 2015, Vol. 35
2015, Vol. 35文章信息
- 顾婷婷, 许敏, 赵以军, 程凯
- GU Tingting, XU Min, ZHAO Yijun, CHENG Kai
- 温度与CO2浓度升高对集胞藻PCC 6803修复UV损伤的关键基因转录量的影响
- Effects of increased CO2 and elevated temperature on transcript levels of key ultraviolet damage repair genes in Synechocystis sp. PCC 6803
- 生态学报, 2015, 35(9): 3132-3137
- Acta Ecologica Sinica, 2015, 35(9): 3132-3137
- http://dx.doi.org/10.5846/stxb201306101617
-
文章历史
- 收稿日期:2013-06-10
- 修订日期:2014-05-22






2. 湖北工业大学, 资源与环境工程学院, 河湖生态修复与藻类利用湖北省重点实验室, 武汉 430068
2. College of Resources and Environmental Engineering, Key Laboratory of Ecological Remediation of Lakes and Rivers and Algal Utilization of Hubei Province, Hubei University of Technology, Wuhan 430068, China
日光中的UVB(ultraviolet-B)辐射对水生生物具有明显的伤害作用,特别是由于形成了环丁烷嘧啶二聚体(cyclobutane-pyrimidine dimers,CPD)和6-4光产物(6-4 photoproducts,6-4PPs)[1],从而阻碍DNA复制与转录,影响蛋白质的功能,造成浮游植物(包括蓝藻)、大型藻类和其它水生植物的光合作用降低,甚至会对初级生产力产生严重影响[1]。与此同时,为了抵消UVB对生物体的负面影响,很多水生生物都进化出了相应的保护、回避或修复机制,这些机制对于水生生物种群的维持和生态功能的发挥具有重要意义[1]。
集胞藻PCC 6803(Synechocystis sp. PCC 6803)是蓝藻光合作用研究的模式生物,其对UV损伤的修复机制主要体现在以下两个层面:在DNA水平上,phrA基因编码的光聚酶(photolyase)能够直接修复损伤的DNA[2];在蛋白质水平上,D1蛋白作为光系统Ⅱ(PSⅡ)光反应中心的重要部件,同时也是UV损伤的关键位点,而在编码集胞藻PCC 6803 D1蛋白的psbA基因家族中,psbA2和psbA3是PSⅡUV损伤修复的关键基因[3],psbA1在正常条件下则未检测到表达[4]。
大气中的CO2是蓝藻光合作用的基本原料,其浓度变化对蓝藻的生长、生理及生态功能有着巨大影响[5]。至21世纪初,大气中的CO2浓度比工业化前增加了约40%,并将在本世纪末达到550—900μmol/mol[6],届时,全球气温将比工业化前升高1.6—7.0℃[7]。大气CO2浓度与温度同步升高将对蓝藻的生长、生理及生态功能产生更为复杂的影响[8],但目前关于这方面的研究主要集中于蓝藻的生长、细胞成份等方面 [8-10],而缺乏有关大气CO2浓度与温度升高对蓝藻UV修复的联合作用的研究。本文以集胞藻PCC 6803为研究对象,采用RT-qPCR技术探索了CO2浓度升高以及温度升高对其UV损伤修复基因转录量的影响,有助于推测全球气候变化对蓝藻的UV损伤修复能力的影响,具有重要的生态学意义。
1 材料与方法 1.1 藻种及其传代周期集胞藻PCC 6803来源于中国科学院水生生物研究所,培养基为BG-11[11],光照强度为1200 lx,采用静置培养(每天手工摇动1—2次),光暗周期为12 ∶ 12。整个实验过程中每隔2周传代1次,传代后的初始藻细胞密度为5×106个/mL,培养1周后达到对数期。
1.2 培养条件本实验使用的自动控制培养箱可在箱体内控制CO2浓度(并非向培养基中充气)和温度[12]。本研究涉及5种处理:(1)25℃+ CO2浓度400μmol/mol(CK组),(2)29℃+ CO2浓度400μmol/mol(温度升高组),(3)25℃+ CO2浓度800μmol/mol(CO2升高组1),(4)29℃+ CO2浓度800μmol/mol(温室效应组),(5)25℃+ CO2浓度1200μmol/mol(CO2升高组2)。每组均设3平行。
1.3 转录本拷贝数的测定 1.3.1 样品的采集于第7次传代后的第8天(此时集胞藻PCC 6803在上述各培养条件下培养的时长总计为15周)取对数期藻液用于RT-qPCR检测,同时进行藻细胞计数。
1.3.2 RNA的提取取50—100 mg冷冻的液氮研磨至粉末状,置于1.5 mL离心管中,加入1mL Trizol充分匀浆,室温静置5min;加入0.2mL氯仿,剧烈振荡15s,静置3min;4℃,12000r/min离心10min,取上清;加入0.5mL异丙醇,混匀,冰上静置20—30min;4℃,12000r/min离心10min,弃上清;加入1mL 75%乙醇洗涤沉淀后于室温放置晾干,再加入适量的无RNA酶的双蒸水溶解[13]。
1.3.3 cDNA的合成使用的是BBI(波士顿生物技术公司)第一链cDNA合成试剂盒将获得的总RNA反转录合成cDNA,于-20℃保存待用。
1.3.4 qPCR本研究采用Taqman探针法进行绝对定量qPCR,目的基因中psbA基因家族中的3个基因的引物均来源于文献报道[13],16S rRNA和phrA基因的引物是通过primer5.0软件设计(表 1)。
| 基因名称 Gene name | 引物方向与探针 Orientation and probe | 引物序列(5′-3′) Primer sequence(5′-3′) |
| qPCR的总体系为25μL:12.5μL HS qPCR Master Mix(2X),0.5μL引物F(20μmol/L),0.5μL引物R(20μmol/L),0.5μL探针P(20μmol/L),9μLddH2O,2μLcDNA模板;PCR循环的条件为:95℃ 2 min变性后进行40个循环(95℃ 10 s,60℃ 1min) | ||
| 16S rRNA | 上游 | AAAGCGTCCGTAGGTGGTTAT |
| 下游 | GTCCCTCAGTGTCAGTTTCAGC | |
| 探针 | CCCAGTGTAGCGGTGAAATGCGTA | |
| phrA | 上游 | AAAGCGAGATTTAGCAGTGGC |
| 下游 | CCAAAAGGGAGTGTAAACCGT | |
| 探针 | ATCGCCACGGAATGGGATCAACTCA | |
| psbA1 | 上游 | AGCTTAAACCCAAAATCTTACTTCGT |
| 下游 | CCATCAGAGAAGGAGCCTTGACCA | |
| 探针 | CAAAGCCTGTGGTCACGGTTCTGTT | |
| psbA2 | 上游 | CCTTTAGACTAAGTTTAGTTCCA |
| 下游 | CCATCAGAGAAGGAGCCTTGACCA | |
| 探针 | CCCGTTGCTGGTTCTTTGCTTTAC | |
| psbA3 | 上游 | GAGCTTGAGGCCAAATCCTTTGAACA |
| 下游 | CCATCAGAGAAGGAGCCTTGACCA | |
| 探针 | TTCCTTGCTCTACGGTAACAACATCATC | |
将样品中目的基因的总拷贝数(X0)除以对应的藻细胞密度,得到单个细胞中各目的基因的转录本的拷贝数(A),其中X0的计算方法如下[14]:
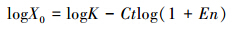
式中,K为产物荧光信号达到设定阈值时扩增产物的量;En为扩增效率(通过质粒样品的标准曲线得到);X0为起始模板量;Ct为产物荧光信号达到设定阈值时的循环次数。
1.5 转录本拷贝数变幅的计算、作图与统计分析各试验组单个细胞中转录本拷贝数相对于CK组的变幅(V)的计算方法为:
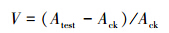
式中,Atest为各试验组中单个细胞中目的基因的转录本拷贝数;Ack为CK组中单个细胞中目的基因的转录本拷贝数。
作图使用的是GraphPad Prism 5(误差线均用SD表示,n=3),统计分析采用SPSS20.0的ANOVA分析方法(其多重比较采用LSD法)。
2 结果和分析 2.1 16S rRNA转录本拷贝数的变化表 2是单个细胞中各目的基因的转录本拷贝数。
| 实验组Groups | 16S rRNA | phrA | psbA1 | psbA2 | psbA3 |
| phrA:光聚酶编码基因;psbA1、psbA2、psbA3:D1蛋白编码基因 | |||||
| CK | 365.45±73.84 | 0.25±0.05 | 0.22±0.06 | 20.89±0.70 | 21.67±4.12 |
| 温度升高组Elevated temperature | 68.67±2.12 | 0.25±0.03 | 0.14±0.03 | 2.13±0.17 | 3.78±0.69 |
| CO2升高组1 Elevated CO2 1 | 56.17±5.13 | 0.02±0.00 | 0.03±0.02 | 0.85±0.09 | 1.47±0.53 |
| 温室效应组 Greenhouse effect | 158.22±19.52 | 0.13±0.02 | 0.10±0.03 | 3.29±0.51 | 5.72±1.42 |
| CO2升高组2 Elevated CO2 2 | 79.65±11.27 | 0.06±0.00 | 0.05±0.02 | 2.55±0.44 | 3.93±0.69 |
根据表 2,得到各试验组16S rRNA转录本拷贝数相对于CK组的变幅如图 1所示:各试验组的16S rRNA转录本拷贝数都发生了不同程度的下调,其中下调幅度最大的为CO2升高组1(约85%),而下调幅度最小的为温室效应组(约60%)。

|
| 图 1 不同条件下单个细胞中16S rRNA转录量的变幅 Fig.1 The transcripts variation of 16S rRNA of each cell in different conditions |
根据表 2,得到各试验组phrA/psbA1/psbA2/psbA3基因转录本拷贝数相对于CK组的变幅如图 2所示。

|
| 图 2 不同条件下单个细胞中phrA/psbA1/psbA2/psbA3基因的转录量 Fig.2 The transcripts variation of phrA/psbA1/psbA/psbA3 of each cell in different conditions phrA:光聚酶编码基因;psbA1、psbA2、psbA3:D1蛋白编码基因 |
由图 2可见,除了温度升高组的phrA基因外,其它情况下各UV修复基因的转录本拷贝数都发生了明显下调,平均下调幅度约50%,且表现出了以下特点:1)各试验组中4种基因的转录本拷贝数的下调幅度差异显著(P<0.05),且均以psbA2基因转录量的下调幅度最大,psbA3基因的下调幅度仅略小于psbA2基因;2)温度升高与CO2浓度升高对psbA3/psbA2基因转录本拷贝数下调的联合作用结果表现为互相抵消,如:温度升高组中psbA3基因的降幅为82%左右,CO2浓度升高组1中psbA3基因的降幅为93%左右,但在温室效应组中psbA3基因的降幅却仅为73%左右,16S rRNA也表现出了类似的现象(图 1)。3)通过对比CO2升高组1与CO2升高组2的结果,发现CO2浓度的持续升高不但不能进一步增加各基因转录本数的下调幅度,而且会导致各基因转录本拷贝数的下调幅度的明显减少,16S rRNA也表现出了类似的现象(图 1)。
3 讨论本研究发现CO2浓度升高时,集胞藻PCC 6803的16S rRNA基因与关键UV修复基因转录本的拷贝数均明显减少。Baosheng Qiu等人的研究显示CO2浓度加倍虽然会导致铜绿微囊藻生物量的增加52%—77%,但是多糖的合成却会减少[10]。此外,也有研究表明加倍的CO2浓度和同步的温度升高将虽然会使海洋聚球藻的藻胆素含量、叶绿素a含量和光合速率大幅提高[8],但这些效应在原绿球藻中却不存在[8],说明不同的微藻对CO2浓度以及温度的响应机制是存在较大差异的。
正常条件下,集胞藻PCC 6803中psbA2的转录本拷贝数占psbA转录本总拷贝数的90%[4]。有证据表明,作为PSⅡUV损伤修复关键基因的psbA2/psbA3基因,其转录本拷贝数的下调会导致集胞藻PCC 6803UV损伤修复能力的下降[4]。集胞藻PCC 6803中phrA基因在正常情况下的表达量虽很低,但是其表达产物对UV辐射造成的DNA损伤的修复却起着至关重要的作用[15-16],而且蓝藻对PSⅡ的UV损伤的修复能力也会受到光聚酶的修复能力的影响[17]。因此,本研究所观察到的psbA2/psbA3/phrA基因转录本拷贝数大幅下调的现象预示着温室效应将有可能导致蓝藻的UV损伤修复能力的下降,这意味着表层水中蓝藻因UV损伤而死亡或生长变慢的可能性会增加,而臭氧层破坏所导致的全球地表紫外线的增强将使这一可能性进一步提高[1],但是考虑到CO2和温度的升高同时也会明显的刺激藻类的生长[8],所以其综合效应(特别是考虑到不同微藻对CO2、水温的响应幅度还存在差异[8])仍难以准确判断。
与psbA2/psbA3基因不同,psbA1基因由于表达量极低,其功能通常被忽视[4],目前发现该基因仅在微氧条件[13]下有微量表达,其余基本上不受环境变化的影响,而本研究发现,各实验组中psbA1基因转录本的拷贝数均明显下调,其中温室效应组的下调幅度超过50%,说明psbA1基因的转录量有可能随着全球气候变化而改变。
核糖体是生物体发挥正常生理功能的基础,16S rRNA基因也是RT-qPCR技术中的常用内参基因[18],但是,本研究却发现集胞藻PCC 6803中该基因的表达量在温室效应条件下会明显下降,这可能与细胞生长状态的改变有关[19]。上述结果一方面说明温室效应将可能影响蓝藻的核糖体合成,另一方面也说明在研究蓝藻的基因转录量时,应慎重采用16S rRNA基因作为RT-qPCR的内参基因[20]。
本研究表明,温度升高与CO2浓度升高对16S rRNA及psbA2/psbA3基因转录本拷贝数的联合作用表现为互相抵消,说明温度升高与CO2浓度升高的联合作用的机制比较复杂[8],需要通过试验进行研究而不能简单相加。
综上所述:温度与CO2浓度的升高可以导致集胞藻PCC 6803的psbA2/psbA3基因和16S rRNA转录本拷贝数的大幅减少,说明温室效应将有可能导致蓝藻的UV损伤修复能力与核糖体合成能力的下降;温度升高和CO2浓度升高对psbA2/psbA3基因和16S rRNA转录本拷贝数的联合作用表现为互相抵消,说明温度升高与CO2浓度升高的联合作用的机制较复杂,值得深入研究。
| [1] | Hader D P, Helbling E W, Williamson C E, Worrest R C. Effects of UV radiation on aquatic ecosystems and interactions with climate change. Photochemical and Photobiological Sciences, 2011, 10(2): 242-260. |
| [2] | Eker A P, Kooiman P, Hessels J K, Yasui A. DNA photoreactivating enzyme from the cyanobacterium Anacystis nidulans. Journal of Biological Chemistry, 1990, 265(14): 8009-8015. |
| [3] | Renu M, Vashudha P J, Swapan K, Bhattacharjee. psbA gene family coding D1 proteins of photosystem Ⅱ of cyanobacteria Synechocystis PCC 6803 is essential for DNA repair/protection. Current Science, 2011, 100(1): 58-68. |
| [4] | Mulo P, Sicora C, Aro E M. Cyanobacterial psbA gene family: optimization of oxygenic photosynthesis. Cellular and Molecular Life Sciences, 2009, 66(23): 3697-3710. |
| [5] | Behrenfeld M J, Robert T O, David A S, Charles R M, Jorge L S, Gene C F, Allen J M, Paul G F, Ricardo M L, Emmanuel S B. Climate-driven trends in contemporary ocean productivity. Nature, 2006, 444(7120): 752-755. |
| [6] | Lindroth R L. Impacts of elevated atmospheric CO2 and O3 on forests: phytochemistry, trophic interactions, and ecosystem dynamics. Journal of Chemical Ecology, 2010, 36(1): 2-21. |
| [7] | Intergovernmental Panel on Climate Change. Climate Change: Synthesis Report. Contribution of Working Groups I, Ⅱ and III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Geneva: IPCC, 2007. |
| [8] | Fu F X, Warner M E, Zhang Y H, Feng Y Y, Hutchins D A. Effects of increased temperature and CO2 on photosynthesis, growth, and elemental ratios in marine Synechococcus and Prochlorococcus (Cyanobacteria). Journal of Phycology, 2007, 43(3): 485-496. |
| [9] | Fu F X, Mulholland M R, Garcia N S, Beck A, Bernhardt P W, Warner M E, Sa udo-Wilhelmy S A, Hutchins D A. Interactions between changing pCO2, N2 fixation, and Fe limitation in the marine unicellular cyanobacterium Crocosphaera. Limnology and Oceanography, 2008, 53(6): 2472-2484. |
| [10] | Qiu B S, Gao K S. Effects of CO2 enrichment on the bloom-forming cyanobacterium Microcystis aeruginosa(cyanophyceae): physiological responses and relationships with the availability of dissolved inorganic carbon. Journal of Phycology, 2002, 38(4): 721-729. |
| [11] | Stanier R Y, Kunisawa M M, Cohen B G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriological Reviews, 1971, 35(2): 171-205. |
| [12] | 牛晓莹, 程凯, 荣茜茜, 许敏, 赵以军, 赵进. CO2浓度和温度升高对噬藻体PP 增殖的联合作用. 生态学报, 2012, 32(22): 6917-6924. |
| [13] | Sicora C I, Ho F M, Salminen T, Styringc S, Aroa E M. Transcription of a "silent" cyanobacterial psbA gene is induced by microaerobic conditions. Biochimicaet Biophysica Acta, 2009, 1787(2): 105-112. |
| [14] | 唐永凯, 贾永义. 荧光定量PCR 数据处理方法的探讨. 生物技术, 2008, 18(3): 47-56. |
| [15] | Ng W O, Pakrasi H R. DNA photolyase homologs are the major UV resistance factors in Synechocystis sp. PCC 6803. Molecular and General Genetics, 2001, 264(6): 924-970. |
| [16] | Ng W O, Zentella R, Wang Y, Taylor J S, Pakrasi H B. phrA, the major photoreactivating factor in the cyanobacterium Synechocystis sp. strain PCC 6803 codes for a cyclobutane-pyrimidine-dimer-specific DNA photolyase. Archives of Microbiology, 2000, 173(5/6): 412-417. |
| [17] | Vass I Z, Kos P B, Sass L, Nagy C I, Vass I. The ability of cyanobacterial cells to restore U-VB radiation induced damage to photosystem II is influenced by photolyase dependent DNA repair. Photochemistry and Photobiology, 2013, 89(2): 384-390. |
| [18] | Devers M, Soulas G, Martin-Laurent F. Real-time reverse transcription PCR analysis of expression of atrazine catabolism genes in two bacterial strains isolated from soil. Journal of Microbiological Methods, 2004, 56(1): 3-15. |
| [19] | Nolan T, Hands R E, Bustin S A. Quantification of mRNA using real-time RT-PCR. Natural Protocols, 2006, 1(3): 1559-1582. |
| [20] | Rueckert A, Cary S C. Use of an armored RNA standard to measure microcystin synthetase E gene expression in toxic Microcystis sp. by reverse-transcription qPCR. Limnology Oceanography Methods, 2009, 7: 509-520. |