 2016, Vol. 36
2016, Vol. 36文章信息
- 孟鹏, 安宇宁, 白雪峰
- MENG Peng, AN Yuning, BAI Xuefeng.
- 沙地赤松光合及叶绿素a快相荧光动力学特性
- Photosynthetic characteristics and chlorophyll a fluorescence induction parameters of Pinus densiflora on sandy soil
- 生态学报[J]. 2016, 36(11): 3469-3478
- Acta Ecologica Sinica[J]. 2016, 36(11): 3469-3478
- http://dx.doi.org/10.5846/stxb201503230552
-
文章历史
- 收稿日期: 2015-03-23
- 修订日期: 2015-11-30

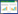




以CO2气体交换为特征的光合作用是植物的重要生理过程之一,植物的光合性能反映着其对光能的吸收与转换能力,与生长和生产力水平密切相关。叶绿素荧光分析技术是用于光合作用机理研究的一种“内在探针”,与 “表观性”的气体交换指标相比,叶绿素荧光参数更具有反映“内在性”的特点[3],这种方法被称为JIP-测定[5]。以该技术为切入点研究一些松属树种光合作用内在机理已有报道,比如Busch 等探讨了班克松(P. banksiana) 的能量耗散机制[7],Pollastrini等探讨了意大利松(P. pinea)在遭受胁迫的初期针叶内电子传递效率和光合性能[9]。沙地赤松(P.densiflora)由辽宁省固沙造林研究所(下文简称固沙所)于1965年开始陆续在科尔沁沙地南缘引种栽培,至今已有50年的引种历史,于2014年被辽宁省林木良种审定委员会审定为赤松优良新品种。20世纪90年代开始,章古台沙地个别地点引种樟子松林(P. sylvestris var.mongolica)就出现了衰退现象,遭受了松枯梢病,外在表现为从底轮枝开始针叶枯黄萎蔫,而同期相同地点引种的沙地赤松则未表现任何衰退特征,感病指数几乎为0[11]。在光适应范围研究方面,Han QM等人较全面的测定了赤松一级枝在水平和垂直方向上的光梯度,从叶片发育过程中形态指标的改变揭示了成熟赤松具有较强的光适应性[2],但该研究没有从叶绿素荧光角度深入探讨光适应性的机理。综上,针对特殊自然生境下赤松成熟林分的研究较少,沙地赤松成熟林分的光合及叶绿素a快相荧光动力学特性研究还未见报道。
本研究以同龄樟子松为对照,在对成林阶段沙地赤松生长指标进行调查的基础上,采用光合及JIP-测定相结合的方法对比二者的光合指标,并深入了解沙地赤松的光合机理,为在沙地环境下推广应用该针叶树良种提供一些基础的理论依据。
1 材料与方法 1.1 研究区概况位于辽宁省彰武县章古台镇固沙所三家子实验区,地处42°43′N,122°29′E,海拔226.5 m,年均降水量为433 mm,蒸发量1570 mm,年均温6.7 ℃,无霜期154 d,≥10 ℃有效积温2800—3200 ℃。
1.1.1 试验林分试验林分建于1976年4月上旬,通过引种栽植2年生实生苗形成了沙地赤松与樟子松两个同龄的相邻林分,林分间相距6.0m。两林分面积分别为2.7hm2(沙地赤松)和2.1 hm2(樟子松),初植株行距为1.0m×3.0m,后经历了相同的抚育和营林措施,于2002年进行了一次间伐,株行距变化为2.0m×6.0m。此后由于樟子松遭到了病虫害,对受害严重的单株进行了清除,因此樟子松林密度(701株/hm2)略低于沙地赤松(803株/hm2)。栽植地点为平缓沙地,高程差不超过0.4m。
1.1.2 土壤试验林建立时,土壤为灰沙土,瘠薄干旱,腐殖质含量极低,pH值呈中性。土壤表层20—50cm为灰沙,其下埋有50—100cm不等的黑褐色沙土,最下面则为白沙,沙层厚30 m左右。土壤地下水位为3.6m,0—50cm土层土壤含水率年变化很大,在4.0%—8.0%之间。
开展本次研究时,0—50cm土层土壤含水率年变化进一步加大,在1.5%—9.5%之间,土壤地下水位下降为5.0m。两林分间土壤水分条件没有显著差异。
1.1.3 植被试验林建立时该区植被稀疏,均为一般沙地常见的旱生植物,有的形成群落,有的则不规律的散生,物种包括黄柳(Salix gordejevii)、胡枝子(Lespedeza bicolor)、差巴嘎蒿(Artemisia halodendron)、拂子矛(Calamagrostis epigeios)、碱草(Elymus dahuricus)等。
开展本次研究时,该区植被类型有所增加,主要是以樟子松、油松(P.tabuliformis)、沙地赤松为主的固沙针叶林,林下有少量伴生植物,包括鼠李(Rhumus parvifolius)、榆树(Ulmus pumila)、桑树(Morus alba)、胡枝子、南蛇藤(Celastrus articulatus)、白蔹(Ampelopsis japonica)、锦鸡儿(Caragana microphylla)、山里红(Crataegus pinnatifida)、黄菠萝(Phellodenron amurense)、小叶朴(Celtes bungeana)、小叶杨(Populus simoni)等。
1.2 生长量和生物量调查于2013年8月上旬,在试验林内分树种各设置1块30m×30m标准地,对林分密度、树高、胸径等指标进行调查。在各自标准地内,根据平均胸径、平均树高,选择3株标准木。在测定完光合及叶绿素荧光指标后,于8月中旬将标准木伐倒,利用解析木法测定连年树高和胸径生长量,并计算连年单株材积生长量。采用分层切割法测定干、枝、叶、球果等地上生物量[14],为了避免细根的损失,挖了长8m、宽6m、深4m的坑,用自制的三齿耙十分小心的从根尖部向根基部追踪式挖掘,根部挖出后用自来水洗净,然后烘干测生物量。各器官生物量测定后,计算干叶比(干生物量与叶生物量比值)和根茎比(根生物量与地上生物量比值)。
1.3 光合特性调查测定时间为8月上旬。在每株所选标准木树冠阳面中部选择样枝并做标记,用Li-6400光合测定系统,在不加任何人为环境控制的自然状态下测定两树种当年生针叶净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)等指标。将被测针叶尽量摆满叶室,表面积由针叶长度和表面积形成的回归方程来确定[5]。测定时选择晴朗天气,测定3d,每天8:00开始测定,每2h测1次,18:00结束,每个时间点每株标准木重复测定3次。测定期内光合有效辐射(PAR)变化范围为52—1704μmol m-2 s-1,均值为839.5μmol m-2 s-1;气温变化范围为22—29℃,均值为26.7℃;相对湿度(RH)变化范围为45%—73%,均值为59.2%。由于测定枝距地面4m左右,所以测定时需将叶室架设于4m高铝合金人字梯上。水分利用效率(WUE)由Pn与Tr的比值求得。
光响应曲线参数的测定:在测定光合日变化的当日,于10:00叶片充分活化时进行光响应参数测定。测定时叶室水平放置。在正常大气CO2浓度、最适温度范围、RH(70±5)%条件下,利用Li-6400光合测定系统自带的红蓝光源测定,光量子通量密度(PPFD)设置为2000、1800、1500、1200、1000、800、500、200、100、80、50、20、0μmol m-2 s-1,设置最小等待时间为120s,最大等待时间为200s,测定不同PPFD下针叶的Pn。两个树种Pn与PPFD之间的关系可用SPSS V17.0软件Nonlinear模块,按下述经验方程进行拟合[6]:
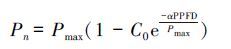
|
(1) |
式中,Pmax(μmolCO2 m-2 s-1)为最大净光合速率; α为弱光下光量子利用效率(即表观量子效率,AQY); C0为度量弱光下净光合速率趋近于零的指标。当PPFD为0时,Pn为负值,即是暗呼吸速率(Rd),通过公式(1)可以计算。通过适合性检验,若方程拟合效果良好,则可用下式计算光补偿点(LCP):
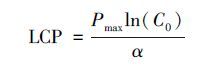
|
(2) |
假定Pn达到Pmax的99%时的PPFD为光饱和点(LSP),则:
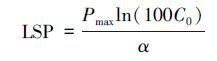
|
(3) |
与光合指标同期测定。具体方法是于8:00 剪取光合测定时所选样枝附近见光良好、粗细均匀的当年生枝5条,迅速插入备好的水瓶中,并装入放有湿巾的黑塑料袋内,立即带回实验室,暗适应30min。然后用Pocket PEA植物效率分析仪 (型号PPEA,Handstech Instrument Ltd. 英国)测定叶绿素a快相荧光的多相跃升,并读取相关参数[17]。当年生针叶平行排列在荧光夹中,完全覆盖4 mm2的测试孔。测定光源为6 个发光二极管提供的波长为650 nm 的红光,光照强度为3500μmol m-2 s-1的饱和光强,荧光信号的记录时程为1s,起始点为照光后10μs,每种针叶测定20 个重复。Pocket PEA可以产生以时间为横坐标,原始荧光值(从最小荧光Fo至最大荧光Fm)为纵坐标的叶绿素快相荧光(OJIP)曲线。为对比树种间差异,克服原始OJIP 曲线变异性较大的缺点,对原始曲线进行了标准化。本研究用Fm-Fo 进行标准化,标准化的荧光信号数据用相对可变荧光(Vt)表示,Vt =(Ft -Fo) / (Fm-Fo),用Vt绘制的OJIP 曲线O 相的荧光强度均为0,P 相的荧光强度均为1[18]。解析从OJIP曲线中得到的主要荧光参数的生理意义(表 1)。
| 参数 Parameter | 涵义 Meaning | 参数 Parameter | 涵义 Meaning |
| ABS/RC | 单位反应中心吸收的光量 | ETo/CSo | 单位激发态面积用于电子传递的能量 |
| TRo/RC | 被单位反应中心捕获的光量 | DIo/CSo | 单位激发态面积热耗散的能量 |
| ETo/RC | 单位反应中心用于电子传递的能量 | ABS/CSm | 单位激发态面积吸收的光量(以最大荧光为基础) |
| DIo/RC | 单位反应中心热耗散的能量 | RC/CSm | 单位激发态面积反应中心数目(以最大荧光为基础) |
| RC/CSo | 单位激发态面积反应中心数目 | PIABS | 以吸收光能(ABS)为基础的性能指数 |
| ABS/CSo | 单位激发态面积吸收的光量 | PICSo | 以基础荧光(Fo)为基础的性能指数 |
| TRo/CSo | 单位激发态面积被反应中心捕获的光量 | PICSm | 以最大荧光(Fm)为基础的性能指数 |
| ABS:吸收的光量Absorption flux; RC:反应中心 Reaction center; TR:捕获的光量Trapped energy flux; ET:电子传递的能量Electron transport flux; DI: 热耗散的能量Dissipated energy flux; CS: 激发态面积Excited cross section; PI:性能指数Performance index | |||
采用混合液法测定[19]。
1.6 数据处理所有数据均采用SPSSV17.0软件进行数理统计分析,对组间数据进行单因素方差分析(one-way ANOVA),采用Fisher的LSD法做多重比较分析;显著性水平设定为α=0.05。采用Excel 2003制图。
2 结果与分析 2.1 沙地赤松与樟子松单株生长量及生物量对比 2.1.1 生长量对标准地内沙地赤松和樟子松树高和胸径生长量进行对比(表 2),发现沙地赤松树高较高,是樟子松的1.15倍;胸径较大,是樟子松的1.23倍。而且,无论是树高还是胸径指标,沙地赤松的变异系数均较小,表明沙地赤松林分生长离散度较小,单株间生长量更趋于一致。进行本次研究的当年及前一年,沙地赤松的连年单株材积生长量均显著高于樟子松。
| 树种 Spieces | 树高 Tree height/m | 胸径 DBH/cm | 连年材积生长量/m3Current annual volume increment | |||||
| 平均值 Mean value | 变异系数 CV | 平均值 Mean value | 变异系数 CV | 2012年 | 2013年 | |||
| 沙地赤松P. densiflora | 9.36±0.69a | 0.074 | 18.44±2.50a | 0.136 | 0.0036±0.0004a | 0.0048±0.0005a | ||
| 樟子松P. sylvestris var. mongolica | 8.17±1.15b | 0.141 | 14.99±2.50b | 0.167 | 0.0017±0.0004b | 0.0011±0.0002b | ||
| 不同小写字母表示不同树种间差异显著;表中数据为平均值±标准差,在树高和胸径指标方面,沙地赤松n=69,樟子松n=74;连年材积生长量指标方面,两树种n=3 | ||||||||
表 3所示,两树种生物量排序规律是一致的,即:干>根>枝>叶>果。沙地赤松总生物量较大,是同龄樟子松的1.73倍,其中干、果、根生物量显著高于樟子松,分别是樟子松的1.69、7.8和1.97倍,而枝、叶生物量与樟子松差异不显著。沙地赤松干叶比和根茎比均显著高于樟子松。
| 树种 Spieces | 生物量 Biomass/(kg/株) | ||||||
| 干 Stem | 枝 Branch | 叶 Needle | 果 Cone | 根 Root | 干叶比 Stem∶Needle | 根茎比 Root∶Shoot | |
| 沙地赤松P. densiflora | 60.38±10.05a | 19.16±3.74a | 6.86±2.07a | 0.39±0.14a | 35.58±5.01a | 8.80±2.56a | 0.41±0.06a |
| 樟子松P. sylvestris var. mongolica | 35.81±8.88b | 9.78±5.37a | 7.03±4.48a | 0.05±0.04b | 18.10±9.57b | 5.09±1.47b | 0.34±0.08b |
| 不同小写字母表示不同树种间差异显著;表中数据为平均值±标准差,n=3 | |||||||
光响应曲线反映了在其它环境因子相对稳定的条件下,植物Pn随PPFD变化的规律。利用公式(1)拟合出两树种的光响应曲线(图 1右),拟合公式如下:

|
| 图 1 两树种利用公式(1)拟合前(a)和拟合后(b)的光响应曲线 Fig. 1 PPFD response curves of net photosynthetic rate (Pn) for the two species before (a) and after (b) fitted by equation (1) |
沙地赤松
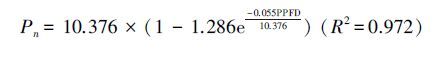
|
樟子松
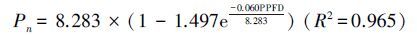
|
根据公式1—3计算出各光响应参数(表 4)。Pmax/Rd的比值反应植物的光合效率,沙地赤松Pmax/Rd较高,是樟子松的1.74倍。沙地赤松Pmax较高,是樟子松的1.25倍,且Rd较低,导致Pmax/Rd较高。由光响应曲线可以看出,沙地赤松在几乎任意光强下Pn均大于樟子松(图 1)。光合日变化可以看出,沙地赤松Pn日变化范围在3.290—7.349μmolCO2 m-2 s-1之间,平均Pn为4.902μmol CO2 m-2 s-1,比樟子松大36.2%,除14:00和16:00外其他时间点均显著超过樟子松(表 5)。另外沙地赤松Tr比樟子松小34.8%,因此水分利用效率(WUE)显著高于樟子松,约是樟子松的2.06倍。并且,沙地赤松能在较小Gs下保持较高Pn (表 6)。
| 树种 Spieces | LSP/ (μmolCO2 m-2 s-1) | LCP/ (μmol CO2 m-2 s-1) | AQY | Pmax/ (μmol CO2 m-2 s-1) | Rd/ (μmol CO2 m-2 s-1) | Pmax/Rd | Chl/ (mg/g) |
| 沙地赤松P. densiflora | 916.240 | 47.453 | 0.055 | 10.376 | 2.968 | 3.496 | 1.380 |
| 樟子松P. sylvestris var. mongolica | 691.442 | 55.698 | 0.060 | 8.283 | 4.116 | 2.012 | 0.694 |
| 树种Tree species | Pn /(μmolCO2 m-2 s-1) | |||||
| 8:00 | 10:00 | 12:00 | 14:00 | 16:00 | 18:00 | |
| 沙地赤松P. densiflora | 7.349±0.618a | 6.211±0.800a | 4.001±0.776a | 3.931±0.841a | 4.631±0.646a | 3.290±1.207a |
| 樟子松P. sylvestris var. mongolica | 5.365±1.412b | 5.441±1.174b | 2.293±0.828b | 3.783±0.646a | 3.523±0.778a | 1.190±0.395b |
| 表中数值平均值±样本标准差;每个时间点随机取20个数据,即n=20;a,b表示不同树种间差异显著 | ||||||
| 树种Tree species | Gs /(mmolH2O m - 2 s-1) | Pn /(μmolCO2 m-2 s-1) | Tr /(mmolH2O m-2 s-1) | 水分利用效率WUE/(μmol CO2/mmol H2O) |
| 沙地赤松P. densiflora | 199.828±34.424a | 4.902±1.640a | 4.866±1.336a | 1.005±0.390a |
| 樟子松 P. sylvestris var. mongolica | 225.820±16.702b | 3.599±1.797b | 7.458±0.737b | 0.488±0.379b |
| 表中数值平均值±样本标准差;每个时间点随机取20个数据,所以n=120;a,b表示不同树种间差异显著 | ||||
沙地赤松具有更高的光饱和点(LSP)和更低的光补偿点(LCP)(表 4)。通过实际光响应曲线(图 1a),发现樟子松易于发生动态光抑制(dynamic photoinhibition),在PPFD达1800μmol m-2 s-1时,光合速率迅速下降,Pn为峰值的77.4%;而当PPFD达2000μmol m-2 s-1时,Pn仅为峰值的30.5%。相比之下,沙地赤松在PPFD为1800μmol m-2 s-1时,并未发生光抑制,当PPFD达2000μmol m-2 s-1时,仅发生了轻微的光抑制,Pn为峰值的91.4%。本研究还发现沙地赤松针叶具有较高的Chl,比樟子松高1倍,达到1.380mg/g(表 4)。
2.3 叶绿素a快相荧光动力学特征对比 2.3.1 特征位点比较由Vt 绘制的OJIP 曲线可以看出(图 2),两树种均为典型的叶绿素快相荧光动力学曲线,具有O、J、I、P等相点。它们的曲线形态比较相似,但到达P相点的时间(Tfm)不相同,沙地赤松为700ms(P1位点),而樟子松为900ms(P2位点)。该图亦可看出,两树种针叶OJIP 曲线的差异主要在J 相(2ms)附近。为更清楚的观察包括J 相在内各相荧光的差异,以樟子松Vt 绘制的OJIP 曲线为对照,然后用沙地赤松曲线与对照曲线的标准化荧光差值(ΔVt)重新作图(图 2)。结果显示,从O相到接近P相的过程中,沙地赤松标准化荧光差值(ΔVt)大部分时间点均低于樟子松,两树种差异主要出现在300μs—30ms范围内,即J相附近区域,其中最大差异出现在J—I相之间(时间约为104μs)。在快接近P相时,沙地赤松标准化荧光值出现了高于樟子松的一个窄峰。

|
| 图 2 沙地赤松与樟子松由Vt和ΔVt绘制的叶绿素快相荧光动力学曲线 Fig. 2 Fast chlorophyll fluorescence transients of P. densiflora and P. sylvestris var. mongolica based on Vt and ΔVt |
比活性参数是指活跃的单位反应中心(RC)或单位受光面积(CS)的各种量子效率(ABS/RC、TRo/RC、ETo/RC、DIo/RC、ABS/CSo、TRo/CSo、ETo/CSo、DIo/CSo、ABS/CSm)以及单位面积上的反应中心的数量(RC/CSo、RC/CSm)(表 1)。图 3 是以樟子松的荧光参数作为对照(参数值为1),用沙地赤松的荧光参数与对照参数的比值绘制成的雷达图。由图 3可见除ABS/RC与DIo/RC小于樟子松外,沙地赤松其它比活性参数指标均等于或大于樟子松,尤其是RC/CSo、RC/CSm、ABS/CSo、ABS/CSm、TRo/CSo、ETo/CSo显著高于樟子松。图 3还表明,沙地赤松性能指数PI(AB/CSo/CSm) 较高,分别是樟子松的1.42、1.65和1.63倍。

|
| 图 3 沙地赤松与樟子松叶绿素a荧光比较 Fig. 3 Radar plot of fluorescence data of P. densiflora and P. sylvestris var. mongolica |
固沙所从1965年开始就陆续在科尔沁沙地南缘地区开展了沙地赤松引种造林试验,目前沙地赤松林树势健康,生长稳定。通过本研究发现在同样立地条件下,虽然沙地赤松密度较大,单株营养面积和受光照条件不及樟子松,但其单株生长量和生物量均大于樟子松,尤其是在测定光合指标的当年及前一年的连年材积生长量显著大于樟子松,说明沙地赤松可以固定较多的CO2,并积累较多的光合产物用于生长。由于植物固定CO2的能力和其光合性能及吸收光能后的电子传递具有相关性[20],推断沙地赤松光合性能和光能利用效率可能好于樟子松。
为验证这一推论,本研究采用光合生理指标和叶绿素a快相荧光动力学指标相结合的方法,对沙地赤松光合水分生理特性进行了剖析。研究发现沙地赤松具有较强的光合能力,Pmax和平均Pn均高于樟子松,能在较小Gs下保持较高Pn。这意味着沙地赤松可以产生较多的光合产物。在光合产物分配方面,沙地赤松也具有自己的优势:干叶比较高,说明其把更多的光合产物积累到了树干中,也因此连年材积生长较高;根茎比较高,应归因于沙地赤松较强的光合能力导致地上部分能够向根系供给充足的碳氢化合物[21],用于了根系的横向和垂直向的生长[22],提高了自身的吸水能力和抗旱性,也因此更有利于适应土壤长期水分亏缺的沙地环境。沙地环境的另一个特点是多风,在章古台地区树木生长季常以南及西南风为主,最大瞬间风速32m/s,年5m/s以上的风可达240次。在这样的多风环境下,植物的蒸腾耗水量增加,极易使植物处于干旱胁迫之下。而沙地赤松能够保持较小Tr和较高的WUE,增加了其在沙地环境下长期生存生长的稳定性。另外暗呼吸速率(Rd)较低保证了沙地赤松对光合产物的低消耗[23],同时形成了较高的光合效率(Pmax/Rd)。与樟子松相比,沙地赤松具有更高的LSP,因此在较高光强(2000μmol m-2 s-1)下仅发生轻微的光抑制;具有更低的LCP,说明其具有较强的利用弱光的能力,这和其较高的叶绿素含量(Chl)有关,较高的Chl有利于叶绿体对光能的吸收与转化,也保障了其较高的光合能力[24]。
JIP-测定表明,从O相到接近P相的过程中,沙地赤松标准化荧光值大部分时间点均低于樟子松,其中显著差异出现在J相附近和J—I相之间,说明沙地赤松PSⅡ反应中心捕获的能量中用于电子传递的比例较高,在电子越过初级醌受体A(QA)的过程中及从Q-A向次级醌受体(QB)传递过程中耗散能量较小。
前人研究表明,比活性参数可以更确切的反映植物的光合器官对光能的吸收、转化和耗散等状况[25],长期多次干旱胁迫会造成沙地植物在ETo/CSo等指标的下降,从而降低用于电子传递的能量[26]。本研究发现沙地赤松大部分比活性参数高于樟子松,包括RC/CSo、RC/CSm、ABS/CSo、ABS/CSm、TRo/ CSo、ETo/CSo等,表明其在沙地环境条件下单位激发态面积反应中心数目(RC/CSo、RC/CSm)、吸收的光量(ABS/CSo、ABS/CSm)、被反应中心捕获的光量(TRo/CSo)、用于电子传递的能量(ETo/CSo)均保持较高水平。性能指数(PI(ABS/CSo/CSm))包含了3 个参数[RC/CS、φPo(PSⅡ 最大光化学效率) 和ψo(表示电子越过QA的能量占反应中心捕获能量的比例)],这3 个参数相互独立[27],所以性能指数和推动力(即性能指数的对数)可以更准确地反映植物光合机构的状态[28]。沙地赤松性能指数PI(ABS/CSo/CSm) 较高,证明沙地赤松光反应活性较强。沙地赤松光合机构的这些优良特性使其能在较小Gs下保持较高Pn,也使其在弱光下能够吸收更多的光能,导致较低的LCP;在强光下能利用较多的光能,从而避免激发能的积累,产生光抑制,保持较高的LSP。以上研究揭示了沙地赤松对光具有很强的适应性,也揭示了沙地赤松生长量和生物量均较大的内在生理机制。
在P相时荧光水平达到了最大值(Fm),Fm是在1s内获得的,它代表了PSⅡ反应中心的关闭,代表了PSⅡ和质体醌A(QA)所有原初电子受体的完全还原,及PSⅡ受体侧电子流的饱和[30]。这说明沙地赤松并非所有的光合机构性能指标都好于樟子松,只是总体性能优于樟子松而已。
4 结论4.1 与樟子松相比,成林阶段沙地赤松具有较大的生长量、生物量,这与其较强的光合作用密切相关。沙地赤松Pmax和平均Pn均高于樟子松,且光适应范围较大,具有较高的LSP和较低的LCP,暗呼吸速率(Rd)较低,光合效率(Pmax/Rd)较高。
4.2 JIP-测定揭示了沙地赤松较强光合作用的内在机制,在O相至P相范围内,其相对可变荧光值(Vt)总体较低,说明其在电子传递过程中耗散能量较低,而用于光化学的能量较高。比活性参数进一步表明沙地赤松单位激发态面积反应中心数目、吸收的光量、被反应中心捕获的光量、用于电子传递的能量均较高,也因此具有较高的性能指数,其PI(ABS/CSo/CSm)分别是樟子松的1.42、1.65和1.63倍。
4.3 在沙地环境下,沙地赤松生长表现和光合性能优于樟子松,研究结果为扩大该树种在沙地上的引种栽培提供了光合理论依据。
| [1] | Krause G H, Weis F.Chlorophyll fluorescence and photosynthesis: the basics.Annual Review of Plant Physiology and Plant Molecular Biology,1991,42 313–349. |
| [2] | Demmig-Adams B, Adams W W III.The role of xanthophyll cycle carotenoids in the protection of photosynthesis.Trends in Plant Science,1996,1(1): 21–26. |
| [3] | 张谧, 王慧娟, 于长青.超旱生植物沙冬青高温胁迫下的快速叶绿素荧光动力学特征.生态环境学报,2009,18(6): 2272–2277. |
| [4] | Mereu S, Gerosa G, Marzuoli R, Fusaro L, Salvatori E, Finco A, Spano D, Manes F.Gas exchange and JIP-test parameters of two Mediterranean maquis species are affected by sea spray and ozone interaction.Environmental and Experimental Botany,2011,73 80–88. |
| [5] | Stirbet A, Govindjee.On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and photosystemⅡ: Basics and applications of the OJIP fluorescence transient.Journal of Photochemistry and Photobiology B: Biology,2011,104(1/2): 236–257. |
| [6] | Busch F, Hüner N P A, Ensminger I.Increased air temperature during simulated autumn conditions does not increase photosynthetic carbon gain but affects the dissipation of excess energy in seedlings of the evergreen conifer jack pine.Plant Physiology,2007,143(3): 1242–1251. |
| [7] | Martínez-Ferri E, Balaguer L, Valladares F, Chico J M, Manrique E.Energy dissipation in drought-avoiding and drought-tolerant tree species at midday during the Mediterranean summer.Tree Physiology,2000,20(2): 131–138. |
| [8] | Pollastrini M, Luchi N, Michelozzi M, Gerosa G, Marzuoli R, Bussotti F, Capretti P.Early physiological responses of Pinus pinea L.Tree Physiology,2015,35(3): 331–340. |
| [9] | Porcar-Castell A, Pfündel E, Korhonen J F J, Juurola E.A new monitoring PAM fluorometer (MONI-PAM) to study the short- and long-term acclimation of photosystem II in field conditions.Photosynthesis Research,2008,96(2): 173–179. |
| [10] | 宋晓东, 陈江燕, 刘桂荣, 徐贵军, 李淑华.樟子松枯梢病的侵染发生规律.森林病虫通讯,2000(4): 14–17. |
| [11] | 金永焕, 李敦求, 姜好相.不同土壤水分对赤松光合作用与水分利用效率的影响研究.中国生态农业学报,2007,15(1): 71–74. |
| [12] | Han Q M, Kawasaki T, Katahata S, Mukai Y, Chiba Y.Horizontal and vertical variations in photosynthetic capacity in a Pinus densiflora crown in relation to leaf nitrogen allocation and acclimation to irradiance.Tree Physiology,2003,23(12): 851–857. |
| [13] | 林思祖, 杨梅, 曹子林, 刘洪波, 陈艳.不同强度人为干扰对马尾松地上部分生物量及生产力的影响.西北植物学报,2004,24(3): 516–522. |
| [14] | Rauf S, Sadaqat H A.Effects of varied water regimes on root length, dry matter partitioning and endogenous plant growth regulators in sunflower (Helianthus annuus L.Journal of Plant Interactions,2007,2(1): 41–51. |
| [15] | Myers D A, Thomas R B, Delucia E H.Photosynthetic capacity of loblolly pine (Pinus taeda L.Plant, Cell and Environment,1999,22(5): 473–481. |
| [16] | Bassman J H, Zwier J C.Gas exchange characteristics of Populus trichocarpa, Populus deltoides and Populus trichocarpa × P.P. deltoides clones. Tree Physiolog,1991,8(2): 145–159. |
| [17] | Perveen S, Shahbaz M, Ashraf M.Regulation in gas exchange and quantum yield of photosystem Ⅱ (PSⅡ) in salt-stressed and non-stressed wheat plants raised from seed treated with triacontanol.Pakistan Journal of Botany,2010,42(5): 3073–3081. |
| [18] | Zeliou K, Manetas Y, Petropoulou Y.Transient winter leaf reddening in Cistus creticus characterizes weak (stress-sensitive) individuals, yet anthocyanins cannot alleviate the adverse effects on photosynthesis.Journal of Experimental Botany,2009,60(11): 3031–3042. |
| [19] | 陈福明, 陈顺伟.混合液法测定叶绿素含量的研究.林业科技通讯,1984(2): 4–8. |
| [20] | Maxwell K, Johnson G N.Chlorophyll fluorescence-a practical guide.Journal of Experimental Botany,2000,51(345): 659–668. |
| [21] | Hossain M D, Hanafi M M, Saleh G, Foroughi M, Behmaram R, Noori Z.Growth, photosynthesis and biomass allocation of different kenaf (Hibiscus cannabinus L.Australian Journal of Crop Science,2012,6(3): 480–487. |
| [22] | Gregory P J. Roots and the physico-chemical environment. In: Gregory P J, ed. Plant Roots: Growth, Activity, and Interaction with Soils. Oxford: Blackwell Publishing, 2006: 131-173. |
| [23] | Heimann M, Reichstein M.Terrestrial ecosystem carbon dynamics and climate feedbacks.Nature,2008,451(7176): 289–292. |
| [24] | Cayón S G.The evolution of photosynthesis, transpiration and chlorophyll during the development of leaves of plantain (Musa AAB Simmonds).Infomusa,2001,10(1): 12–15. |
| [25] | 李鹏民, 高辉远, StrasserR J.快速叶绿素荧光诱导动力学分析在光合作用研究中的应用.植物生理与分子生物学学报,2005,31(6): 559–566. |
| [26] | Luo Y Y, Zhao X Y, Zhang J H, Li Y L, Zuo X A, Sun D C.Photosynthesis of Digitaria ciliaris during repeated soil drought and rewatering.Sciences in Cold and Arid Regions,2015,7(1): 81–87. |
| [27] | Strasser R J, Tsimilli-Michael M, Srivastava A. Analysis of the chlorophyll a fluorescence transient. In: Papageorgiou G, Govindjee, eds. Advances in Photosynthesis and Respiration. Netherlands: Springer, 2004: 321-362. |
| [28] | van Heerden P D R, Strasser R J, Krüger G H J.Reduction of dark chilling stress in N2-fixing soybean by nitrate as indicated by chlorophyll a fluorescence kinetics.Physiologia Plantarum,2004,121(2): 239–249. |
| [29] | Schansker G, Tóth S Z, Strasser R J.Methylviologen and dibromothymoquinone treatments of pea leaves reveal the role of photosystem I in the Chl a fluorescence rise OJIP.Biochimica et Biophysica Acta (BBA) - Bioenergetics,2005,1706(3): 250–261. |
| [30] | Govindjee.Sixty-three years since Kautsky: Chlorophyll a fluorescence.Australian Journal of Plant Physiology,1995,22(2): 131–160. |