 2015, Vol. 35
2015, Vol. 35文章信息
- 祝英, 熊俊兰, 吕广超, ASFABatool, 王兆滨, 李朴芳, 熊友才
- ZHU Ying, XIONG Junlan, LÜ Guangchao, ASFA Batool, WANG Zhaobin, LI Pufang, XIONG Youcai
- 丛枝菌根真菌与植物共生对植物水分关系的影响及机理
- Effects of arbuscular mycorrhizal fungi and plant symbiosis on plant water relation and its mechanism
- 生态学报, 2015, 35(8): 2419-2427
- Acta Ecologica Sinica, 2015, 35(8): 2419-2427
- http://dx.doi.org/10.5846/stxb201306091556
-
文章历史
- 收稿日期:2013-06-06
- 网络出版日期:2014-05-16






2. 甘肃省科学院生物研究所, 兰州 730000;
3. 兰州大学信息科学与工程学院, 兰州 730000
2. Institute of Biology, Gansu Academy of Sciences, Lanzhou 730000, China;
3. School of Information Science and Engineering, Lanzhou University, Lanzhou 730000, China
丛枝菌根真菌(AMF)与大多数陆生植物关系密切,与植物形成共生体后,通过进一步改良土壤结构影响根际微生物群落结构、宿主植物的生理生态功能、物质元素生物地球化学循环、陆生生态系统结构和功能等,并对气候环境变化发挥反馈作用[1, 2, 3]。菌根与植物水分代谢密切相关,1885年Frank首次提到“mykorhiza”一词,并指出水分和营养物质经过外生菌丝运送到植物根尖,植物与真菌之间形成互惠共生关系[4]。1971年Safir等人[5]将接种Glomus mosseae可降低大豆水运输的阻力,在不改变根系形态的情况下,同时促进地上部分生长,其实验结果在Science上发表,从此开始了丛枝菌根真菌与植物水分关系的系统研究。
旱地农业系统主要指年平均降雨量在300—550 mm的生态系统和农业区域,水分是该区域作物生长的主要限制因子。大量研究表明,干旱条件下,AMF与植物共生可以调节宿主植物根源信号合成[6, 7],提高渗透调节能力[8, 9, 10, 11]、氧化酶活性[12, 13, 14]和水分吸收利用效率[15, 16]等,赋予作物优良的耐旱性和生长特性,这为旱地农业向高产潜能和高耐旱性发展提供一种有效途径。虽然已经公认AMF能改善宿主植物水分代谢和增强抗旱性的功能,但是其作用机制还尚未形成共识和系统总结。特别是干旱胁迫下,AMF对宿主非水力信号产生及根冠通讯的影响更缺乏深入系统认识。本文从干旱胁迫下AMF调节宿主植物各种生理生态指标和可能的机理入手,着重综述并讨论AMF提高土壤水分吸收、运输和对宿主根源信号产生及根冠通讯的影响,以深入了解水分胁迫下,AMF促进水分吸收和提高宿主对非水力信号的响应机制,发挥其在干旱农业生产中的应用潜力。
1 菌根共生对宿主干旱适应性的影响植物通过改变自身的生理、形态和物候特性等方式对干旱胁迫做出反应。AMF与植物共生能改变水分运移、生理和形态特性,进而影响植物对干旱的适应性。1979年Reid指出菌根有利于干旱胁迫下的植物逃避干旱损伤。大量的研究证实了这个观点,接种AMF的植物与没有接种AMF的相比,在干旱胁迫下AMF明显提高宿主植物氮和碳的同化速率,表现在可溶性蛋白、氨基酸、含氮酶和组织氮素含量较高[17, 18];提高酶活性[10];降低脯氨酸积累和其他含氮物质引起的氧化损伤[8, 9, 10, 11];根系较大[19];增加蜡质层保护叶片,增强耐脱水性能[20];增加土壤水分摄取,降低永久萎焉点的相应土壤含水量,并能迅速从干旱胁迫状态恢复正常等[21];并且AMF与植物共生提高宿主对干旱的耐受性与营养元素的吸收密切相关。80%的菌根研究表明,在干旱胁迫下接种AMF明显促进宿主植物生长,这表现在AMF有利于植物对营养物质的吸收,对提高耐旱性发挥重要作用;在充分供水条件下,AMF植物比没有接种AMF的植物生长快速,表明AMF能促进营养元素的吸收,缓解磷元素胁迫;而叶片中磷的含量可以控制保卫细胞对ABA的敏感性,进而对调节气孔行为发挥作用[22]。
AMF通过改变植物系统耐旱性和细胞耐旱性两方面来提高宿主对干旱的耐受性。系统耐旱性主要是指在干旱胁迫下,AMF有利于宿主植物改变形态结构,例如株型变小、根系变大、调整生物量分配、关闭气孔、分泌球囊霉素团聚土壤水分等方式以达到防止植物脱水的功能;细胞耐旱性是指干旱胁迫下,AMF有利于宿主植物快速调节细胞内的生理生化物质,以表现出抵御干旱胁迫的能力,例如提高氧化酶活性、积累蛋白质、脯氨酸、甜菜碱等渗透调节物质,提高植物耐干旱的耐受性。据此,吴强盛总结了AMF提高宿主干旱适应性的可能机制模式框架[23],为了更直观的了解菌根共生过程中AMF发挥的作用,在此基础上对机制模式图进一步修改(图 1)。该图全面概括了40多年来AMF提高宿主植物抗旱性的生理生态机制研究成果。但是,所有生理生态反应最基础的应该是信号物质和信号传递方式与速度,没有信号物质积累和传递,宿主不会作出相应的生理生态改变,因此信号物质产生和传递,是宿主对干旱胁迫作出生理生态响应最原始的推动力。下面将从AMF有利于宿主植物水分吸收、转运和非水力信号产生及传递入手,讨论干旱胁迫下,AMF影响宿主植物信号调节和传递的可能机制。

根系吸水主要依靠两种动力:横向根压和纵向蒸腾拉力。三种吸水途径:质外体途径(指水分通过细胞壁、细胞间隙等没有细胞质部分的移动,阻力小,所以这种移动方式速度快),跨膜途径(指水分从一个细胞移动到另一个细胞,要两次通过质膜,还要通过液泡膜,故称跨膜途径)和共生体途径(指水分从一个细胞的细胞质经过胞间连丝,移动到另一个细胞的细胞质,形成一个细胞质的连续体,移动速度较慢)。1971年Safir首次发表接种Glomus mosseae可降低大豆水运输的阻力的观点[5],而Sands和Theodorou认为应将水分运输分成两部分,一部分是水分从土壤运输到根系,即横向根压;另一部分是植物体内的水分运输,即纵向蒸腾拉力。他们在对松树苗的研究发现,干旱胁迫时,AMF植物根系土壤的阻力更大,这可能与AMF植物根系结构有关[24]。利用示踪技术研究松树苗菌根根系和非菌根根系水分的吸收转运差异,结果菌根根系的水分运移速度明显高于非菌根根系,为菌根根系促进水分运移提供直接证据,接着又对菌根和非菌根根系结构观察,为菌根根系降低水分运输阻力提供证据[25, 26, 27, 28]。Khalvati等人在干旱胁迫下,利用大麦的分根实验来衡量菌根菌丝对宿主吸水的贡献,结果有4%的水分通过AMF菌丝运送到宿主根部[29]。Ruth等人利用高分辨率在线水含量传感器定量分析菌根真菌菌丝对水分的吸收效率,结果表明,菌丝提高总水分吸收率的20%[30]。有关菌丝对水分的运输能力报道不一,一方面是由于研究菌丝对水分运输能力的方法不一致,另一方面可能因宿主植物和AMF的不同而存在差异。
根据土壤根系吸水模型Ohm′s law,公式计算:
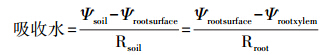
式中,R是水分的运输阻力;Ψsoil是土壤水势;Ψroot surface是根系表面水势;Ψroot xylem是根木质部水势。AMF外延菌丝的水分运输提高宿主根部的横向压力,也就是提高了土壤和根系表面的水势差,降低水分在土壤中的运输阻力,因此AMF有利于宿主植物的水分吸收。
同时,AMF与植物共生通过增加蒸腾速率或(和)气孔导度等方式,提高植物的纵向蒸腾拉力,有利于土壤水分的吸收。早在1980年Levy和Krikum在柑橘根部接种G. fasciculatus的实验发现,在水分胁迫和胁迫解除期间,菌根增强宿主植物的蒸腾速率和气孔导度,有利于水分更通畅快速的运输[16]。接着Allen研究表明,接种和不接种AMF植物的叶面积和根长没有显著差别,在根和叶水势不变的情况下,菌根植物叶片蒸腾速率提高100%,整个植株体内水分运输阻力减少50%,AMF根外菌丝到根部水分运输速率每个侵入位点约为2.8×10-5 mg/s[31]。之后大量的研究证实菌根植物与非接菌植物相比,宿主植物的叶片气孔导度、蒸腾速率和光合作用均有显著提高[15, 32, 33, 34],很大程度上提高植物的纵向蒸腾拉力,对土壤水分吸收有益。还有大量研究表明,菌根共生在不改变总根系生物量的情况下,能影响根系分支、根直径和根系密度等[35, 36, 37],这些特点均有利于说明菌根共生对宿主植物水分吸收有贡献。
由于AMF是多核无隔膜或隔膜非常稀少的菌丝体,水分可直接通过菌丝到达丛枝,菌丝内的水分运输几乎没有阻力,到达菌丝顶端丛枝后,水分渗出到宿主根内细胞,缩短水分在根内运输路径的同时,可能提供一种特殊的吸水途径。Alexopolis等[38]和Allen[3]分别指出水分在菌根根外菌丝的运输方式(图 2)。菌丝顶端细胞壁具有弹性和亲水性,因此顶端细胞壁有时可以张开小口,使水分渗出[39]。Querejeta等用染料在干旱胁迫下研究水分的运移情况时发现,白天叶片气孔打开,水分由F向A运输;然而在夜间,由于土壤严重干旱土壤水势较低,水分由A向F运输,甚至水分能从菌丝顶端渗出到土壤的现象[40]。Bárzana等的研究也表明在充分供水和干旱胁迫下,AMF确实具有调节宿主质外体和细胞间水分运输的途径,AMF这种灵活调节宿主水分的运输能力可能会根据植物水分的需求,更灵活的响应干旱胁迫,并利用示踪染料抑制水通道蛋白活性实验,发现水分亏缺条件下,根据宿主植物水的储存和地上部分需求,AMF共生能更灵活的调节水分运输[41]。Li等人从根内球囊霉中首次克隆、鉴定两个功能水通道蛋白基因GintAQPF1和GintAQPF2,为AMF向宿主植物提供水分运输从而提高植物耐旱性提供强有力的证据[42]。AMF菌丝提供的特殊吸水功能,有别于植物本身具有的3种吸水方式,在对宿主植物响应干旱胁迫方面发挥重要作用。

|
| 图 2 菌根共生根部示意图(a)菌根外生菌丝吸水模式图(b) Fig. 2 Water transport within a hypha (a) Schematic of root mycorrhizal symbiosis (b) |
图 2表明水分在菌丝内的运输方式:A和B是宿主植物与菌丝之间水分运移,E和F是菌丝与土壤之间水分运移。这两者之间的水分运输必须穿过菌丝膜,从而限制水分运输,但是菌丝内部没有或很少有隔膜,水分能快速实现B—C—D—E之间的运移(参照1996年Alexopolis等提出的AMF吸水模型[38])
3 菌根共生对宿主根冠通讯的影响机制1985年英国的Blackman和Davies[43]对玉米盆栽分根控水实验研究表明:受到干旱胁迫的那部分根能够引发地上部分叶片的气孔关闭,但叶水势、膨压和脱落酸的含量与充分供水时没发生显著变化,据此而提出根冠通讯的理论。也就是说,气孔导度的下降不是由于水分亏缺直接引起的,而是根系首先感受水分胁迫而产生根信号,根信号传递到叶片来控制气孔行为,这种根信号被称为非水力信号(non-hydraulic root-sourced signal,nHRS),并且在长期进化过程中这种信号向拉伸或弱化的方向演变[44, 45, 46]。Graham[47],Levy[48]和Sands[49]等研究不同植物菌根在干旱胁迫下对水力导度的影响,结果发现AMF均能影响宿主的水力导度。由于AMF植物根系有利于水分吸收和利用,因此土壤要比非菌根植物干旱的快,所以根系非水力信号的产生也早于非菌根植物。
1991年Augé等在玫瑰根围接种根内球囊霉(Glomus intraradices Schench & Smith,Gi)和不接种Gi的控水分根实验研究显示,两种处理的气孔导度有显著差别,接种Gi干旱/不接种Gi供水的处理较早降低气孔导度,而两种处理叶水势和叶片含水量没有显著变化,这意味着干旱胁迫下,菌根植物对土壤水分的吸收速度较快,而较早产生非水力信号,使气孔导度下降,减少蒸腾,达到节水保水的目的[50]。干旱胁迫条件下,叶片气孔行为是最直观的表现,菌根脱水较早而首先引发非水力信号产生,通过纵向蒸腾拉力将这些化学信号通过木质部运输到叶片的保卫细胞,保卫细胞通过失去膨压而关闭气孔,从而达到非水力信号通过根冠通讯而使气孔关闭目的,来平衡宿主植物的水分利用。
根冠通讯信号主要包括脱落酸(ABA)、细胞分裂素(CTK)、生长素、木质部pH 值和钙离子(Ca2+)等[51]。ABA是感受干旱胁迫的重要非水力化学信号分子之一,是调节气孔关闭的信号分子[52]。研究表明AMF不但能调节宿主植物ABA的含量[53, 54, 55, 56],而且它本身也能产生ABA[57]。干旱胁迫下,AMF共生影响宿主植物的碳源分配,减少非必要类异戊二烯(单萜和倍半萜烯)合成,提高必要类异戊二烯(脱落酸,叶绿素和类胡萝卜素)含量[7],有利于ABA的快速合成和积累,提高宿主植物对干旱的反应速度,进而提高耐旱能力。而且Aroca等通过对野生型和 ABA 基因突变性西红柿研究,发现菌根形成和对干旱耐受性能力,均受植物 ABA 显性基因调节[6]。干旱胁迫下,AMF与宿主植物共生影响ABA控制气孔行为的机制有两种猜测:1)AMF与植物共生影响ABA在根冠的运输;2)AMF与植物共生影响叶片保卫细胞对ABA的敏感性。Ebel等研究表明,菌根共生影响木质部ABA含量[58];Goicoechea等人研究表明丛枝菌根的ABA含量低于没有AMF共生的紫花苜蓿根系[55];Estrada-Luna和Davies的研究也表明接种AMF辣椒叶片ABA含量低于未接种植物[56]。而且Duan等对豇豆的研究表明,菌根共生并不影响气孔对ABA的敏感性[59]。可见,AMF共生通过影响ABA含量和根冠运输来调节叶片气孔行为的。菌根共生植物根、木质部和叶片ABA含量均低于非菌根植物,表明AMF与植物共生对宿主非水力信号的影响可能也存在弱化的作用。
大量研究表明,植物激素水平例如细胞分裂素、生长素、生长素相关物质、脱落酸以及茉莉酸等由于与AMF共生而改变[52, 60, 61, 62, 63]。Murakami-Mizukami 等研究发现AMF可以改变宿主根系ABA和IAA的含量,影响叶片气孔行为,从而有利于宿主逃避干旱胁迫[54];ABA与细胞分裂素之间的平衡对叶片气孔的调节效果比单独ABA或细胞分裂素效果要好[55]。菌根共生增加宿主植物IAA、GA和CTK的含量,降低ABA和乙烯含量,对宿主植物平衡水分代谢应对干旱胁迫有非常重要的作用[64]。但是目前有关AMF与植物共生的信号交互作用对干旱响应机制的研究报道还不多。因此,AMF与植物共生信号的交互效应对干旱胁迫的响应是目前需要研究的热点和难点问题[65]。
AMF共生植物与非共生植物对土壤营养物质的吸收不同,AMF共生植物有利于磷和钙元素向地上部分运输。研究发现,气孔导度与向日葵木质部磷浓度和阴阳离子密切相关,而接种AMF能显著提高木质部磷元素含量[66]。叶片细胞质和质外体的钙离子浓度参与ABA对气孔的调控,AMF与植物共生影响叶片钙离子的浓度,对气孔行为也发挥一定的调节作用[67, 68]。菌根共生影响木质部pH值,可能存在其它信号物质而影响干旱条件下非水力信号的根冠通讯[69]。少量氢离子从木质部运输到叶片,却能对叶片质外体pH值有很大影响;叶片pH值升高有利于ABA向保卫细胞转移,进而调控气孔行为[70]。随着土壤干旱的加剧,叶水势下降,水力信号产生(hydraulic root sourced signal,HRS),并由nHRS和HRS共同调节叶片的水分代谢和气体交换,在整个植株水平上对干旱刺激作出生理生态响应,例如干旱胁迫下的渗透调节作用和活性氧代谢调节。大量研究表明,AMF能提高宿主植物的渗透调节能力[11, 71, 72, 73],降低活性氧对宿主植物造成的损伤[12, 13, 14, 74]。
总之,菌根共生有利于宿主对水分和营养物质的吸收。干旱胁迫下,由于菌根共生特殊的水分吸收方式,使根部较早的产生非水力信号,由于菌根植物自身的养分积累和AMF菌丝对宿主植物激素含量有调节功能,有利于根源信号的传递和宿主植物对信号的快速应答,最终提高了宿主植物对干旱的耐受性。
4 菌根共生对根冠通讯研究存在的问题及可能解决方法根冠通讯理论是植物水分关系研究领域最重要的进展之一。该理论基于水分胁迫下植物的生理生态特征变化而提出的,该理论对植物抗干旱胁迫机制研究具有重要的意义。我们将根冠通讯分子机制研究存在的难题分解成三个问题:1)根细胞如何感知水分胁迫?2)早期抗旱基因的表达与nHRS 的关系?3)晚期抗旱基因的表达与HRS的关系?研究不同水分梯度下根系分泌物和根成分差异,对揭示植物根细胞感知水分胁迫的信号组件具有潜力。ABF (ABA-binding factor)是早期响应干旱胁迫的重要转录子,而Ca2+作为第二信使,在HRS产生前,协同ABA调控保卫细胞的关闭发挥重要作用。因此,研究不同水分梯度下植物各器官ABA、Ca2+等含量与分布,有望在揭示早期抗旱基因的表达与nHRS 的关系方面有所突破。HRS产生,一系列晚期抗旱功能基因表达,如RD系列脱水响应基因、水离子通道蛋白基因和渗透调节蛋白基因等,晚期抗旱基因的表达对整个植物株型、生物量分配和细胞生理生化等性能产生影响,所以通过研究不同水分梯度下植物的系统耐旱性(指标包括株型、生物量分配等)和细胞耐旱性(抗氧化酶系和渗透物质等),可望揭示晚期抗旱基因的表达与HRS的关系。菌根共生对以上3个环节均有影响,系统展开以上3个问题的研究,以期对揭示AMF影响根冠通讯分子机制研究有所启示。
5 结语AMF与植物共生对宿主植物生理生态影响和提高植物对非生物胁迫(重金属、盐碱和干旱等)的耐受性方面已经取得很多成果。AMF与植物共生一方面通过根系分泌物团聚土壤来保留土壤水分和外延菌丝促进水分吸收,另一方面菌根共生对根源化学信号产生和传递有一定的调节作用,将干旱信息迅速传递到地上部分,使宿主植物迅速对干旱响应,减少气孔导度、降低水的蒸腾损耗,积累渗透物质、提高氧化酶活性,以合理分配和利用现有水分资源,度过干旱胁迫。正是由于菌根能更迅速的感受干旱胁迫,并积极对水分胁迫做出响应,所以AMF赋予宿主植物更强的干旱耐受性和干旱胁迫下继续生长的特性。目前,AMF与植物共生对干旱耐受性机理的研究,已经逐渐由生理生态研究,进一步转向以基因组、蛋白质组学和生理生态为基础的研究。AMF与植物共生提高宿主植物干旱耐受性的信号产生和传递机制以及生理生态响应机理研究,为现代旱地农业育种和发展开辟新的途径。
致谢: 西澳大学植物生物学院何新华博士、兰州大学生命科学学院赵旭哲博士帮助写作,特此致力。| [1] | 李涛, 赵之伟. 丛枝菌根真菌产球囊霉素研究进展. 生态学杂志, 2005, 24(9): 1080-1084. |
| [2] | Wu Q S, Xia R X, Zou Y N. Improved soil structure and citrus growth after inoculation with three arbuscular mycorrhizal fungi under drought stress. European Journal of Soil Biology, 2008, 44(1): 122-128. |
| [3] | Allen M F. Bidirectional water flows through the soil-fungal-plant mycorrhizal continuum. New Phytologist, 2009, 182(2): 290-293. |
| [4] | Frank A B. Üeber die auf Wurzelsymbiose beruhende Ernährung gewisser Baüme durch unterirdische Pilze. Berichteder Deutschen Botanischen Gesellschaft, 1985, 3: 395-409. |
| [5] | Safir G R, Boyer J S, Gerdemann J W. Mycorrhizal enhancement of water transport in soybean. Science, 1971, 172(3983): 581-583. |
| [6] | Aroca R, Ruiz-Lozano J M, Zamarreño A M, Paz J A, García-Mina J M, Pozo M J, López-Ráez J A. Arbuscular mycorrhizal symbiosis influences strigolactone production under salinity and alleviates salt stress in lettuce plants. Journal of Plant Physiology, 2013, 170(1): 47-55. |
| [7] | Asensio D, Rapparini F, Peñuelas J. AM fungi root colonization increases the production of essential isoprenoids vs. nonessential isoprenoids especially under drought stress conditions or after jasmonic acid application. Phytochemistry, 2012, 77: 149-161. |
| [8] | Navarro G A, Del Pilar Bañón Árias S, Morte A, Sánchez-Blanco M J. Effects of nursery preconditioning through mycorrhizal inoculation and drought in Arbutus unedo L. plants. Mycorrhiza, 2011, 21(1): 53-64. |
| [9] | Lambais M R, Rios-Ruiz W F, Andrade R M. Antioxidant responses in bean (Phaseolus vulgaris) roots colonized by arbuscular mycorrhizal fungi. New Phytologist, 2003, 160(2): 421-428. |
| [10] | Abbaspour H, Saeidi-Sar S, Afshari H, Abdel-Wahhab M A. Tolerance of Mycorrhiza infected pistachio (Pistacia vera L.) seedling to drought stress under glasshouse conditions. Journal of Plant Physiology, 2012, 169(7): 704-709. |
| [11] | Wu Q S, Xia R X, Zou Y N, Wang G Y. Osmotic solute responses of mycorrhizal citrus (Poncirus trifoliata) seedlings to drought stress. Acta Physiologiae Plantarum, 2007, 29(6): 543-549. |
| [12] | Wu Q S, Xia R X, Zou Y N. Reactive oxygen metabolism in mycorrhizal and non-mycorrhizal citrus (Poncirus trifoliata) seedlings subjected to water stress. Journal of Plant Physiology, 2006, 163(11): 1101-1110. |
| [13] | Yousef S, Gholamreza H, Weria W, Kazem G G, Khosro M. Changes of antioxidative enzymes, lipid peroxidation and chlorophyll content in chickpea types colonized by different Glomus species under drought stress. Symbiosis, 2012, 56(1): 5-18. |
| [14] | Alguacil M M, Hernández J A, Portillo B, Roldán A. Antioxidant enzyme activities in shoots from three mycorrhizal shrub species afforested in a degraded semi-arid soil. Physiologia Plantarum, 2003, 118(4): 562-570. |
| [15] | Zhu X C, Song F B, Liu S Q, Liu T D. Effects of arbuscular mycorrhizal fungus on photosynthesis and water status of maize under high temperature stress. Plant and Soil, 2011, 346(1/2): 189-199. |
| [16] | Levy Y, Krikun J. Effect of vesicular-arbuscular mycorrhiza on Citrus jambhiri water relations. New Phytologist, 1980, 85(1): 25-31. |
| [17] | Miri M R, Moghadam H R T, Ghooshchi F, Zahedi H. Growth, seed yield and phosphorus uptake of wheat as influenced by Azotobacter and arbuscular mycorrhizal colonization under drought stress conditions. Research on Crops, 2012, 13(1): 21-28. |
| [18] | Ruiz-Lozano J M, Gómez M, Nuñez R, Azcón R. Mycorrhizal colonization and drought stress affect Δ13C in CO2-labeled lettuce plants. Physiologia Plantarum, 2000, 109(3): 268-273. |
| [19] | Veresoglou S D, Menexes G R, Matthias C. Do arbuscular mycorrhizal fungi affect the allometric partition of host plant biomass to shoots and roots? A meta-analysis of studies from 1990 to 2010. Mycorrhiza, 2012, 22(3): 227-235. |
| [20] | Goicoechea N, Merino S, Sanchez-Diaz M. Contribution of arbuscular mycorrhizal fungi (AMF) to the adaptations exhibited by the deciduous shrub Anthyllis cytisoides L. under water deficit. Physiologia Plantarum, 2004, 122(4): 453-464. |
| [21] | Jose B, Ronco M G. Improved tolerance of wheat plants (Triticum aestivum L.) to drought stress and rewatering by the arbuscular mycorrhizal fungus Glomus claroideum: Effect on growth and cell membrane stability. Brazilian Journal of Plant Physiology, 2008, 20(1): 29-37. |
| [22] | Augé R M. Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis. Mycorrhiza, 2001, 11(1): 3-42. |
| [23] | 吴强盛. 园艺植物丛枝菌根研究与应用. 北京: 科学出版社, 2010: 85-91. |
| [24] | Sands R, Theodorou C. Water uptake by mycorrhizal roots of radiata pine seedlings. Australian Journal of Plant Physiology, 1978, 5(3): 301-309. |
| [25] | Duddridge J A, Malibari A, Read D J. Structure and function of mycorrhizal rhizomorphs with special reference to their role in water transport. Nature, 1980, 287(5785): 834-836. |
| [26] | Brownlee C, Duddridge J A, Read D J. The structure and function of mycelial systems of ectomycorrhizal roots with special reference to their role in forming inter-plant connections and providing pathways for assimilate and water transport. Plant and Soil, 1983, 71(1/3): 433-443. |
| [27] | Ignacio Q J, Egerton-Warburton L M, Prieto I, Vargas R, Allen M F. Changes in soil hyphal abundance and viability can alter the patterns of hydraulic redistribution by plant roots. Plant and Soil, 2012, 355(1/2): 63-73. |
| [28] | Tataranni G, Montanaro G, Dichio B, Xiloyannis C. Effects of mycorrhizas on hydraulic conductivity in micrografted myrobolan 29C rootstocks. Acta Horticulturae, 2012, 966: 235-240. |
| [29] | Khalvati M A, Hu Y, Mozafar A, Schmidhalter U. Quantification of water uptake by arbuscular mycorrhizal hyphae and its significance for leaf growth, water relations, and gas exchange of barley subjected to drought stress. Plant Biology, 2005, 7(6): 706-712. |
| [30] | Ruth B, Khalvati M, Schmidhalter U. Quantification of mycorrhizal water uptake via high-resolution on-line water content sensors. Plant and Soil, 2011, 342(1/2): 459-468. |
| [31] | Allen M F. Influence of vesicular-arbuscular mycorrhizae on water movement through Bouteloua gracilis (H. B. K.) Lag ex Steud. New Phytologist, 1982, 91(2): 191-196. |
| [32] | Bethlenfalvay G J, Brown M S, Franson R. Glycine-Glomus-Rhizobium symbiosis X. relationships between leaf gas exchange and plant and soil water status in nodulated, mycorrhizal soybean under drought stress. Plant Physiology, 1990, 94(2): 723-728. |
| [33] | Shrestha Y H, Ishii T, Kadoya K. Effect of vesicular-arbuscular mycorrhizal fungi on the growth, photosynthesis, transpiration and the distribution of photosynthates of bearing satsuma mandarin trees. Journal of the Japanese Society for Horticultural Science, 1995, 64(3): 517-525. |
| [34] | Fidelibus M W, Martin C A, Stutz J C. Geographic isolates of Glomus increase root growth and whole-plant transpiration of Citrus seedlings grown with high phosphorus. Mycorrhiza, 2001, 10(5): 231-236. |
| [35] | Kabouw P, van Dam N M, van der Putten W H, Arjen B. How genetic modification of roots affects rhizosphere processes and plant performance. Journal of Experimental Botany, 2012, 63(9): 3475-3483. |
| [36] | Allaway W G, Ashford A E. Structure of hair roots in Lysinema ciliatum R. Br. and its implications for their water relations. Annals of Botany, 1996, 77(4): 383-388. |
| [37] | Feil W, Kottlee I, Oberwinkler F. The effect of drought on mycorrhizal production and very fine root system development of Norway spruce under natural and experimental conditions. Plant and Soil, 1988, 108(2): 221-231. |
| [38] | Alexopoulos C J, Mims C W, Blackwell M. Introductory Mycology. 4th ed. New York: John Wiley and Sons, 1996. |
| [39] | Allen M F. Water dynamics of mycorrhizas in arid soils // Gadd G M, ed. Fungi in Biogeochemical Cycles. Cambridge: Cambridge University Press, 2006: 74-97. |
| [40] | Querejeta J I, Eggerton-Warburton L, Allen M. Direct nocturnal water transfer from oaks to their mycorrhizal symbionts during severe soil drying. Oecologia, 2003, 134(1): 55-64. |
| [41] | Bárzana G, Aroca R, Paz J A, Chaumont F, Martinez-Ballesta M C, Carvajal M, Ruiz-Lozano J M. Arbuscular mycorrhizal symbiosis increases relative apoplastic water flow in roots of the host plant under both well-watered and drought stress conditions. Annals of Botany, 2012, 109(5): 1009-1017. |
| [42] | Li T, Hu Y J, Hao Z P, Li H, Wang Y S, Chen B D. First cloning and characterization of two functional aquaporin genes from an arbuscular mycorrhizal fungus Glomus intraradices. New Phytologist, 2013, 197(2): 617-630. |
| [43] | Blackman P G, Davies W J. Root-to-shoot communication in maize plants of the effects of soil drying. Journal of Experimental Botany, 1985, 36(1): 39-48. |
| [44] | Xiong Y C, Li F M, Xu B C, Hadgkinson K C. Hydraulic and non-hydraulic root-sourced signals in old and modern spring wheat cultivars in a semiarid area. Journal of Plant Growth Regulation, 2006, 25(2): 120-136. |
| [45] | Xiong Y C, Li F M, Zhang T, Xia C. Evolution mechanism of non-hydraulic root-to-shoot signal during the anti-drought genetic breeding of spring wheat. Environmental and Experimental Botany, 2007, 59(2): 193-205. |
| [46] | Xiong Y C, Li F M, Zhang T. Performance of wheat crops with different chromosome ploidy: root-sourced signals, drought tolerance, and yield performance. Planta, 2006, 224(3): 710-718. |
| [47] | Graham J H, Syvertsten J P. Influence of vesicular-arbuscular mycorrhiza on the hydraulic conductivity of roots of two Citrus rootstocks. New Phytologist, 1984, 97(2): 277-284. |
| [48] | Levy Y, Syvertsen J P, Nemec S. Effect of drought stress and vesicular-arbusuclar mycorrhiza on Citrus transpiration and hydraulic conductivity of roots. New Phytologist, 1983, 93(1): 61-66. |
| [49] | Sands R, Fiscus E L, Reid C P P. Hydraulic properties of pine and bean roots with varying degrees of suberization, vascular differentiation and mycorrhizal infection. Australian Journal of Plant Physiology, 1982, 9(5): 559-569. |
| [50] | Augé R M, Duan X. Mycorrhizal fungi and nonhydraulic root signals of soil drying. Plant Physiology, 1991, 97(2): 821-824. |
| [51] | 李冀南, 李朴芳, 孔海燕, 熊俊兰, 王绍明, 熊友才. 干旱胁迫下植物根源化学信号研究进展. 生态学报, 2011, 31(9): 2610-2620. |
| [52] | Davies W J, Tardieu F, Trejo C L. How do chemical signals work in plants that grow in dry soil? Plant Physiology, 1994, 104(2): 309-314. |
| [53] | Allen M F, Moore T S, Christensen M. Phytohormone changes in Bouteloua gracilisinfected by vesicular-arbuscular mycorrhizae. II. Altered levels of gibberellin-like substances and abscisic acid in the host plant. Canadian Journal of Botany, 1982, 60(4): 468-471. |
| [54] | Murakami-Mizukami Y, Yamamoto Y, Yamaki S. Analyses of indole acetic acid and abscisic acid content in nodules of soybean plants bearing VA mycorrhizas. Soil Science and Plant Nutrition, 1991, 37(2): 291-298. |
| [55] | Goicoechea N, Antolin M C, Sánchez-Díaz M. Gas exchange is related to the hormone balance in mycorrhizal or nitrogen-fixing alfalfa subjected to drought. Physiologia Plantarum, 1997, 100(4): 989-997. |
| [56] | Estrada-Luna A A, Davies F T. Arbuscular mycorrhizal fungi influence water relations, gas exchange, abscisic acid and growth of micropropagated chile ancho pepper (Capsicum annuum) plantlets during acclimatization and post-acclimatization. Journal of Plant Physiology, 2003, 160(9): 1073-1083. |
| [57] | Esch H, Hundeshagen B, Schneiderpoetsch H, Bothe H. Demonstration of abscisic acid in spores and hyphae of the arbuscular-mycorrhizal fungus Glomus and in the N2-fixing cyanobacterium Anabaena variabilis. Plant Science, 1994, 99(1): 9-16. |
| [58] | Ebel R C, Duan X, Still D W, Augé R M. Xylem sap abscisic acid concentration and stomatal conductance of mycorrhizal Vigna unguiculata in drying soil. New Phytologist, 1997, 135(4): 755-761. |
| [59] | Duan X, Neuman D S, Reiber J M, Green C D, Saxton A M, Augé R M. Mycorrhizal influence on hydraulic and hormonal factors implicated in the control of stomatal conductance during drought. Journal of Experimental Botany, 1996, 47(10): 1541-1550. |
| [60] | Hause B, Mrosk C, Isayenkov S, Strack D. Jasmonates in arbuscular mycorrhizal interactions. Phytochemistry, 2007, 68(1): 101-110. |
| [61] | Edriss M H, Davis R M, Burger D W. Influence of mycorrhizal fungi on cytokinin production in sour orange. Journal of the American Society for Horticultural Science, 1984, 109: 587-590. |
| [62] | Drüge U, Schönbeck F. Effect of vesicular-arbuscular mycorrhizal infection on transpiration, photosynthesis and growth of flax (Linum usitatissimum L.) in relation to cytokinin levels. Journal of Plant Physiology, 1992, 141(1): 40-48. |
| [63] | Thiagarajan T R, Ahmad M H. Phosphatase activity and cytokinin content in cowpeas (Vigna unguiculata) inoculated with a vesicular-arbuscular mycorrhizal fungus. Biology and Fertility of Soils, 1994, 17(1): 51-56. |
| [64] | Cruz A F, Ishii T, Kadoya K. Effects of arbuscular mycorrhizal fungi on tree growth, leaf water potential, and levels of 1-aminocyclopropane-1-carboxylic acid and ethylene in the roots of papaya under water-stress conditions. Mycorrhiza, 2000, 10(3): 121-123. |
| [65] | Thomas F, Bettina H. Drought and symbiosis-why is abscisic acid necessary for arbuscular mycorrhiza? New Phytologist, 2007, 175(3): 383-386. |
| [66] | Gollan T, Schurr U, Schulze E D. Stomatal response to drying soil in relation to changes in the xylem sap composition of Helianthus annuus. I. The concentration of cations, anions, amino acids in, and pH of, the xylem sap. Plant, Cell and Environment, 1992, 15(5): 551-559. |
| [67] | Thompson D S, Wilkinson S, Bacon M A, William J D. Multiple signals and mechanisms that regulate leaf growth and stomatal behaviour during water deficit. Physiologia Plantarum, 1997, 100(2): 303-313. |
| [68] | De Silva D L R, Cox R C, Hetherington A M, Mansfield T A. The role of abscisic acid and calcium in determining the behaviour of adaxial and abaxial stomata. New Phytologist, 1986, 104(1): 41-51. |
| [69] | Wilkinson S, Davies W J. Xylem sap pH increase: a drought signal received at the apoplastic face of the guard cell that involves the suppression of saturable abscisic acid uptake by the epidermal symplast. Plant Physiology, 1997, 113(2): 559-573. |
| [70] | Hartung W, Wilkinson S, Davies W J. Factors that regulate abscisic acid concentrations at the primary site of action at the guard cell. Journal of Experimental Botany, 1998, 49: 361-367. |
| [71] | Augé R M, Schekel K A, Wample R L. Osmotic adjustment in leaves of VA mycorrhizal and nonmycorrhizal rose plants in response to drought stress. American Society of Plant Biologists, 1986, 82(3): 765-770. |
| [72] | Osonubi O. Osmotic adjustment in mycorrhizal Gmelina arborea Roxb. seedlings. Functional Ecology, 1989, 3(2): 143-151. |
| [73] | Kubikova E, Moore J L, Ownley B H, Mullen M D, Augé R M. Mycorrhizal impact on osmotic adjustment in Ocimum basilicum during a lethal drying episode. Journal of Plant Physiology, 2001, 158(9): 1227-1230. |
| [74] | Lee B R, Muneer S, Jung W J, Avice J C, Ourry A, Kim T H. Mycorrhizal colonization alleviates drought-induced oxidative damage and lignification in the leaves of drought-stressed perennial ryegrass (Lolium perenne). Physiologia Plantarum, 2012, 145(3): 440-449. |