 2015, Vol. 35
2015, Vol. 35文章信息
- 张东菊, 左平, 邹欣庆
- ZHANG Dongju, ZUO Ping, ZOU Xinqing
- 基于加权Ripley's K-function的多尺度景观格局分析——以江苏盐城滨海湿地为例
- Multi-scale analysis of landscape characteristics in coastal wetlands in Yancheng, Jiangsu, based on the weighted Ripley's K-function method
- 生态学报, 2015, 35(8): 2703-2711
- Acta Ecologica Sinica, 2015, 35(8): 2703-2711
- http://dx.doi.org/10.5846/stxb201307191915
-
文章历史
- 收稿日期:2013-07-19
- 修订日期:2014-10-12






2. 南京大学海岸与海岛开发教育部重点实验室, 南京 210046
2. Ministry Education Key Laboratory for Coast and Island, Nanjing University, Nanjing 210046, China
空间异质性、生态学过程和尺度的关系一直以来都是景观生态学研究的热门命题和重要研究内容,而景观格局是景观空间异质性的外在表现,也是各种生态过程在不同大小空间尺度上相互作用的结果[1, 2, 3, 4],因此,对景观格局-生态过程关系的全面认知和解析理应基于不同的研究尺度。目前针对盐城滨海湿地的景观格局研究多基于单一尺度,一定程度上限制了我们在多尺度空间上对景观类型及其生态过程的理解和分析,因此同时在多尺度上开展该区域景观的空间分布特征研究,成为理解该区域格局和生态过程相互作用的关键和重点[5, 6]。已有文献[7, 8, 9, 10]对盐城湿地景观类型的变化分析多采用景观格局指数,转移矩阵(优势转移过程及其贡献率)以及土地利用动态模型等方法,在这其中,尤以景观指数法应用最多。然而,这些基于空间非连续性数据的传统景观指数法大都基于单一尺度,且众多景观指数在表征景观格局特征方面具有片面性,在指示生态学过程方面具有局限性。在不改变源数据空间尺度的前提下,不能同时在宏观和微观等连续尺度上揭示景观格局的特征和生态过程。因此有必要应用多尺度景观格局分析方法来剖析景观格局与生态过程关系[11]。当前,多尺度的研究方法和理念被广泛应用到林地、草地以及海洋等生态系统的种群分布格局研究上[12, 13, 14, 15, 16, 17, 18, 19, 20, 21],实践证明其可以有效地揭示出研究物种在多个尺度上的生态学特性和过程。
Ripley′s K-function恰恰是一种可以在多尺度上分析研究对象空间分布格局的有效手段,其提供的空间特征尺度和空间分布强度两个指数可以很好地表征景观类型的斑块大小和空间聚集程度,具有很强的生态学指示意义,且在景观格局上的研究上已显示出简单,高效等诸多优点[22, 23, 24, 25]。Ripley′s K-function是一种基于点状数据的格局分析方法,因此在计算之前需要将各种格式的数据进行点状处理。考虑到所涉及的研究区域及研究对象均较小,相关文献大多采取将研究对象直接点状化的方式进行空间格局计算,而忽略研究对象的分布范围等属性。在景观空间分析中,传统的Ripley′s K-function主要是在ArcGIS中进行以下两种方式的点状化处理:一是将土地利用类型与随机点结合。高凯等[22]利用create random points,通过生成的随机点跟矢量图叠置分析,将土地的利用类型赋值给随机点,这些赋值后的随机样点将代表各景观类型进行点格局计算。笔者认为这种方法与数据源本身的提取精度关系较大,随机点的数量较难控制,该方法随机性稍显大些。例如,对某些大面积分布的景观来说,其在地理信息系统图层中虽然有明显的聚集特征,但仅靠生成的几个随机点会将聚集性弱化甚至导致最终无法被检测出来;二是将面状格式的土地利用或者景观数据直接转为点状数据。王金亮等[23]利用Feature to Point命令,将林地斑块面状数据的质心转换为点状数据,通过将点状数据与生态屏障区图层叠加,得到研究区林地景观点格局样点数据。然而,对于某些零星分布的景观来说,只考虑样点质心位置不考虑样点本身的分布范围等属性值,容易造成局部“伪聚集性或过聚集”现象。
鉴于本研究所涉及的研究对象及其所在区域的空间尺度均相对较大,因此不能将景观类型的空间分布范围属性直接忽略,为弥补传统Ripley′s K-function方法在景观类型点状化过程中的粗糙和随机性等缺陷,本文考虑将研究对象的面积属性以权重的方式纳入到空间分布类型的计算中,并以盐城滨海湿地为例,对其格局20年间的变化进行了分析。首先,将整个研究区划分为宽度一致长度不等的多个连续样带,以达到对原始面状数据进行简化的目的;其次,提取各湿地类型的空间位置(某类型湿地所在样带的质心坐标)和分布范围属性值(某类型湿地在样带内的分布面积);然后,将所得信息应用到加权Ripley′s K-function方法中,从而避免了仅考虑样点空间位置信息而造成的“伪分散”(尤其是针对大面积块状或者带状分布的湿地类型)或者“伪聚集”(特别是对破碎化严重,零星分布的湿地类型)等现象。
1 研究区与研究方法 1.1 研究区概况本研究区域的地理范围包括盐城沿海响水、滨海、射阳、大丰、东台5个县(市)的潮间带滩涂、港河口、部分潮下带以及由堤内的耕作农田向堤外滩涂过渡区的芦苇鱼塘、林地、盐场、撂荒地等[7](图 1)。该区域生物多样性异常丰富,是东北亚-澳大利亚候鸟迁徙的重要通道和停歇地,该区域设立的江苏盐城珍禽自然保护区,是全球最大的丹顶鹤越冬基地,也是亚洲最大的沿海滩涂湿地型保护区[26]。然而,随着滩涂的大规模围垦、资源的过度开发利用以及环境污染,该区域也面临着景观破碎化,生态功能退化等一系列问题。因此对该区域湿地景观的空间分布特征及其变化的研究具有较强的现实意义,可以为开展湿地生物多样性监测保护及恢复提供更详细的定量指标。
研究区内的自然湿地(从植被类型角度阐述)由海到陆的演替层次十分明显,且部分地区仍保留着海岸带原生植被演替序列,过渡的类型依次为:光滩或泥滩(无植被)-米草滩(Spartina alterniflora,1982年形成最大的互花米草盐沼生态系统[27])-碱蓬滩(Suaeda glauca)-獐茅滩(Aeluropus littoralis)-白茅草滩(Rhizoma Imperatae)或芦苇滩(Phragmites communis)。
结合当地滩涂开发利用现状和规划方案,本文选取20年来在空间分布尺度和强度上变化幅度较大的几大湿地类型为重点研究对象。具体来说,自然景观类型的研究主要集中于獐茅滩、芦苇滩、碱蓬滩、互花米草滩;人工湿地类型主要包括农业用地、水产养殖塘和盐田。
1.2 数据来源及处理本研究以覆盖该区域的1987、1992、1997、2002和2007年的Landsat影像(分辨率为30 m)作为基本信息源,并选用TM5,TM4和TM3三个波段进行标准假彩色合成,结合目视判读和ERDAS 9.1中的非监督分类模块,对典型的湿地景观类型进行初步判断和解译,再通过实地调查验证提高结果精度,进而最终识别出主要的自然湿地景观类型和人工湿地景观类型[7]。分类结束后,提取的景观类型的结果图件导入到ArcGIS10.0软件中进行矢量化和各类空间分析,建立1987—2007年间各湿地类型的空间格局数据。同时,为了满足Ripley′s K-function对数据格式的要求,首先,对原有的面状数据进行了条带化概括处理(将各类零散分布且面积不大的湿地类型统一纳入到样带内),将研究区从北到南划分为260个样带(图 1),其中,样带的宽度为1 km,长度不等(切割于实际的边界范围)。然后,将各湿地景观类型数据与样带图层进行叠加,得到研究区各个景观类型的样带分布数据,与此同时,分别统计1987—2007年落入各样带内的各湿地类型的面积(z)。在此基础上,将各个样带进行点状化处理,提取各样带的质心坐标(x,y)和各湿地类型在样带内的分布范围即面积(z),为接下来的加权Ripley′s K-function运算提供空间位置信息和加权变量。

|
| 图 1 研究区位置及其样带划分图 Fig.1 Location of study area and belt transects map |
Ripley′s K-function是点格局分析的一种重要方法,它可以反映点格局(要素质心)在整个空间幅度范围内的变化特征,能够检测研究对象的空间分布特征在邻域大小发生变化时是如何变化的[28, 29]。本文在传统Ripley′s K-function计算公式(1)的基础上,采用改进的加权Ripley′s K-function[30]来分析各湿地类型的空间分布特征,其计算公式如式(2)所示:
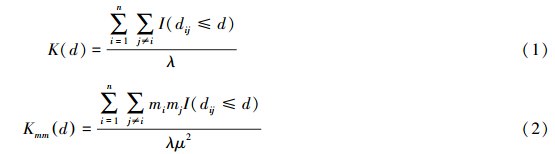
式中,点的位置信息由各湿地类型所在样带的质心点的坐标来表征。d为空间尺度,dij为研究对象在质心点i与质心点j之间的距离,且i,j=1,2,3,…,n(i≠j);n为研究对象的个数;当dij≤d时,I=1;当dij>d时,I=0;λ为假设研究对象在研究区内呈均匀分布时的空间密度值。由公式(1)可知,传统方法根据位置信息统计出不同距离尺度内研究对象的数目,进而判读该研究对象的空间分布状态。mi与mj为权重变量,是研究对象在质心点i和质心点j上所关联的属性值-即该研究对象的分布范围(z),μ是研究对象所关联的属性值的均值。为使期望值线性化并保持方差稳定[22],同时也便于直观地理解现实中研究对象的空间分布格局,Besag[31]重新标准化方程构造了指标Lmm(d):

式中,假设样点呈完全空间随机分布(CSR),Lmm(d)的期望为0。当Lmm(d)大于0,说明景观类型呈聚集分布;Lmm(d)小于0,说明景观类型呈现分散分布的趋势;Lmm(d)等于0,则表示景观类型呈随机分布。
本文的加权Ripley′s K-function计算是在 Matlab R2011a软件中运行,以样带中心点的经度、纬度和各湿地类型所覆盖的面积分别作为空间位置变量和加权变量,分别计算1987—2007年各湿地类型的空间特征尺度和空间分布强度。同时,指数偏离随机性的显著性检验采用蒙特卡罗模拟法(M-C),置信度取99%。运算结束后,首先根据模拟法检验所生成的上、下包迹线曲线判断各湿地的空间分布类型:若景观类型的Lmm(d)值大于上包迹线,则该湿地类型呈现显著聚集分布;若Lmm(d)值小于下包迹线,则对应的湿地类型呈现分散分布;若Lmm(d)值落入上下包迹线区间范围内,说明相关湿地类型的空间分布是完全随机分布。其次,当景观类型呈现聚集的空间分布状态时,提取两个重要的空间分布特征数:空间分布强度和空间特征尺度。空间分布强度,即某一类型的湿地在空间上聚集的规模,是由Lmm(d)曲线中的第一个有效峰值来度量,峰值越高,所对应的湿地类型在空间上的聚集强度或规模就越大;空间特征尺度,是Lmm(d)曲线中第1个有效峰值所对应的d值,d值愈大,所计算的湿地类型在空间上的聚集尺度越大。
2 结果分析 2.1 湿地景观的面积及其样带变化分析从盐城滨海湿地景观面积的百分比变化来看(表 1)[7],自然湿地仍是盐城滨海湿地中的主要景观类型,但部分典型自然湿地的总面积和比重都有明显下降的趋势,且景观中的植被群落类型多样性明显减少(如獐茅滩湿地在2007年影像中已经基本检测不到),并有逐渐向单一优势种群过渡的趋势(在盐城滨海滩涂,互花米草已取代碱蓬成为先锋植物群落并迅速扩散),相对而言,人工湿地的面积和比重却有着显著上升的趋势。图 2中自然湿地和人工湿地的面积在大部分样带内的增减量变化(增减量=2007年的自然或者人工湿地面积-1987年的湿地面积)更好地印证了二者存在着此消彼长的关系。具体来说,176个样带中的自然湿地处于减量区(以0值为分界线,值大于0代表该类型湿地有增量;小于0代表该湿地类型有减量),193个样带中的人工湿地位于增量区。因篇幅原因,各个湿地类型的面积增减量未在此列出。
| 湿地类型Wetland Landscapes | 1987 | 1992 | 1997 | 2002 | 2007 |
| 獐茅滩Aeluropus littoralis/% | 5.64 | 4.78 | 2.91 | 1.06 | 0.00 |
| 芦苇滩Phragmites communis/% | 2.33 | 2.85 | 2.34 | 2.13 | 2.33 |
| 碱蓬滩Suaeda glauca/% | 6.66 | 5.76 | 5.19 | 3.49 | 2.85 |
| 互花米草滩Spartina alterniflora/% | 0.01 | 0.21 | 0.96 | 2.04 | 2.12 |
| 水产养殖塘Aquacultural ponds/% | 2.78 | 2.71 | 3.96 | 9.14 | 11.23 |
| 盐田Salt fields/% | 6.53 | 6.67 | 6.22 | 5.69 | 5.18 |
| 农业用地Agricultural land/% | 3.57 | 5.45 | 7.44 | 7.47 | 11.55 |
| 其它Others/% | 72.03 | 71.63 | 71.63 | 70.66 | 66.50 |

|
| 图 2 1987—2007年研究区湿地在各样带内的面积变化量 Fig.2 Variations of wetland area in each belt of the study area from 1987 to 2007 |
由图 3和表 2可以看出,1987—2007年间,各年份各湿地景观的Lmm(d)曲线基本都在上包迹线以上,表明在设定的多个不同空间尺度上,无论何种类型的湿地,均表现出显著的聚集分布特征。自然湿地方面,碱蓬 滩和獐茅滩的空间特征尺度和空间分布强度都有明显的下降趋势,从大范围的高度聚集向多个尺度的局部小范围聚集分布发展,獐茅滩尤甚。碱蓬滩的空间特征尺度从1987年的81 km 下降到2007年的11 km,与此同时,其空间分布强度也从160.65下降到16.93,曲线中出现更多的小峰值,说明研究期间碱蓬滩从开始的较大斑块转变为两个尺度上的较小斑块。獐茅滩曲线基本呈现出与碱蓬滩一致的变化趋势,其空间特征尺度也不断萎缩,从1987年的75 km到2007年在遥感影像中消失,且其空间聚集强度也不断下降,从94.35到基本消失。芦苇滩的空间特征尺度在1987—1997期间变化不大,基本在8 km左右,到了2002年尺度上升为26 km,到2007年再次下降为12 km;与此同时,空间分布强度表现出先稳步下降后于2007年显著上升的趋势,但相对于1987年仍有减弱,从1987年的15.39减小为2007年的12.61,由此可判断芦苇湿地的空间分布形态除了在2002年出现较大波动以外,其他年份的聚集尺度和规模基本保持稳定。互花米草自1992年引进以来,呈现不断蔓延扩散的趋势,甚至一度被作为入侵种进行研究,与其他自然湿地的空间分布变化相反,该湿地类型是自然湿地中唯一聚集性和分布规模逐渐增强的类型,其空间特征尺度从1992年的32 km上升到2007年的73 km,空间分布强度指数从1992年的5.81上升到2007年的39.04,互花米草滩相对均匀地逐年快速增加使其在局部范围内迅速聚集壮大并蔓延开来,逐渐由局部的小聚集演变为大范围的高度聚集。

|
| 图 3 1987—2007年各湿地景观类型的 Ripley′s K-function 指数变化图 Fig.3 Ripley′s K-function indexes changes of different wetland landscapes from 1987 to 2007 |
| 湿地类型 Wetland Landscapes | 空间特征尺度Patch radius/km | 空间分布强度Crowding | ||||||||
| 1987 | 1992 | 1997 | 2002 | 2007 | 1987 | 1992 | 1997 | 2002 | 2007 | |
| 獐茅滩Aeluropus littoralis | 75 | 79 | 49 | 7 | 94.35 | 82.09 | 35.12 | 8.49 | ||
| 芦苇滩Phragmites communis | 8 | 8 | 8 | 26 | 12 | 15.39 | 7.94 | 11.74 | 13.46 | 12.61 |
| 碱蓬滩Suaeda glauca | 81 | 44 | 54 | 11 | 11 | 160.65 | 35.15 | 44.16 | 11.41 | 16.93 |
| 互花米草滩Spartina alterniflora | 32 | 54 | 64 | 73 | 5.81 | 17.83 | 39.22 | 39.04 | ||
| 水产养殖塘Aquacultural ponds | 21 | 12 | 36 | 80 | 82 | 10.22 | 7.89 | 17.60 | 132.20 | 126.20 |
| 盐田Salt fields | 30 | 28 | 27 | 29 | 31 | 125.53 | 120.27 | 173.91 | 131.51 | 136.75 |
| 农业用地Agricultural land | 23 | 24 | 34 | 90 | 89 | 39.83 | 30.43 | 34.10 | 74.20 | 109.22 |
人工湿地基本表现出与自然湿地相反的空间分布趋势,其呈现出从多个尺度上的局部小范围聚集到单尺度上的大范围高度聚集分布的变化。这点在水产养殖塘和农业用地两个湿地类型方面表现的尤为突出,二者的Lmm(d)曲线峰值均出现了明显的变化,表明相应湿地类型正从多个不同尺度上的弱聚集逐渐演变为单个较大尺度上的强聚集。水产养殖塘的空间特征尺度从1987年的21 km上升到2007年的82 km,空间分布强度从1987年的10.22猛烈上升到2007年的126.20;农业用地的空间特征尺度从1987年的23 km上升到2007年的89 km,空间分布强度从1987年的39.83迅速上升到2007年的109.22,和水产养殖塘基本表现出一致的空间变化趋势;盐田的空间特征尺度变化不大,集中在27—31 km之间,空间分布强度整体变化幅度较小,且在1997年达到最高,表明其在局部小范围的聚集尺度和规模基本保持不变。
3 讨论左平等[7]采用斑块个数、斑块平均面积和平均斑块最近距离3个景观指数对受人类活动干扰较强的湿地类型进行了变化分析,本文的相关结论与其结论中对各湿地景观类型变化趋势的表述基本一致。对照来看,其结论指出盐城滨海湿地的景观变化总体上有破碎化程度加剧的趋势,自然湿地方面,獐茅滩和碱蓬滩的面积均持续减少,芦苇滩也呈现出先增加后减少的情况,此外互花米草滩无论是斑块个数还是面积均是逐年 递增,而这恰恰和本研究中自然湿地在多尺度的变化相吻合,如獐茅滩和碱蓬滩在多尺度上的聚集性减弱及空间分布强度不断下降,芦苇滩的特征尺度先稳步下降后迅速增加再迅速减少的趋势,互花米草滩聚集性迅速增强等;而人工湿地方面,其结论中鱼塘的面积和平均斑块最近距离有大范围增加的趋势,盐田的斑块平均面积出现了先减少后增加的趋势,这也和本研究中所对应的人工湿地的聚集尺度和规模的变化趋势相似。此外,张华兵等[32]指出盐城国家级自然保护区核心区湿地景观格局经历了从1987年一定的规律带状分布到2007年的镶嵌格局转变的过程,这与本文中部分自然湿地减少(如獐茅滩、碱蓬滩等),从规则的条带状分布到聚集特征尺度骤减,零散分布的趋势基本相同。
本文借助点格局理论,依托于遥感和地理信息技术,应用加权Ripley′s K-function分析了盐城滨海湿地景观格局20年间的变化情况,该方法最大限度地利用点格局的位置和距离信息,同时考虑了各样点本身的分布范围。结论表明,加权Ripley′s K-function方法能够在一定程度上减少和避免传统Ripley′s K-function方法所造成的两种伪空间分布结果:一方面,在传统分析方法中,由于大面积成片状或者带状分布的湿地类型直接点状化往往只能生成较少数量的样点,导致某一类型的湿地虽然在GIS图层上有明显的聚集变化,但却被传统方法解读为随机或者分散分布,而加权方法由于考虑了样点的分布范围,体现出了研究对象在连续空间上的面积变化情况,因此可以将该种类型湿地在多尺度上的聚集变化表征出来,本研究中大面积成片分布的人工湿地的提取和空间分布结论证明了加权方法在这方面的科学性;另一方面,对因破碎化比较严重而生成众多样点的湿地类型而言,传统方法容易生成“伪聚集或过聚集”[30]现象,而改进方法中由于加入了众多样点的面积值作为权重变量,使得运算结果更加接近于现实中的分布状态,如本方法较好地避免了破碎化严重的獐茅滩的“过集聚”分布的现象,揭示了其在聚集尺度和规模上逐渐萎缩甚至几近消失的变化情况。另外需要指出的是,在本研究中,改进方法中的尺度变化指的各类型湿地在南北方向上的距离差异,距离计算基于的是样带质心组成的一维空间,长度不等及其样带内的差异导致的实际距离差异在本研究中予以忽略,考虑到研究区域的差异性,将该方法应用到其他案例时,样带的大小确实会是一个值得严谨考虑的问题,其在一定程度上影响到质心的位置及其湿地类型的多少,且样带内的差异同样也值得探讨。本文未对此进行详细的说明和进一步的分析,略显不足,今后将加以弥补和完善。实践证明,利用改进的Ripley′s K-function可以很好地反映出各类型湿地在1987—2007年间在多尺度的空间变异,该方法能简单有效地体现湿地格局从宏观到微观的空间分布特征,并能给出空间分布格局的最大聚集强度及其对应的空间特征尺度,且计算结果和实际的空间聚集变化趋势保持一致,今后可以作为持续分析盐城滨海湿地景观变化及其生态过程的重要工具。
景观的空间格局是各种生态过程在不同尺度上的长期相互作用的结果,因此驱动力的分析也应基于不同的研究尺度。针对滨海湿地景观演变的驱动力分析目前已有较多论述[7, 32, 33],且论述基本一致,主要的分析集中在自然地理等生态机制(海岸地貌侵蚀与淤积过程以及植被的自然演替过程)和人为干扰等要素(米草物种的引进和扩散,各时期不同程度的滩涂围垦等经济开发活动以及区域人口增长等)两个方面。这两大因素相辅相成,密不可分,共同决定了滨海湿地在各个时间段内区域潜在的异质性空间格局,而这些格局最终也通过各湿地类型的空间特征尺度和空间分布强度在本研究中充分表现出来。如自然湿地方面,以互花米草为例,由于其疯狂的繁殖能力,在局部区域迅速聚集为较大斑块,并逐渐扩散开来,表现为聚集尺度和规模的激增;人工湿地方面,以水产养殖塘为例,过度捕捞导致许多渔民被迫走上滨海养殖道路,表现为水产养殖塘在2002和2007年以来的聚集空间特征尺度和规模的大幅上升。鉴于研究区的各类景观类型在多尺度上受到来自各类海洋物理,生物生态,化学以及人为等驱动因子的共同作用,且在多个尺度上的作用强度以及方向不尽一致,从格局到过程的推绎仍然面临一些挑战,因此本文只是揭示出各类自然湿地类型在多个空间特征尺度上的变化情况,未对各类生态过程在多尺度空间上如何作用于湿地类型进行详细分析,对该部分内容进行专门分析以便为管理层提供更好的量化指标将成为下一步的研究重点。
致谢: 感谢Charles “Pete” H. Peterson教授对英文写作的帮助。
| [1] | Zhang S L, Sadras V, Chen X P, Zhang F S. Water use efficiency of dryland wheat in the Loess Plateau in response to soil and crop management. Field Crops Research, 2013, 151: 9-18. |
| [2] | Gan Y T, Siddique K H M, Turner N C, Li X G, Niu J Y, Yang C, Liu L P, Chai Q. Chapter Seven-Ridge-furrow mulching systems-an innovative technique for boosting crop productivity in semiarid rain-fed environments. Advances in Agronomy, 2013: 118: 429-476. |
| [3] | Wang F H, He Z H, Sayre K, Li S D, Si J S, Feng B, Kong L G. Wheat cropping systems and technologies in China. Field Crops Research, 2009, 111(3): 181-188. |
| [4] | 张德奇, 廖允成, 贾志宽. 旱区地膜覆盖技术的研究进展及发展前景. 干旱地区农业研究, 2005, 23(1): 208-213. |
| [5] | 李凤民, 王静, 赵松岭. 半干旱黄土高原集水高效旱地农业的发展. 生态学报, 1999, 19(2): 259-264. |
| [6] | Han J, Jia Z K, Wu W, Li C S, Han Q F, Zhang J. Modeling impacts of film mulching on rainfed crop yield in Northern China with DNDC. Field Crops Research, 2014, 155: 202-212. |
| [7] | 马忠明, 杜少平, 薛亮. 不同覆膜方式对旱砂田土壤水热效应及西瓜生长的影响. 生态学报, 2011, 31(5): 1295-1302. |
| [8] | 张永涛, 汤天明, 李增印, 王开增. 地膜覆盖的水分生理生态效应. 水土保持研究, 2001, 8(3): 45-47. |
| [9] | 孙惠民, 程满金, 郑大玮, 张建新. 北方半干旱集雨补灌旱作区节水农业发展模式. 应用生态学报, 2005, 16(6): 1072-1076. |
| [10] | 王俊, 李凤民, 宋秋华, 李世清. 地膜覆盖对土壤水温和春小麦产量形成的影响. 应用生态学报, 2003, 14(2): 205-210. |
| [11] | 李儒, 崔荣美, 贾志宽, 韩清芳, 路文涛, 侯贤清. 不同沟垄覆盖方式对冬小麦土壤水分及水分利用效率的影响. 中国农业科学, 2011, 44(16): 3312-3322. |
| [12] | 张淑芳, 柴守玺, 蔺艳春, 常磊, 杨长刚. 干旱年份地膜覆盖模式对春小麦土壤水分和产量的影响. 中国农业气象, 2011, 32(3): 368-374. |
| [13] | 陈辉林, 田霄鸿, 王晓峰, 曹玉贤, 吴玉红, 王朝辉. 不同栽培模式对渭北旱塬区冬小麦生长期间土壤水分、温度及产量的影响. 生态学报, 2010, 30(9): 2424-2433. |
| [14] | 王琦, 张恩和, 李凤民. 半干旱地区膜垄和土垄的集雨效率和不同集雨时期土壤水分比较. 生态学报, 2004, 24(8): 1820-1823. |
| [15] | 李廷亮, 谢英荷, 任苗苗, 邓树元, 单杰, 雷震宇, 洪坚平, 王朝辉. 施肥和覆膜垄沟种植对旱地小麦产量及水氮利用的影响. 生态学报, 2011, 31(1): 212-220. |
| [16] | 廖允成, 温晓霞, 韩思明, 贾志宽. 黄土台原旱地小麦覆盖保水技术效果研究. 中国农业科学, 2003, 36(5): 548-552. |
| [17] | 薛澄, 王朝辉, 李富翠, 赵护兵, 周玲, 李小涵. 渭北旱塬不同施肥与覆盖栽培对冬小麦产量形成及土壤水分利用的影响. 中国农业科学, 2011, 44(21): 4395-4405. |
| [18] | Gao Y H, Xie Y P, Jiang H Y, Wu B, Niu J Y. Soil water status and root distribution across the rooting zone in maize with plastic film mulching. Field Crops Research, 2014, 156: 40-47. |
| [19] | 王喜庆, 李生秀, 高亚军. 地膜覆盖对旱地春玉米生理生态和产量的影响. 作物学报, 1998, 24(3): 348-353. |
| [20] | Qin S H, Zhang J L, Dai H L, Wang D, Li D M. Effect of ridge-furrow and plastic-mulching planting patterns on yield formation and water movement of potato in a semi-arid area. Agricultural Water Management, 2014, 131: 87-94. |
| [21] | Ramakrishna A, Tam H M, Wani S P, Long T D. Effect of mulch on soil temperature, moisture, weed infestation and yield of groundnut in northern Vietnam. Field Crops Research, 2006, 95(2/3): 115-125. |
| [22] | 王红丽, 张绪成, 宋尚有, 马一凡, 于显枫, 柳燕兰. 旱地全膜双垄沟播玉米的土壤水热效应及其对产量的影响. 应用生态学报, 2011, 22(10): 2609-2614. |
| [23] | Liu C A, Zhou L M, Jia J J, Wang L J, Si J T, Li X, Pan C C, Siddique K H M, Li F M. Maize yield and water balance is affected by nitrogen application in a film-mulching ridge-furrow system in a semiarid region of China. European Journal of Agronomy, 2014, 52(Part B): 103-111. |
| [24] | 李巧珍, 李玉中, 郭家选, 刘晓英, 徐春英. 覆膜集雨与限量补灌对土壤水分及冬小麦产量的影响. 农业工程学报, 2010, 26(2): 25-30. |
| [25] | Li F M, Wang J, Xu J Z. Plastic film mulch effect on spring wheat in a semiarid region. Journal of Sustainable Agriculture, 2005, 25(4): 5-17. |
| [26] | 李凤民, 鄢珣, 王俊, 李世清, 王同朝. 地膜覆盖导致春小麦产量下降的机理. 中国农业科学, 2001, 34(3): 330-333. |
| [27] | 王红丽, 张绪成, 宋尚有, 马一凡, 于显枫. 西北黄土高原旱地全膜双垄沟播种植对玉米季节性耗水和产量的调节机制. 中国农业科学, 2013, 46(5): 917-926. |
| [28] | 王红丽, 张绪成, 宋尚有. 半干旱区旱地不同覆盖种植方式玉米田的土壤水分和产量效应. 植物生态学报, 2011, 35(8): 825-833. |
| [29] | 于振文. 作物栽培学各论(北方本). 北京: 中国农业出版社, 2013: 54-55. |
| [30] | 龚月桦, 杨俊峰, 王俊儒, 李生秀, 刘普灵. 覆膜对小麦14C-储备物在灌浆期转运分配的影响. 中国农业科学, 2007, 40(02): 258-263. |
| [31] | Du Y J, Li Z Z, Li W L. Effect of different water supply regimes on growth and size hierarchy in spring wheat populations under mulched with clear plastic film. Agricultural Water Management, 2006, 79(3): 265-279. |
| [32] | 杨长刚, 柴守玺, 陈恒洪, 常磊, 岳云. 不同覆膜种植方式对旱地冬小麦干物质积累与分配的影响. 甘肃农业大学学报, 2012, 47(6): 49-56. |
| [33] | 潘晓云, 王永芳, 王根轩, 曹琴东, 王静. 覆膜栽培下春小麦种群的生长冗余与个体大小不整齐性的关系. 植物生态学报, 2002, 26(2): 177-184. |