 2015, Vol. 35
2015, Vol. 35文章信息
- 曾娟
- ZENG Juan
- 我国草地螟轻发年份时空特征及其气候背景
- Temporal and spatial patterns of meadow moth (Loxostege sticticalis L.) in mild occurrence years in china and the associated climatic background
- 生态学报, 2015, 35(6): 1899-1909
- Acta Ecologica Sinica, 2015, 35(6): 1899-1909
- http://dx.doi.org/10.5846/stxb201305090993
-
文章历史
- 收稿日期:2013-10-14
- 修订日期:2014-04-05






草地螟Loxostege sticticalis L.是广泛分布于欧亚大陆干旱、半干旱地区的世界性害虫,在暴发年份对农牧业生产造成巨大影响和严重威胁;在与气候、生态等环境条件协同进化的过程中,草地螟具备了远距离迁飞、滞育、产卵选择和间歇性暴发等生存对策和环境适应机制,具有种群时空波动的典型现象。在国外,草地螟的研究历史由来已久,前苏联地区从 19 世纪中期至20世纪70年代一直有草地螟大发生的记载[1];20世纪70年代以后,关于草地螟在西西伯利亚、西伯利亚和远东地区、哈萨克斯坦以及俄罗斯欧洲部分和亚洲部分的大发生记载也在持续[2, 3, 4, 5, 6, 7]。在我国,新中国成立以来,草地螟已经出现了3个暴发为害周期[8]。总体看来,无论从草地螟年度间发生的历史变化规律来讲,还是从年度内发生的时空分布动态来讲,不同暴发周期之间、同一个暴发周期内的不同年份之间,草地螟发生期早晚、持续时间长短、重发区域分布和最终为害程度均存在显著差异,且这种差异不仅体现在大发生年份与轻发年份之间,也体现在总体轻发的不同年份之间。然而,对于暴发周期的研究往往集中分析大发生年份的发生特点、虫源推演和环境促使因素[9, 10, 11, 12, 13],对轻发年份鲜有关注;而对于轻发年份的记载也往往以单一年度为对象[14, 15, 16, 17],缺乏更为深入的系统性、对比性研究。与重发年份一样,轻发年份同样是构成草地螟长周期种群波动不可或缺的环节,因此很有必要对草地螟种群数量低谷期的时空分布特征和主导因素进行研究,这样有助于对草地螟发生的时空变化规律形成普遍性结论和宏观性判断。
本文以草地螟轻发年份为研究对象,总结轻发特点的共同性,比较时空分布的多样性,分析气象条件的大尺度变化与草地螟发生轻重的相关性,对掌握草地螟发生危害的年度差异、阐明气候变化对其发生趋势的持续影响,提高监测预警水平具有重要意义。
1 资料与方法 1.1 我国草地螟1949—2012年度发生面积波动分析草地螟虫情资料系全国农业技术推广服务中心收集整理。1949—2012年全国草地螟(幼虫)年度发生面积资料分为三个阶段,其中1949—1978年数据引自《中国植物保护五十年》[18],1979—1995年数据引自《全国植保专业统计资料》,以上两个时间段内全国草地螟年度发生面积由“大豆草地螟”、“其他粮作草地螟”和“其他经作草地螟”发生面积合计得出;1996—2012年数据由北方农牧区各省草地螟年度发生统计表中各代次(幼虫)在农田、林地、草场的发生面积数据合计得出。
根据我国草地螟发生面积数据三个历史阶段统计方法、数据涵盖内容和资料整齐度的差异,且三个暴发周期分别处于三个不同的历史阶段,因此按1953—1959年、1979—1985年、1996—2012年3个暴发周期分别进行年度发生面积的时间序列分析。在同一个暴发周期内,假定其为遵从正态分布的随机变量,计算年度发生面积序列的均值μ和标准误σ,以明确序列内的总体发生水平和离散程度,并以(μ±σ)作为得出同一暴发周期内重发生、轻发生年份的阈值划分标准:发生面积大于(μ+σ)的,计为重发生;发生面积小于(μ-σ)的,计为轻发生;发生面积在(μ-σ)与(μ+σ)之间的,计为一般发生。
1.2 我国草地螟2006—2012年时空分布特征分析2006—2012年草地螟年度时空分布特征资料,包括我国北方主要发生省份每年各代次各虫态发生期出现时间、持续时间和发生县点数。同一年份内的同一项数据,按照草地螟发生主要区域进行细分,具体分为内蒙古中西部(自巴彦淖尔至锡林郭勒盟)、内蒙古东南部(通辽市、赤峰市)、内蒙古东北部(兴安盟、呼伦贝尔市)、河北、山西、北京、天津、辽宁、吉林、黑龙江、陕西、宁夏、新疆北疆、新疆南疆共计14个区域。
1.2.1 发生期早晚每一年份内,分别收集14个草地螟发生区域的越冬代成虫、一代幼虫、一代成虫、二代幼虫的始见期或盛期,按旬统计同一虫态在各发生区域出现时间分布频次,如2006年内蒙古中西部越冬代成虫始见期为5月14日,则将其归入5月中旬并计频次。在Excel中作2006—2012年各虫态发生期出现早晚频次的面积堆积图。
1.2.2 盛期持续时间每一年份内,分别收集14个草地螟发生区域的越冬代成虫、一代幼虫、一代成虫、二代幼虫的发生盛期,并乘以各发生区域内的实际见虫县点数,得出该区域内该虫态发生的有效持续时间。利用DPS9.50软件,对2006—2012年各虫态发生盛期的有效持续时间进行方差分析(显著性差异0.05)。
1.2.3 发生区域每一年份内,分别收集14个草地螟发生区域的越冬代成虫、一代幼虫、一代成虫、二代幼虫的发生县点数。在Excel中作2006—2012年各虫态发生区域数的柱状图。
1.3 大尺度气象条件影响分析 1.3.1 气象数据处理气象资料来自中国气象数据共享服务网。参照霍治国等[19]分析长时间序列上气象条件与虫害发生面积相关性方法,将全国564个气象站点与我国北方草地螟发生区域(2006—2012年草地螟发生县级点)进行比对,筛选出169个草地螟发生区气象站点,采用各气象站点1951—2012年大尺度气象资料(1949—1950年所有站点气象数据缺测),包括年平均气温、1月份平均气温和6—8月份累计降水量。
设x因子第i个站点第j年表示为 xij(i=1,2,…,169;j=1,2,…,61),则草地螟发生区第j年x因子的年度平均值为:
1.3.2 相关性分析
在Excel中,结合草地螟年度发生面积,对1951—2012年草地螟发生区各气象因子的年度平均值进行时间序列分析,作出各气象因子和草地螟发生面积年度变化趋势线,并进行比较。利用DPS9.50软件,对各气象因子与草地螟年度发生面积的相关性进行分析,计算得出线性方程及相关指数。
2 结果与分析 2.1 草地螟轻发年份特点 2.1.1 轻发年份频次多于重发年份1949—2012年,我国草地螟发生面积的年度变化呈现明显的周期性,能引起农作物受害的年份集中于3个暴发周期,其余年份为间歇期(图 1)。同一个暴发周期内,根据不同年份发生面积与平均值和标准误之间关系,区分重发生年(图 1中实心五角星标注)、轻发生年(图 1中空心五角星标注)和一般发生年,结果表明,3个暴发周期内轻发生年份出现频次多于重发生年份。如,第一暴发周期内(1953—1959年),3a为轻发生,2a为重发生,2a为一般发生;第二暴发周期内(1979—1985年),3a为轻发生,1a为重发生,3a为一般发生;第三暴发周期内(1996—2012年),9a为轻发生,5a为重发生,3a为一般发生。总体来看,暴发周期内,轻发生年份出现比率为48.4%,重发生年份出现比率为25.8%,一般年份出现比率为25.8%,即在暴发周期中轻发生年份占多数。

|
| 图 1 我国1949—2012年草地螟发生面积及轻重年份对比 Fig. 1 Comparison of meadow moth occurrence area during 1949—2012 in China |
草地螟间歇期发生面积更能体现“轻发”的普遍性。处于间歇期的年份共有33a,占整个统计周期(64a)的51.6%。间歇期内,发生面积为0的年份有18a,占间歇期的54.5%;发生面积为0—1 万hm2的年份有4a,发生面积为1—3 万hm2的年份有8a,二者合计占间歇期的36.4%;其余4a发生面积为5—20 万hm2。
2.1.2 轻发年份发生期迟2006—2012年,我国北方每年度各代次各虫态的发生期频次分布(14个草地螟发生区域累计值),总体特点为重发年份发生期偏向提前分布、轻发年份发生期偏向推迟分布,即重发年份发生期偏早、轻发年份发生期偏迟(图 2)。7a中,各代次各虫态一般出现时期(分布频次超过50%以上)及频次比率、提前时期和推迟时期中重发年份和轻发年份的分布频次具体如下:

|
| 图 2 我国草地螟2006—2012年各世代各虫态发生时期频次 Fig. 2 Occurrence frequency of meadow moth in each stage and generation during different periods among 2006—2012 in China |
越冬代成虫始见期 一般出现在4月下旬至5月中旬,出现频次占66.7%;提前出现时期为4月上旬至中旬,其中,重发年份(2008、2009年)出现4次,轻发年份(2010、2011年)出现3次;推迟出现时期为5月下旬至6月中旬,其中重发年份(2008、2009年)出现4次,轻发年份(2006、2010、2011、2012年)出现9次。
越冬代成虫盛期 一般出现在5月下旬至6月中旬,出现频次占56.0%;提前出现时期为4月下旬至5月中旬,其中,重发年份(2009年)出现8次,轻发年份(2011、2012年)出现2次;推迟出现时期为6月中旬至下旬,其中重发年份未出现,轻发年份(2006、2010、2011、2012年)出现8次。
一代幼虫盛期 一般出现在6月中旬至下旬,出现频次占71.1%;提前出现时期为6月上旬,其中,重发年份(2008、2009年)出现4次,轻发年份(2010、2011、2012年)出现6次;推迟出现时期为7月上旬至中旬,其中重发年份未出现,轻发年份(2006、2012年)出现2次。
一代成虫盛期 一般出现在7月上旬至8月上旬,出现频次占67.7%;提前出现时期为6月下旬,其中,重发年份(2009年)出现1次,轻发年份(2010年)出现5次;推迟出现时期为8月中旬至下旬,其中重发年份(2008年)出现1次,轻发年份(2006年)出现2次。
二代幼虫盛期 一般出现在7月下旬至8月中旬,出现频次占63.0%;提前出现时期为7月中旬,其中,重发年份(2009年)出现2次,轻发年份(2010年)出现1次;推迟出现时期为8月下旬至9月中旬,其中重发年份(2008年)出现1次,轻发年份(2006、2010、2012年)出现6次。
2.1.3 轻发年份发生盛期短我国北方每年度各代次各虫态发生盛期有效持续时间(14个草地螟发生区域平均值),2006—2012年的不同年份之间存在显著性差异,且这种差异集中体现了重发年份与轻发年份的不同(图 3)。具体表现为:

|
| 图 3 我国草地螟2006—2012年各世代各虫态发生盛期有效持续时间 Fig. 3 Effectively continuing period of meadow moth in each stage and generation among 2006—2012 in China |
越冬代成虫盛期有效持续时间,2009年为299.8 d,2007年为109.6 d,2008年为53.4 d,其余年份为50 d以下;在P≤5%的显著性水平进行差异分析,2009年(重发年份)显著多于其他年份,其他年份彼此之间差异不显著。
一代幼虫盛期有效持续时间,2009年为223.4 d,2007年为71.4 d,2008年为60.0 d,其余年份为50 d以下;在P≤5%的显著性水平进行差异分析,2009年(重发年份)显著多于2006、2010、2011年(均为轻发年份),2007年(一般发生年份)、2008年(重发年份)和2012年(轻发年份)介于以上二种差异之间。
一代成虫盛期有效持续时间,2008年为229.6 d,其余年份为40 d以下;在P≤5%的显著性水平进行差异分析,2008年(重发年份)显著多于其他年份,其他年份之间差异不显著。
二代幼虫盛期有效持续时间,2008年为320.7 d,其余年份为20 d以下;在P≤5%的显著性水平进行差异分析,2008年(重发年份)显著多于其他年份,其他年份之间差异不显著。
2.1.4 轻发年份发生区域少我国北方每年度各代次各虫态发生区域涉及县点数,2006—2012年的不同年份之间存在明显差异,且这种差异集中体现了重发年份与轻发年份的不同(图 4)。具体表现为:

|
| 图 4 我国草地螟2006—2012年各世代各虫态发生区域数(县级) Fig. 4 Occurrence regions(counties)of meadow moth in each stage and generation among 2006—2012 in China |
越冬代成虫发生区域涉及县点数,2009年为261个,2007年为110个,2008年为97个,2010—2012年为70—80个,2006年为34个;一代幼虫发生区域涉及县点数,2009年为152个,2008年为82个,2007年为79个,2006年及2010—2012年为30—40个。7a间,越冬代成虫和一代幼虫的发生区域数差异特点表现一致,即,2009年(重发年份)明显偏多,轻发年份(2006年、2010—2012年)明显偏少,2008年(重发年份)和2007年(一般发生年份)居中。
一代成虫发生区域涉及县点数,2008年为352个,2009年为88个,2007年及2010—2012年为40—60个,2006年为4个;二代幼虫发生区域涉及县点数,2008年为252个,2009年为35个,2007年及2010—2012年为15—25个,2006年为1个。7a间,一代成虫和二代幼虫的发生区域数差异特点表现一致,即,2008年(重发年份)明显偏多,轻发年份及一般发生年份(2006年、2010—2012年及2007年)明显偏少,2009年(重发年份)居中。
2.2 大尺度气象条件变化趋势与草地螟年度发生动态的相关性 2.2.1 年平均气温1951—2012年,我国北方草地螟发生区年平均气温(169点平均值)与草地螟年度发生面积的时间序列走势一致(图 5)。具体表现为:1)在62a的时间序列上,年平均气温与年度发生面积总体呈递增趋势,且趋势线斜率近似。2)在草地螟第二、三暴发周期内,年平均气温与年度发生面积的走势高度一致。如第二暴发周期内,年平均气温与年度发生面积两条曲线都出现2个峰,峰型形似;最大峰值同时出现在1982年;在暴发周期开始和结束的年份,两条曲线同时回落到趋势线平均值以下。又如第三暴发周期内,年平均气温与年度发生面积两条曲线都出现5个峰,峰型相似;尽管二者峰值出现年份不一致,但重发年份(1999、2002、2004、2008、2009年)对应的年平均气温总在各波动区间的峰值位置或次峰位置,而轻发年份(1996、2000、2005、2006、2010—2012)对应的年平均气温总在各波动区间的谷值位置或次谷位置;在暴发周期开始和结束的年份,两条曲线同时回落到趋势线平均值以下。3)在草地螟发生间歇期和第一暴发周期,由于发生面积总体水平较低,无法比较其与年平均气温的趋势关系。相关性分析表明,年平均气温与草地螟发生面积之间呈正相关,其线性方程为:


|
| 图 5 草地螟发生面积和年平均气温时间序列 Fig. 5 Time series of meadow moth occurrence area and annual average temperature |
式中,y为草地螟年度发生面积,ta为年平均气温。
2.2.2 1月月平均气温1951—2012年,我国北方草地螟发生区1月份月平均气温(169点平均值)与草地螟年度发生面积的时间序列走势吻合度不高(图 6)。具体表现为:1)在62a的时间序列上,1月月平均气温趋势线较为平缓、增加趋势并不明显,与草地螟年发生面积总体增加的趋势吻合度不高。2)在草地螟第二、三暴发周期内,1月月平均气温与年度发生面积的曲线走势总体一致,但二条曲线在部分波动区间存在“峰谷”错位的现象。如第二暴发周期内,1月月平均气温与年度发生面积两条曲线都出现2个峰,峰型形似;重发年份(1982年)对应的1月月平均气温处于第2个峰的次峰位置,一般发生年份(1980年,处于发生面积曲线的第1个峰值)对应的1月月平均气温处于谷值。又如第三暴发周期内,1月月平均气温与年度发生面积两条曲线都出现5个峰,峰型相似;1999、2002、2004年等重发年份对应的1月月平均气温处于各波动区间的峰值位置,但重发年份2008年对应的1月月平均气温处于该波动区间的谷值位置。3)在草地螟发生间歇期和第一暴发周期,由于发生面积总体水平较低,无法比较其与1月月平均气温的趋势关系。

|
| 图 6 草地螟发生面积和1月月平均气温时间序列 Fig. 6 Time series of meadow moth occurrence area and average temperature of January |
相关性分析表明,1月月平均气温与草地螟发生面积之间呈正相关,其线性方程为:
式中,y为草地螟年度发生面积,t1为1月月平均气温。 2.2.3 6—8月累计降雨量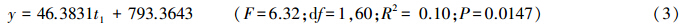
1951—2012年,我国北方草地螟发生区6—8月累计降雨量(169点平均值)与草地螟年度发生面积的时间序列走势相反(图 7)。1)在62a的时间序列上,我国北方草地螟发生区各点平均6—8月累计降雨量总体呈递减趋势,与年度发生面积的递增趋势相反。2)在草地螟第二、三暴发周期内,6—8月累计降雨量与年度发生面积的走势相反,尤其是在峰谷位置上大部分相反。如第二暴发周期内,处于发生面积曲线峰值的1982年(重发年份)和1980年(一般发生年份)对应的6—8月累计降雨量处于各波动区间的谷值;而轻发年份(1981、1984、1985年)对应的6—8月累计降雨量处于各波动区间的峰值或次峰值。又如第三暴发周期内,1999年(重发年份)对应的6—8月累计降雨量处于该波动区间的谷值,其他重发年份(2002、2004、2008、2009年)对应的6—8月累计降雨量处于各波动区间的次峰值,而轻发年份(1996、1998、2003、2005、2006、2010、2011、2012年)对应的6—8月累计降雨量都处于各波动区间的峰值或次峰值。3)在草地螟发生间歇期和第一暴发周期,由于发生面积总体水平较低,无法比较其与6—8月累计降雨量的趋势关系。

|
| 图 7 草地螟发生面积和6—8月累计降雨量时间序列 Fig. 7 Time series of meadow moth occurrence area and accumulated precipation during 6—8 month |
相关性分析表明,6—8月累计降雨量与草地螟发生面积之间呈负相关,其线性方程为:
式中,y为草地螟年度发生面积,p为6—8月累计降水量。 3 结论与讨论 3.1 轻发年份在草地螟种群动态监测中的意义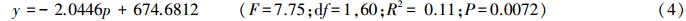
草地螟是一种大范围迁飞性害虫,其种群主要是在农牧交错区进行繁殖,只有当种群增大到一定程度时,才会大量迁入农田为害农作物[9]。自建国以来,我国对草地螟幼虫为害农作物面积有了连续记载,记为农区发生面积。因此,本文分析草地螟发生面积的有无或多少,反映了草地螟种群增大到足以引起农作物危害及其危害轻重的情况,也从一个侧面反映了我国北方生态系统中草地螟种群的消长动态。结果表明,无论是在暴发周期内,还是间歇期内,轻发年份总是占大多数。这意味着,就我国草地螟种群动态的年际变化而言,在小范围内保存和延续种群、不进入或少进入农区是常态,大范围暴发、引起农作物严重受害是少数现象。从病虫害监测预警的角度而言,持续监测“常态”,才有可能把握草地螟种群动态变化的长期规律,从而加强对突发暴发现象的预测预报能力。
3.2 草地螟发生轻重的判断指标以年度(幼虫)发生面积为主要指标进行发生轻重的总体判断,与年度内各区域、各代次、各虫态的具体指标(包括发生期出现早晚、发生盛期有效持续时间、发生区域多少)分析结果表现一致。总体趋势为轻发年份发生期迟、发生盛期短、发生区域少,与之相对应的是,重发年份发生期早、发生盛期长、发生区域多;且轻发年份与重发年份在这些具体指标上存在显著性差异。
以年度(幼虫)发生面积为指标的总体判断适用于长时间序列中不同暴发周期、不同年份之间的比较,具有一目了然的效果。结果表明,暴发周期与间歇期之间、不同暴发周期之间以及同一暴发周期的不同年份间,草地螟发生面积存在轻、重发生的明显差异,总体发生趋势与前人分析结果一致[8, 9]。同时,本文得出的草地螟发生面积长期序列也与我国草地螟长期发生存在周期性节律的结果一致[20],结合其太阳黑子活动周期的预测、且基于2010—2012年轻发年份连续出现的事实,可以判断我国草地螟第三个暴发周期结束、并已进入间歇期。
年度内各代次各虫态发生期、盛期持续时间和发生区域数等具体指标,除了可以印证轻、重年份的总体趋势,也可以用于分析同一年份内不同代次和虫态发生比重的差异。如,通过发生盛期有效持续时间和发生区域数比较,可以看出,同样为重发年份,2008年是二代幼虫重发,具体表现为一代成虫和二代幼虫盛期有效持续时间及发生区域数明显高于其他年份;2009年是一代幼虫重发,具体表现为越冬代成虫和一代幼虫有效持续时间及发生区域数明显高于其他年份;而2008年的越冬代成虫和一代幼虫、2009年的一代成虫和二代幼虫的对应指标与轻发年份或一般发生年份间都没有显著差异。以发生面积、发生期、盛期持续时间、发生区域数等量化指标进行时空分布特征的具体分析,并得出轻、重发生年份之间存在的差异,可作为探究草地螟年度发生时空变化规律的有力支撑。
前人关于草地螟种群动态的研究结果,往往集中于某一年份,多以成虫灯下虫量、百步惊蛾量、幼虫密度等为指标,分析种群数量动态和分布规律[9, 10, 11, 12, 13, 14, 15, 16, 17, 21];而本文由于在长时间序列上、全国范围内进行数据分析的复杂性,没有采用以上指标。为提高种群动态分析的准确性,同时也适应长时间、大范围分析的需要,今后可进一步细化指标,如使用虫量达到一定密度的发生面积、发生区域数等,或更能体现草地螟种群动态的变化规律。
3.3 大尺度气象条件变化引起草地螟年度发生波动关于草地螟发生年度差异与气候环境变化的关系,主要有两方面的典型事例:一是在大发生年份总是对应出现特别适宜草地螟种群突增和生长发育的环境条件[9, 11, 12, 13],如适宜越冬、羽化、产卵和幼虫发生的温湿度条件、蜜源、产卵环境和可随时取食的寄主,适合大规模种群迁入定殖的高空风场和沉降气流等;二是在轻发年份总是对应出现不适宜的气象和环境条件[14, 15, 16, 17],如夏季大范围的高温干旱、蜜源植被缺乏、生态环境恶化,成虫羽化产卵和幼虫孵化等关键发育期与气候环境条件适宜时期不配合等。关于草地螟种群数量与气象条件相关性的数值研究,主要是在大发生年份、具备大量虫源的基础上,分析温、湿度条件的影响[21];但在单个发生区(河北康保),草地螟发生面积和成、幼虫种群波动的长期序列与温湿度、降雨量等气象因子之间的相关性不明确或不稳定[22]。由于草地螟具有适生范围广、迁飞能力强、间歇性暴发等特征,同时其滞育、羽化、迁飞、交配、产卵等行为受外界环境条件、特别是受气象条件的影响十分复杂[23, 24, 25, 26],因此,试图用一时一地、或者短时间局部区域的气象条件作用机制来说明长时间、大范围内的影响规律,是不切实际的。本文选取我国北方草地螟发生区的年平均气温、1月月平均气温、6—8月累计降雨量(169个气象站点平均值)作为大尺度气象条件指标,分析其变化趋势与草地螟年际发生趋势的相关性。结果表明,以上3个气象因子与草地螟年度发生面积在62a的时间序列上存在显著线性相关。
我国草地螟发生区年平均气温呈逐渐增加的趋势,与全球变暖的趋势吻合,也与草地螟暴发周期的持续时间、暴发范围逐步增长的趋势吻合(P<0.0001),但1月份月平均气温的逐年增加趋势及其与草地螟年度发生波动的相关性并不十分显著(P=0.0147)。究其原因,可能是由于草地螟种群扩大是建立在逐年积累的基础上,气候变暖使得草地螟越冬场所扩大、越冬基数增加,导致其种群数量年度动态呈现增长趋势;但由于草地螟越冬幼虫耐寒力很强,发生区域大部各年度1月月平均气温并未超过其平均过冷却点[27],1月月平均气温并未成为草地螟越冬的限制性因素,因此该因素与草地螟年度发生动态的相关性低于年平均气温。在草地螟发生间歇期,年平均气温波动上升的趋势仍然持续,因此,年平均气温升高或者说气候变暖,并不能解释草地螟暴发的周期性节律;但由于年平均气温在总体趋势及第二、三暴发周期内与草地螟年度发生面积波动的一致性,推测其应该是在草地螟种群积累达到一定程度、促成暴发的主要影响因素之一。
另外,我国草地螟发生区6—8月累计降雨量呈总体减少的趋势,且在第二、三暴发周期内,其变化动态与草地螟年度发生动态呈现负相关(P=0.0072),这与一般认为的夏季多雨有利于草地螟发生的观点相悖。笔者认为可以从两方面来分析。一是气象条件影响的尺度不同。从单因素和草地螟发育关键期的角度看,夏季多雨有利于提高环境相对湿度、植被覆盖度、蜜源植物花期盛开程度、产卵寄主幼嫩程度和取食寄主营养丰富度,并可伴随有利于草地螟迁入的沉降气流,因而有利于草地螟的发生;然而,本文得出我国草地螟发生区6—8月份累计降雨量总体减少的趋势,是以62a的时间序列为准线、以其对草地螟种群动态的长期影响为基础的,两种分析采用的气象条件影响尺度不同,因此有可能得出不同的结论。二是该结果提供了一个分析草地螟种群动态变化的新角度。正是由于降雨总体呈减少趋势,导致我国北方土壤湿度减少[28]、北方农牧交错带地表湿润指数下降、植被覆盖度降低[29],使我国北方干旱、半干旱区域增加,草地螟适生范围扩大,同时干旱、脆弱的北方生态系统对草地螟种群的承载和容纳能力下降,进一步加大了草地螟向农区转移并发生为害的几率。
致谢: 感谢内蒙古、河北、山西、吉林、辽宁、黑龙江、北京、天津、陕西、宁夏、新疆等省(区)植保站及各监测点测报技术人员提供大量详实的草地螟发生情况数据。| [1] | Трибель С А. Тактика борьбы с луговым мотыльком. Защита Растений, 1978, 7: 31-32. |
| [2] | Endakov E. On the fields of the Altai. Zashchita Rastenii, 1979, (4): 6-7. |
| [3] | Poplavskii V V. The serious danger presented by the meadow moth. Zashchita Rastenii, 1984, (5): 43-43. |
| [4] | ШуровемковЮВ, Алехим В Т. Луговой мотылек в Восточной Сибири и на Дальнем востоке. Защита Растений, 1984, 2: 40-41. |
| [5] | Knor I B. The problem of outbreaks of the meadow moth in Siberia. Trudy Vsesoyuznogo Entomologicheskogo Obshchestva, 1986, 68: 162-165. |
| [6] | Кузнецова Т Л, Фролов А Н, Смирнова М П. Луговой мотылек активизирутся. Защита И Карантин Растений, 2001, 6: 20-21. |
| [7] | Фролов А Н, Кузнецова Т Л, Малыш Ю М, Смирнова М П. Луговой мотылек: Что показал анализ ситуации. Защита И Карантин Растений, 2005, 5: 37-40. |
| [8] | Luo L Z, Li G B, Cao Y Z. The third outbreak period of meadow moth in China is coming. Plant Protection, 1996, 22(5): 50-51. |
| [9] | Qu X F, Zhao Z R, Wang J Q. Analysis of periodic outbreak of meadow moth in agricultural and pastoral area of North China. Entomological Knowledge, 1999, 36(1): 11-14. |
| [10] | Chen X, Chen J G, Xue Y, Hao L P, Zhang Y, Zhao K J. Immigration of the 1999 outbreak populations of the meadow moth, Loxostege sticticalis L. (Lepidoptera: Pyralidae) into the northeastern part of China. Acta Entomologica Sinica, 2004, 47(5): 599-606. |
| [11] | 屈西峰, 夏冰. 2003年中国草地螟的发生特点和成功治理经验. 中国植保导刊, 2004, 24(1): 22-24. |
| [12] | 罗礼智, 屈西锋. 我国草地螟2004年危害特点及2005年一代危害趋势分析. 植物保护, 2005, 31(3): 69-71. |
| [13] | 罗礼智, 黄绍哲, 江幸福, 张蕾. 我国2008年草地螟大发生特征及成因分析. 植物保护, 2009, 35(1): 27-33. |
| [14] | 曾娟, 姜玉英, 张野. 2009年我国草地螟发生特点及原因分析. 中国植保导刊, 2010, 30(5): 33-36. |
| [15] | 曾娟, 姜玉英. 2010年中国草地螟轻发年份的新特点与成因探讨. 中国农学通报, 2011, 27(18): 273-281. |
| [16] | 曾娟, 姜玉英. 2011年我国草地螟轻发生原因分析. 中国植保导刊, 2012, 32(8): 34-38. |
| [17] | 曾娟, 姜玉英. 我国2012年草地螟发生特点与原因分析. 植物保护, 2014, 40(1): 142-148. |
| [18] | 陈生斗, 胡伯海. 中国植物保护五十年. 北京: 中国农业出版社, 2003: 36-37. |
| [19] | 张蕾, 霍治国, 王丽, 姜玉英. 气候变化对中国农作物虫害发生的影响. 生态学杂志, 2012, 31(6): 1499-1507. |
| [20] | 黄绍哲, 江幸福, 雷朝亮, 罗礼智. 草地螟 (Loxostege sticticalis) 周期性大发生与太阳黑子活动的相关性. 生态学报, 2008, 28(10): 4823-4829. |
| [21] | 黄绍哲, 罗礼智, 姜玉英, 唐继洪, 张蕾. 我国2008年草地螟大发生种群空间分布特征. 植物保护, 2011, 37(4): 76-81. |
| [22] | 黄绍哲. 我国草地螟(Loxostege sticticalis)种群时空动态规律研究 [D]. 北京: 中国农业科学院, 2010. |
| [23] | 陈瑞鹿, 王素云, 暴祥致, 孙雅杰. 草地螟滞育的研究: 光照周期、气温与发育及滞育的关系. 植物保护学报, 1957, 14(4): 253-255. |
| [24] | 程云霞. 草地螟(Loxostege sticticalis)迁飞与生殖行为的调控及互作关系 [D]. 北京: 中国农业科学院, 2012. |
| [25] | 王凯. 草地螟生殖行为和能力及其在不同环境生理条件下的变异 [D]. 武汉: 华中农业大学, 2010. |
| [26] | 姜玉英, 康爱国, 王春荣, 孟正平, 曾娟. 草地螟产卵和取食寄主种类初报. 中国农学通报, 2011, 27(7): 266-278. |
| [27] | 中国农业科学院植物保护研究所主编. 中国农作物病虫害 (第二版). 北京: 中国农业出版社, 1995: 805-805. |
| [28] | 郭维栋, 马柱国, 姚永红. 近50年中国北方土壤湿度的区域演变特征. 地理学报, 2003, 58(S1): 83-90. |
| [29] | 赵媛媛, 何春阳, 李晓兵, 黄庆旭, 杨洋. 干旱化与土地利用变化对中国北方草地与农牧交错带耕地自然生产潜力的综合影响评价. 自然资源学报, 2009, 24(1): 123-135. |