性.生态学杂志,2013,32(7): 1667–1673. |
| [5] | Magurran A E. 生物多样性测量. 张峰, 译. 北京: 科学出版社, 2013. |
| [6] | 李新荣, 何明珠, 贾荣亮.黑河中下游荒漠区植物多样性分布对土壤水分变化的响应.地球科学进展,2008,23(7): 685–691. |
| [7] | 王水献, 董新光, 刘磊.新疆焉耆盆地绿洲水盐双梯度下天然植被多样性分异特征.冰川冻土,2010,32(5): 999–1006. |
| [8] | 杜泉滢, 李智, 刘书润, 刘鸿雁.干旱、半干旱区湖泊周围盐生植物群落的多样性格局及特点.生物多样性,2007,15(3): 271–281. |
| [9] | Hautier Y, Tilman D, Isbell F, Seabloom E W, Borer E T, Reich P B.Anthropogenic environmental changes affect ecosystem stability via biodiversity.Science,2015,348(6232): 336–340. |
| [10] | 夏富才, 潘春芳, 赵秀海, 何海燕, 周海城.长白山原始阔叶红松林林下草本植物多样性格局及其影响因素.西北植物学报,2012,32(2): 370–376. |
| [11] | Sproull G J, Quigley M F, Sher A, González E.Long-term changes in composition, diversity and distribution patterns in four herbaceous plant communities along an elevational gradient.Journal of Vegetation Science,2015,26(3): 552–563. |
| [12] | Visser V, Clayton W D, Simpson D A, Freckleton R P, Osborne C P.Mechanisms driving an unusual latitudinal diversity gradient for grasses.Global Ecology and Biogeography,2014,23(1): 61–75. |
| [13] | 要伊桐, 元正龙, 魏彦波, 赵秀海.长白山次生杨桦林树种多样性格局多尺度分析.北京林业大学学报,2014,36(6): 86–92. |
| [14] | Naz N, Hameed M, Ahmad M S A, Ashraf M, Arshad M.Is soil salinity one of the major determinants of community structure under arid environments? Community Ecology, 2010, 11(1): 84-90.Community Ecology,2010,11(1): 84–90. |
| [15] | Naz N, Hameed M, Nawaz T, Ahmad M S A, Ashraf M.Soil-plant relationships in the arid saline desert of Cholistan.Arid Land Research and Management,2013,27(2): 140–152. |
| [16] | 张雪妮, 吕光辉, 杨晓东, 贡璐, 秦璐, 何学敏, 刘昊奇.基于盐分梯度的荒漠植物多样性与群落、种间联接响应.生态学报,2013,33(18): 5714–5722. |
| [17] | 靳朝, 雷霆.水分梯度下洞庭湖洲滩植物群落多样性特征.湿地科学,2015,13(2): 177–183. |
| [18] | 王雪梅, 柴仲平, 塔西甫拉提·特依拜, 龚爱谨, 程红梅.干旱区土壤盐渍化及其影响因子分析-以渭干河-库车河三角洲绿洲为例.土壤,2009,41(3): 477–482. |
| [19] | 徐万里, 刘骅, 张云舒.新疆盐渍化土壤氮素矿化和硝化作用特征.西北农林科技大学学报 (自然科学版),2007,35(11): 141–145. |
| [20] | 张雪妮, 吕光辉, 王庭权, 马玉, 阿布里孜·阿不都热合曼, 赵晓英, 郭振洁, 朱修逸.荒漠区垂直河岸带植物多样性格局及其成因.生态学报,2015,35(18): 5966–5974. |
| [21] | 陈亚宁, 张宏锋, 李卫红, 陈亚鹏.新疆塔里木河下游物种多样性变化与地下水位的关系.地球科学进展,2005,20(2): 158–165. |
| [22] | 刘加珍, 李卫红, 吴纯渊, 陈永金.荒漠河岸生态系统退化与物种多样性恢复研究.干旱区研究,2009,26(2): 212–220. |
| [23] | 杨晓东, 吕光辉, 田幼华, 杨军, 张雪梅.新疆艾比湖湿地自然保护区植物的生态分组.生态学杂志,2009,28(12): 2489–2494. |
| [24] | 张雪妮. 荒漠植物群落演替及其营养元素驱动研究. 乌鲁木齐: 新疆大学, 2014. |
| [25] | 南京农业大学. 土壤农化分析. 北京: 农业出版社, 1988. |
| [26] | Lepš J, Šmilauer P. Multivariate Analysis of Ecological Data Using CANOCO. Cambridge: Cambridge University Press, 200350–51. |
| [27] | 彭少麟, 殷祚云, 任海, 郭勤峰.多物种集合的种-多度关系模型研究进展.生态学报,2003,23(8): 1590–1605. |
| [28] | 马克平.生物群落多样性的测度方法Ⅰα多样性的测度方法(上).生物多样性,1994,2(3): 162–168. |
| [29] | 王长庭, 龙瑞军, 王启兰, 曹广民, 施建军, 杜岩功.放牧扰动下高寒草甸植物多样性、生产力对土壤养分条件变化的响应.生态学报,2008,28(9): 4144–4152. |
| [30] | 邬畏, 何兴东, 张宁, 王海涛, 马迪.油蒿群落演替中的植物密度对土壤氮/磷比的响应.土壤学报,2009,46(3): 472–479. |
| [31] | 张珂, 何明珠, 李新荣, 谭会娟, 高艳红, 李刚, 韩国君, 吴杨杨.阿拉善荒漠典型植物叶片碳、氮、磷化学计量特征.生态学报,2014,34(22): 6538–6547. |
| [32] | Marleau J N, Jin Y, Bishop J G, Fagan W F, Lewis M A.A stoichiometric model of early plant primary succession.The American Naturalist,2011,177(2): 233–245. |
| [33] | 谢瑞芝, 董树亭, 胡昌浩.植物硫素营养研究进展.中国农学通报,2002,18(2): 65–69. |
| [34] | Sardans J, Rivas-Ubach A, Peñuelas J.The elemental stoichiometry of aquatic and terrestrial ecosystems and its relationships with organismic lifestyle and ecosystem structure and function: a review and perspectives.Biogeochemistry,2012,111(1/3): 1–39. |
| [35] | Legay N, Personeni E, Slezack-Deschaumes S, Piutti S, Cliquet J B.Grassland species show similar strategies for sulphur and nitrogen acquisition.Plant and Soil,2014,375(1/2): 113–126. |