 2016, Vol. 36
2016, Vol. 36文章信息
- 张瑞玲, 王万雄, 秦丽娟
- ZHANG Ruiling, WANG Wanxiong, QIN Lijuan
- 具有Allee效应的合作进化
- Cooperative evolution of a population with the Allee effect
- 生态学报, 2016, 36(1): 261-267
- Acta Ecologica Sinica, 2016, 36(1): 261-267
- http://dx.doi.org/10.5846/stxb201407261515
-
文章历史
- 收稿日期: 2014-07-26
- 修订日期: 2015-07-27






2. 甘肃农业大学, 理学院, 兰州 730070
2. College of Science, Gansu Agricultural University, Lanzhou 730070, China
达尔文生物进化论的核心是自然选择学说——物竞天择、弱肉强食。物种在生存竞争的过程中,经过自然选择的作用,逐渐产生更能适存的物种,实现生物的进化。大自然中存在着竞争与矛盾冲突,合作行为是生物体有利于其它个体的各种行为策略,是社会性生物的一个重要特征。合作进化机理是长期困扰进化生物学的一个难题,问题的关键是合作如何通过自然选择从背叛种群中进化出来,及合作个体怎么才能避免背叛个体的剥削,从而抵制它们的入侵!囚徒困境的博弈不仅是博弈论中最具代表性的例子,而且作为研究合作进化的一个普遍范式[1]。
此外,Allee[2]指出,群聚有利于种群的增长和存活,每个生物都有自己的最适密度,其过分稀疏和拥挤都会对生殖发生负作用,阻止生长。Allee效应的产生源于很多方面的不利因素[3],例如,配偶,生殖能力,过度掠夺,环境条件,同系繁殖等等。对于一个具有Allee效应的种群来说,当种群密度低于某一阈值时,物种将会灭绝,也就是说该种群具有一个足以维持生存的最低密度。尤其那些濒危的哺乳类种群,其种群稀疏,更容易受到Allee效应的影响。许多理论研究表明Allee效应在种群动力学研究中存在强大的潜在影响,这引起了生态学家的广泛关注[4, 5]。关于Allee效应对种群生态学系统稳定性的影响的研究也越来越受到人们的重视[6, 7, 8, 9]。因此,考虑到Allee效应和合作行为的进化分别是种群生态学和进化生物学的重要理论问题。Feng Zhang[10]等人做了囚徒困境博弈中的生态进化反馈和入侵的研究,结果表明:合作行为在密度依赖选择的条件下更容易进化,该工作没有涉及种群受Allee效应影响下的合作行为的进化,为了使模型更加切合实际。本文提出了受Allee效应影响的囚徒困境博弈模型,并利用动力模型分析及数值模拟方法对新建模型的基本性质进行初步研究,说明Allee效应对合作进化的影响,丰富了合作行为的演化动力学理论。
1 模型建立 1.1 生态囚徒困境博弈模型生物界中,不论在同一种生物内部还是不同的物种之间,从低等的微生物到高等的多细胞生物都有合作行为的存在。尽管合作可以给群体带来利益,但合作者遇到背叛者不可避免地付出代价。因此,合作行为进化的关键问题存在两方面:一方面,如何从背叛者组成的种群中通过自然选择进化产生合作行为?另一方面,合作种群如何有效地抵制背叛个体入侵?
囚徒困境博弈是合作行为进化的隐喻,假设种群由合作(C)和背叛(D)的个体组成,合作个体自身付出一定的代价c,对和它相互作用的对方产生利益b(b>c>0),然而,背叛个体即不付出代价也不会产生利益。这两种行为的囚徒困境博弈的赢得矩阵为:
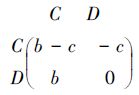
定义合作个体与背叛个体分类相遇的概率[11]:
(1)一个个体遇到采取同样策略个体的概率是m;
(2)一个个体随机遇到其他个体(包括采取同样策略的个体)的概率是1-m。
Feng Zhang,Cang Hui[10]研究了合作个体在囚徒困境博弈中的生态进化反馈及入侵,动力系统模型由如下的微分方程表达:

对系统(1)通过理论分析与数值模拟,结果表明:个体间的分类是至关重要的,同时高死亡率和低的种群占有率更能促进合作进化。
1.2 种群具有Allee效应的合作进化模型林振山等[12]以丹顶鹤为例研究了人类活动影响下具有Allee效应的非自治种群演化模式,其Allee效应形式为$\frac{N}{N+\alpha }$ ,式中N为种群密度,α为Allee效应常数,α越大意味着Allee效应越强。考虑到系统(1)中的种群若具有Allee效应,则其种群的总体占有率为x+y,且种群中的个体不论采取哪种行为策略都会受到Allee效应的影响。将Allee效应代入模型(1),建立具有Allee效应的动力系统模型如下:
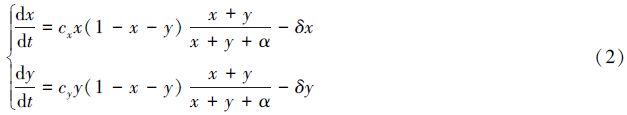
易验证,在分类相遇m>0且密度依赖选择(x+y<1)的情形下,有利于合作行为的进化。进一步通过计算可得系统(2)的平衡点如下:
(1)E(0,0),合作行为与背叛行为都灭绝;
(2)最多存在四个边界平衡点:(ⅰ)当δ<μ且(δ-μ)2>4μδα时,存在四个边界平衡点(其中,两个平衡点是背叛行为独存,两个平衡点是合作行为独存),(ⅱ)当μ+m(b-c)<min{δ,(1-m)(b-c)}时,存在两个边界平衡点(合作行为独存);
(3)最多存在一个内部平衡点(x*,y*),其中:
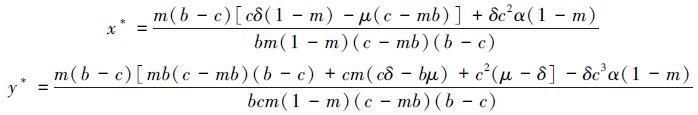
且满足条件:

代表合作行为与背叛行为共存。如果mb>c,系统不存在内部平衡点。
平衡点(x*,y*)的雅克比行列式为:
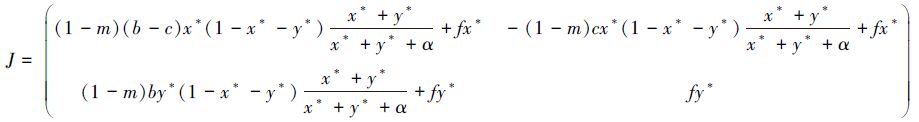
其中,$f=[\mu +(1-m){{x}^{*}}b]\frac{\alpha -2\alpha ({{x}^{*}}+{{y}^{*}})-{{({{x}^{*}}+{{y}^{*}})}^{2}}}{{{({{x}^{*}}+{{y}^{*}}+\alpha )}^{2}}}$ 。因此,可知其对应的特征方程为:

其中,

由于det>0,则当tr2-4det>0且tr>0,平衡点(x*,y*)为不稳定的结点;当tr2-4det>0且tr<0,平衡点(x*,y*)为稳定的结点;当tr2-4det<0且tr>0,平衡点(x*,y*)为不稳定的焦点;当tr2-4det<0且tr<0,平衡点(x*,y*)为稳定的焦点。由此易知,系统(2)最多存在一个内部平衡点,随着参数的变化,内部平衡点会从一个结点演化成一个焦点,即若tr2-4det=0,系统(2)出现分叉现象。与此同时,内部平衡点由稳定状态向不稳定状态演变,即若tr=0,系统(2)存在极限环。与系统(1)相比,Allee效应对合作进化是一个潜在的干扰因素。由于参数的繁杂,很难推断出Allee效应具体如何影响合作进化及系统(1)的稳定性。因此,利用Matlab软件对系统(2)进行数值模拟。
2 数值模拟与结果分析为了定性讨论Allee效应对合作进化的影响,取文献[10]中的参数μ=0.1,m=0.1,b=1,从系统(2)的演化结果可知,在Allee效应α=0.05的影响下,系统(1)中由于合作行为的高死亡率与高代价(c=0.3,δ=0.09)产生的阻尼振荡(即焦点)变成周期性振荡(即极限环,图 1-ⅰ);更高的死亡率(c=0.25,δ=0.15)产生的阻尼振荡将会消失,种群灭绝(图 1-ⅲ);而合作行为的更高代价(c=0.39,δ=0.137)产生的周期性震荡也会消失,种群灭绝;在较低的死亡率和代价(c=0.25,δ=0.15)条件下,Allee效应对系统的影响不大,系统(2)出现一个临界值,当初值大于这个临界值时,系统仍然存在一个稳定的内部平衡点(图 1-ⅱ)。由此可知,在恶劣的环境条件下,若种群由于寻找配偶的困难,社会功能异常,近交衰退等原因受Allee效应的影响时,更容易导致物种灭绝,影响生物的多样性。

|
| 图1 受Allee效应α=0.05影响下系统(2)的演化相图 Fig.1 The evolutive phase diagram of system (2) with Allee effect α=0.05 参数取值:μ=0.1,m=0.1,b=1, (ⅰ) c=0.3,δ=0.09;(ⅱ)c=0.15,δ=0.15; (ⅲ)c=0.25,δ=0.15; (iv)c=0.39,δ=0.137 |
由图(2-ⅰ)可见,取参数μ=0.1,m=0.1,b=1,c=0.15,δ=0.15,不受Allee效应影响时,系统存在一个稳定的结点;然而,在Allee效应常数α=0.05时,若合作种群与背叛种群的初始占有率大于某个临界值(图 1-ⅱ),Allee效应更容易使系统达到稳定状态,促进合作进化(图 2-ⅱ);继续增强Allee效应(α>0.077),就会导致种群灭绝,不利于合作进化(图 2-ⅲ)。说明在环境较为优越的条件下,适当的Allee效应促进合作进化,而继续增强Allee效应将会导致物种灭绝。

|
| 图2 受Allee效应影响的系统(2)的演化曲线 Fig.2 The evolvement curves of system (2) with Allee effect 参数取值:μ=0.1,m=0.1,b=1,c=0.15,δ=0.15,Allee效应: (ⅰ)α=0;(ⅱ)α=0.05;(ⅲ)α=0.15 |
空间网格上的博弈行为是近年进化博弈研究的新焦点[13],基本思想是把种群安置在规则的空间网格上,一部分斑块只有一个采取特定策略的个体占有,所有个体只和它局部相邻斑块上的个体相互作用,根据它们的总赢得和邻体中个体的总赢得决定下一次所采取的策略。取参数μ=0.1,m=0.1,b=1, c=0.15,δ=0.09,对未受Allee效应及受不同Allee效应强度影响的空间囚徒困境博弈,在200×200的空间网格上,只考虑相邻于每个斑块边的4个斑块(冯诺伊曼邻体),进行时间t=200的空间数值模拟(图 3)。由图 3可见,当系统未受Allee效应影响时,除了少数的空斑块,斑块全部被合作策略及背叛策略的个体占有。当种群受Allee效应(α=0.05)影响时,空间中空斑块的数量明显增多,合作个体的占有比率有所增加。然而,继续增加Allee效应(α=0.15),空间的绝大部分斑块未被占有,成为空斑块,只有极少数的被种群占有;再继续增加Allee效应,将导致物种灭绝。

|
| 图3 在200×200网格上,受Allee效应影响的空间囚徒困境在时间t=200的空间格局 Fig.3 At time t=200,spatial patterns of spatial Prisoner′S Dilemma with Allee effect on a 4016401 lattice |
下面将详尽地讨论Allee效应在未使物种灭绝的情况下,如何促进合作行为的进化,如图 4所见,当时间t<80时,Allee效应抑制合作进化;当时间t>80时,Allee效应促进合作进化,且随着Allee效应强度的增强,合作个体的斑块占有率逐渐增多。然而,对于背叛个体,Allee效应强度越强,其斑块则占有率越低。若Allee效应强度大于0.335时,物种灭绝。

|
| 图4 合作个体、背叛个体对Allee效应变化(α=0,0.04,0.08,…,0.36)的相应响应 Fig.4 The response of cooperators and defector under the change of Allee effec(α=0,0.04,0.08,…,0.36) 参数取值:μ=0.1,m=0.1,b=1,c=0.15,δ=0.09 |
以上的数值模拟和预测说明,Allee效应对合作进化的影响显著,对种群的续存是不可忽视的干扰因素[14]。与没有受Allee效应的系统(1)相比,Allee效应破坏原来的平衡,形成新的平衡。合作个体在高代价、高死亡的条件下,Allee效应使以阻尼振荡的相互作用共存的两种行为策略变成周期性振荡的共存模式;在更高死亡率和代价的环境中,Allee效应使博弈共存的物种灭绝。但是,在优越的环境条件下(即死亡率较低),Allee效应反而促进合作进化,且Allee效应的强度越强越有利于合作行为的进化,不过种群的总体斑块占有率会越低。与此同时,Allee效应强度过强,就会导致物种灭绝,行为策略不复存在。也就是说强Allee效应不仅减少物种的多样性,而且影响生物进化。
3 结论Allee效应是种群动力学中的一个重要的调节机制:低的种群密度使雌雄个体相遇机会减少,寻觅配偶的困难大大增加[15](例如在防治橘小实蝇虫害的过程中,就采用了释放不育雄虫制造Allee效应的措施),对于植物来说同样也会由于低密度使授粉成功的机会大大减少。非洲野狗是一种典型的群居动物,它们具有群体合作哺育后代、合作觅食的习性,过小的种群规模致使某些合作技能丧失,不能维持种群个体的生存。一定的种群规模也有利于抵御敌害,增加群体的反捕食的能力。很多自然界的物种曾被报导过具有Allee效应,比如植物[16],海洋无脊椎动物[17],昆虫[18],哺乳动物[19]等。正是因为Allee效应在现实生物种群中是普遍存在的,加之合作进化并不是一个进化稳定策略,在一次囚徒困境博弈中,不论对方采取什么策略,背叛总是最好的策略,所以背叛行为是Nash平衡,也是进化稳定的策略。这意味着合作行为成功地入侵一个背叛的种群几乎是不可能的,即使是已经成为合作的种群,也很难抵制背叛个体的入侵。于是,合作进化对Allee效应的影响更加敏感,将Allee效应这一因素引入囚徒困境博弈是合理的,而且更具有现实意义。
通过平衡态分析与数值模拟发现,在Allee效应的影响下,分类相遇对合作进化仍然是必要条件。Allee效应导致合作行为与背叛行为构成的囚徒困境系统具有更多的平衡态,破坏原来的平衡,形成新的平衡态,并且最终平衡态值随行为策略的初始斑块占有率的变化而改变,这对合作进化的管理与保护具有重要的启示意义。系统(1)研究结果表明,优越的外部环境有利于种群的发展,但却抑制了合作行为的进化,相反,适当的环境压力可以促进合作行为的空间入侵和维持。考虑了种群受Allee效应的影响之后,分析结果说明,在恶劣的环境条件下,Allee效应将导致物种灭绝,这也就意味着合作进化不复存在,而在相对优越的外部环境中,Allee效应的存在促进合作行为的进化,且随着Allee效应强度的增强,更有利于合作行为的入侵,不过过强的Allee效应依旧会导致物种灭绝。因此,从保护生物学的角度来说,Allee效应对合作进化的影响是不容忽视的,尤其是濒临物种。应该采取相应的保护措施,例如,就地保护或迁地保护[20]等,这不仅能保护生物的多样性,而且还可以有效地促进合作行为的进化。
| [1] | Li Y, Lan X, Deng X Y, Sadiq R, Deng Y. Comprehensive consideration of strategy updating promotes cooperation in the prisoner's dilemma game. Physica A: Statistical Mechanics and its Applications, 2014, 403: 284-292. |
| [2] | Allee W C. Animal Aggregations: A study in General Sociology. Chicago: University of Chicago Press, 1931. |
| [3] | Courchamp F, Berec L, Gascoigne J. Allee Effects in Ecology and Conservation. Chicago: University of Chicago Press, 2008. |
| [4] | Shi J P, Shivaji R. Persistence in reaction diffusion models with weak allee effect. Journal of Mathematical Biology, 2006, 52(6): 807-829. |
| [5] | Berec L, Angulo E, Courchamp F. Multiple allee effects and population management. Trends in Ecology & Evolution, 2007, 22(4): 185-191. |
| [6] | Pal P J, Saha T, Sen M, Banerjee M. A delayed predator-prey model with strong Allee effect in prey population growth. Nonlinear Dynamics, 2012, 68(1/2): 23-42. |
| [7] | Bai Z J, Yang W S, Li X P. Qualitative Analysis of a predator-prey system with Allee effect for prey. Annals of Differential Equations, 2007, 23(4): 386-390. |
| [8] | Aguirrea P, González-Olivaresb E, Sáeza E. Two limit cycles in a Leslie-Gower predator-prey model with additive Allee effect. Nonlinear Analysis: Real World Applications, 2009, 10(3): 1401-1406. |
| [9] | Wang W X, Zhang Y B, Liu C Z. Analysis of a discrete-time predator-prey system with Allee effect. Ecological Complexity, 2011, 8(1): 81-85. |
| [10] | Zhang F, Hui C. Eco-evolutionary feedback and the invasion of cooperation in prisoner's dilemma games. PLoS One, 2011, 6(11): e27523. |
| [11] | Eshel I, Cavalli-Sforza LL. Assortment of encounters and evolution of cooperativeness. Proceedings of the National Academy of Sciences of the United States of America, 1982, 79(4): 1331-1335. |
| [12] | 林振山, 刘会玉, 刘红玉. 人类活动影响下具有Allee 效应的非自治种群演化模式的研制及其应用--以丹顶鹤为例. 生态学报, 2005, 25(5): 945-951. |
| [13] | Hauert C, Szabo G. Game theory and physics. American Journal of Physics, 2005, 73(5): 405-414. |
| [14] | 陈玲玲, 林振山. 非Allee与Allee竞争种群系统的动力学研究. 南京师范大学学报: 自然科学版, 2008, 31(2): 124-129. |
| [15] | González-Olivares E, Mena-Lorca J, Rojas-Palma A, Flores J D. Dynamical complexities in the Leslie-Gower predator-prey model as consequences of the Allee effect on prey. Applied Mathematical Modelling, 2011, 35(1): 366-381. |
| [16] | Ferdy J B, Austerlitz F, Moret J, Godelle P H, Godelle B. Pollinator-induced density dependence in deceptive species. Oikos, 1999, 87(3): 549-560. |
| [17] | Stoner A, Ray-Culp M. Evidence for Allee effects in an over-harvested marine gastropod: denity dependent mating and egg production. Marine Ecology Progress Series, 2000, 202: 297-302. |
| [18] | Kuussaari M, Saccheri I, Camara M, Hanski I. Allee effect and population dynamics in the Glanville fritillary butterfly. Oikos, 1998, 82(2): 384-392. |
| [19] | Courchamp F, Grenfell B, Clutton-Brock T. Impact of natural enemies on obligately cooperative breeders. Oikos, 2000, 91(2): 311-322. |
| [20] | 安德烈·普林. 保护生物学. 贾竞波, 译. 北京: 高等教育出版社, 2005. |