 2015, Vol. 35
2015, Vol. 35文章信息
- 丁俊祥, 邹杰, 唐立松, 刘卫国
- DING Junxiang, ZOU Jie, TANG Lisong, LIU Weiguo
- 克里雅河流域荒漠-绿洲交错带3种不同生活型植物的光合特性
- Photosynthetic characteristics of three different life-form plants in the desert-oasis ecotone of Keriya River Basin
- 生态学报, 2015, 35(3): 733-741
- Acta Ecologica Sinica, 2015, 35(3): 733-741
- http://dx.doi.org/10.5846/stxb201310302623
-
文章历史
- 收稿日期:2013-10-30
- 网络出版日期:2014-07-07






2. 中国科学院大学, 北京 100049;
3. 中国科学院阜康荒漠生态系统研究站, 阜康 831505;
4. 新疆大学资源与环境科学学院, 绿洲生态重点实验室, 乌鲁木齐 830046
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Fukang Station of Desert Ecology, Chinese Academy of Sciences, Fukang 831505, China;
4. Key Laboratory of Oasis Ecology, College of Resources and Environment Science, Xinjiang University, Urumqi 830046, China
新疆和田地区于田县位于克里雅河流域,是典型的荒漠-绿洲交错带,同时是干旱区生态系统退化与待恢复的关键区域[1]。由于绿洲外围恶劣的环境条件以及人类频繁的生产活动,致使于田地区经常遭受土壤次生盐渍化、风沙、水土流失、草场退化等一系列严重的环境问题。植被是绿洲生态系统的核心和支柱,在防风固沙、保持水土、生态系统恢复等方面发挥着重要作用[2]。然而,于田地区自然生长的植被种类较少、群落结构简单且覆盖度极低(<11%),主要植被有白刺(Nitraria tangutorum)、柽柳(Tamarix ramosissima)、盐穗木(Halostachys caspica)、芦苇(Phragmites australis)、花花柴(Karelinia caspia)、胡杨(Populus euphratica)等。因此,研究于田绿洲自然植被对荒漠环境的适应特性和机理,将有助于荒漠-绿洲交错带生态系统的恢复与保护。
光合作用是植物在长期进化过程中形成的一种对环境的适应,具有明显的时空变化特征,是植物生长的重要决定因素。近年来,针对干旱区植物的光合生理特性有较多报道,如王珊珊等[3]探讨了新疆古尔班通古特沙漠南缘多枝柽柳的光合作用及水分利用的生态适应性。张利刚等[4]比较研究了多枝柽柳、沙枣、梭梭、头状沙拐枣的光合、水分生理特性。韩刚等[5]研究了不同土壤水分条件下4种沙生灌木的光合光响应特性。然而,大多的研究集中在同种或同生活型植物光合特性的探讨,对荒漠—绿洲交错带内不同生活型植物光合特性的比较研究仍不多见。芦苇、柽柳、胡杨是于田地区草本、灌木、乔木3类生活型植物中的广布种,3种植物的生长面积曾达23.092、93.161、116.74 km2,是构成于田绿洲防止风沙侵袭绿色屏障的重要组成部分[6]。另外,3种植物对干旱、盐碱化、多风沙等恶劣环境具有较强的适应性,在维持绿洲生态系统平衡以及荒漠河岸林构建和保护中发挥着重要作用。本试验选取于田荒漠-绿洲交错带自然生长的芦苇、柽柳、胡杨为实验对象,通过对比3种植物光合特性的差异及其与环境因子间的关系,探讨3者光合生理过程在生态系统交错带的适应特性和机制,为荒漠-绿洲交错带退化植被的恢复和保护提供科学理论依据,同时可为交错带内环境胁迫下植被生态适应性的研究提供借鉴。
1 材料与方法 1.1 研究区与材料实验于2013年7月中旬至8月中旬在位于新疆和田地区于田县境内荒漠—绿洲交错带进行,地处东经81°14′,北纬37°02′。本研究区属暖温带内陆荒漠气候,主要特点是:四季分明、昼夜温差大,热量资源丰富,光照充足,降水稀少,蒸发量大,春夏多风沙和浮尘等灾害天气,多年平均气温为12.4 ℃,多年平均降水量47.7 mm,潜在蒸发量是2432.1 mm[7]。本实验选取克里雅河两岸一定范围内生长的芦苇、柽柳、胡杨为实验材料,其中所选芦苇植株的平均高度和直径分别为(75±0.065)m、(0.5±0.1) cm;柽柳的平均株高和冠幅分别为(2.75±0.15)m、(3.85±0.54)m;胡杨平均高度和胸径分别为(8.7±0.89)m、(0.13±0.04)m。
1.2 测定指标与方法选择生长健壮、长势一致、同一叶位的成熟叶片,从当地时间8:00到19:00,每隔1 h采用便携式光合系统分析仪LI-6400(Li-Cor,Lincoln,NE,USA)分别测定芦苇、柽柳、胡杨叶片的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)等气体交换参数,同时测定胞间CO2浓度(Ci)、光合有效辐射(PAR)、大气温度(Ta)、叶面温度(Tl)、空气相对湿度(RH)等环境因子,每次测定重复3次取值,并取3组叶片作组内重复。实验采用2 cm×3 cm标准叶室,测定时将叶片尽量平铺于整个叶室,避免相互遮挡,光合测定结束后剪下叶室中的叶片,经扫描后用CI-400图像分析软件(CIAS,CID Inc. USA)进行叶面积校正,并根据实际叶面积折算Pn、Gs、Tr等气体交换参数。3种植物叶片的气孔限制值(LS)按公式:LS=1-Ci/Ca×100%[8]计算。选择晴朗天气,于9:00至11:00运用LI-6400便携式光合作用系统测定叶片的净光合速率(Pn),使用开放气路,空气流量为0.5 L/min,叶温控制在28—30 ℃,水蒸汽压差控制在2 Kpa,CO2浓度390 μmol/mol。测定前叶片经过光诱导,光照强度梯度设定为0、60、80、100、200、400、600、800、1000、1200、1400、1600、1800 μmol m-2 s-1。测定时由系统每隔3 min自动记录每个光强下芦苇、柽柳和胡杨叶片的净光合速率(Pn),进行Pn-PAR响应曲线的拟合。另外,将人工光源光强设定为1500 μmol m-2 s-1,温湿度条件与上述一致,通过CO2小钢瓶供应CO2气体,将CO2浓度设定为60、80、100、200、300、400、600、800、1000、1200、1400、1600、1800 μmol/mol共13个水平,由系统每隔2 min自动记录每个CO2浓度所对应的净光合速率,进行Pn-CO2响应曲线拟合。表观量子效率(AQY)的计算用低光强下(200 μmol m-2 s-1以下)光响应曲线的斜率表示,羧化效率(CE)用低CO2浓度下(200 μmol/mol以下)Pn-CO2响应曲线的斜率表示[9, 10, 11]。每种植物测定3株,在各自光强和CO2浓度下重复3次读数取均值。
1.3 数据统计分析采用Excel对测定数据进行简单处理,借助SPSS18.0软件进行相关、偏相关、多元回归和通径分析等处理,并运用origin9.0软件进行绘图。
2 研究结果 2.1 3种植物净光合速率及其主要影响因子日变化野外自然状态下,3种植物净光合速率及主要影响因子的日变化如图 1。3种植物生长环境的光合有效辐射(PAR)和气温(Ta)的日变化均为单峰曲线,峰值分别出现在14:00、15:00附近;空气相对湿度(RH)的日变化趋势与气温(Ta)相反,为一凹形曲线,且变幅较大,在17:00以后,湿度开始有上升趋势(图 1)。

|
| 图 1 3种植物净光合速率及其影响因子的日变化 Fig. 1 Diurnal changes of net photosynthetic rate and its affecting factors of three plants |
晴朗天气下,芦苇和胡杨叶片净光合速率(Pn)的日变化呈明显不对称的双峰曲线,“午休”现象明显,而柽柳在经历Pn峰值后有一短暂波动,然后呈下降趋势(图 1)。芦苇和胡杨叶片净光合速率的主峰(分别为16.2 μmol m-2 s-1和19.5 μmol m-2 s-1)值出现在12:00至13:00之间,柽柳叶片净光合速率的主峰(15.0 μmol m-2 s-1)出现在14:00左右,较芦苇和胡杨晚;芦苇和柽柳净光合速率的次峰值(分别为12.2 μmol m-2 s-1和8.1 μmol m-2 s-1)出现在16:00左右,而胡杨次峰值(13.6 μmol m-2 s-1)出现在17:00附近,稍晚于芦苇和柽柳。3种荒漠植物叶片净光合速率日均值表现为胡杨>芦苇>柽柳,但差异不显著(表 1)。
| 参数 Parameter | 芦苇 Phragmites australis | 柽柳 Tamarix ramosissima | 胡杨 Populus euphratica |
| 数据为平均值±标准误;同行不同字母表示种间在0.05水平差异显著 | |||
| 净光合速率Net photosynthetic rate (Pn) / (μmol m-2 s-1) | 6.90±0.63a | 5.21±1.5a | 7.92±0.71a |
| 气孔导度Stomatal conductance ( Gs) / (mol m-2 s-1) | 0.16±0.01a | 0.27±0.07b | 0.23±0.01c |
| 胞间CO2浓度Intercellular CO2 concentration (Ci) / (μmol/mol) | 309.25±8.72b | 340.48±13.77c | 324.78±7.67b |
| 蒸腾速率Transpiration rate ( Tr) / (mmol m-2 s-1) | 5.63±0.37a | 6.71±0.97b | 6.32±0.11b |
| 叶面水汽压亏缺Vapor pressure deficit (VPD)/kPa | 3.20±0.21c | 3.03±0.39c | 2.59±0.12c |
| 气孔限制值Stomatal limitation value ( LS)/% | 0.23±0.02c | 0.17±0.04b | 0.19±0.02b |
3 种植物的叶片气孔导度(Gs)的日变化与Pn大体一致,为一双峰曲线(图 1),早晨气孔导度快速升高,后缓慢降低,12:00左右气孔进一步关闭,气孔导度降幅加大,14:00过后随气孔开放程度的增加,气孔导度增大。其中,芦苇、胡杨在12:00左右各出现一峰值,且叶片气孔导度在16:00出现峰值后又开始缓慢减小;柽柳在10:00左右出现第1个峰值,随后一直呈下降趋势。3种植物气孔导度日平均值差异显著,并以柽柳的气孔导度日均值最大(表 1)。芦苇和柽柳叶片胞间CO2浓度(Ci)与其气孔限制值(LS)的日变化方向完全相反,分别呈正“V”和倒“V”型,而胡杨Ci日变化呈波动下降。芦苇和柽柳的Ci上午缓慢下降,最低值出现在13:00—14:00之间,随后逐渐上升并有一定波动,而LS上午逐渐升高,至14:00左右达到最大值,此后随Ci缓慢上升,LS则缓慢下降(图 1)。3种植物中,柽柳与芦苇和胡杨的Ci值差异显著,芦苇与柽柳和胡杨的LS值差异显著(表 1)。
蒸腾速率的日变化趋势与净光合速率不同,呈现不稳定的波动变化(图 1)。在14:00之前,3种植物的蒸腾速率整体呈上升趋势,这与叶片温度的变化较一致(图 1)。比较气孔导度与蒸腾速率的变化(图 1)发现,12:00以后因气孔逐渐关闭,气孔导度大幅下降,蒸腾速率也随之下降,除胡杨外,其他2种植物的蒸腾速率均维持在相对较低水平。这表明干旱区植物的气孔调节在适应缺水环境时起着重要作用,可以减少植物因蒸腾作用导致过度失水所面临的风险,同时确保植物能够获得充足的CO2气体进行光合作用。3种植物中,胡杨蒸腾速率曲线变化幅度最大(1.5—13.7 mmol m-2 s-1),芦苇和柽柳的变化幅度相对较小(3.2—12.0 mmol m-2 s-1)。3种植物中,芦苇与柽柳和胡杨蒸腾速率的日均值差异显著,柽柳Tr日均值大于芦苇和柽柳(表 1)。3种植物叶面大气水蒸气压亏缺(VPD)的日变化趋势呈钟形曲线,14:00之前快速上升,之后缓慢下降(图 1),柽柳和胡杨VPD的日变化幅度较芦苇小,这可能与深根系植物在土壤水分利用上的优势有关,然而3种植物间的VPD日均值差异不显著(表 1)。
2.2 净光合速率与主要生理生态因子的关系 2.2.1 相关分析对3种植物叶片净光合速率与主要生理生态因子进行相关分析得知:芦苇叶片净光合速率与气孔导度、蒸腾速率和光合有效辐射极显著相关,与叶温显著相关;柽柳和胡杨叶片净光合速率与气孔导度和蒸腾速率极显著相关;3种植物与胞间CO2浓度和大气蒸汽压亏缺呈负相关关系,但均没达到显著水平(表 2)。
| 植物种类 Plant species | 气孔导度 Stomatal conductance | 胞间CO2浓度 Intercellular CO2 concentration | 水汽压压亏缺 Vapor pressure deficit | 蒸腾速率 Transpiration rate | 叶片温度 Leaf temperature | 光合有效辐射 Photosynthetically active radiation |
| *,* *分别表示在0.05和0.01水平上显著 | ||||||
| 芦苇 Phragmites australis | 0.93* * | -0.53 | -0.47 | 0.98* * | 0.69* | 0.92* * |
| 柽柳 Tamarix ramosissima | 0.98* * | -0.59 | -0.34 | 0.99* * | 0.20 | 0.67 |
| 胡杨 Populus euphratica | 0.89* * | -0.49 | -0.22 | 0.99* * | 0.49 | 0.72 |
3 种植物净光合速率日变化服从正态分布(P>0.05),因此,可以对Pn(y)与Tr(x1)、PAR(x2)、Gs(x3)、Ci(x4)、VPD(x5)、TL(x6)6个影响因子进行多元逐步回归分析,得到如下最优回归方程:
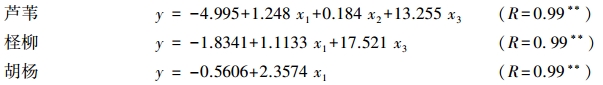
3 个回归方程的可靠性分别达98.84%、99.02%和97.34%,离回归标准误均较小(分别为0.63、0.76和0.86 μmol m-2 s-1),此外,经F检验、复相关系数检验和偏相关系数检验,回归方程均达到极显著水平(表 3)。这表明3种植物叶片净光合速率与相关因子间的线性关系成立,且回归方程具有较小的预测误差。结果显示:Tr(x1)、PAR(x2)、Gs(x3)对芦苇,Tr(x1)、Gs(x3)对柽柳,Tr(x1)对胡杨的净光合速率分别有重要影响,据此可对3种植物净光合速率与各自主要的影响因子进行通径分析。
| 植物种 Plant species | 偏相关系数 Partial correlation coefficient | t检验值 t test value | 显著水平P Significant level | |
| 芦苇 | r(y,x1) | 0.90 | 12.31 | 0.00010 |
| Phragmites australis | r(y,x2) | 0.67 | 5.26 | 0.00010 |
| r(y,x3) | 0.60 | 4.38 | 0.0099 | |
| 柽柳 | r(y,x1) | 0.71 | 5.66 | 0.00010 |
| Tamarix ramosissima | r(y,x3) | 0.52 | 3.39 | 0.012 |
| 胡杨Populus euphratica | r(y,x1) | 0.88 | 43.09 | 0.00090 |
通径分析结果表明:(1) Tr、PAR和Gs对芦苇净光合速率的影响总效应表现为Tr>Gs>PAR,即Tr对芦苇净光合速率的影响最大,通径系数为0.98,其中直接影响效应占主导,为0.63,其次是通过光合有效辐射和气孔导度的间接效应;Tr、PAR和Gs对芦苇Pn的直接效应表现为Tr>PAR>Gs。(2) Tr、Gs对柽柳叶片净光合速率的作用效应表现为Tr>Gs,即蒸腾速率对柽柳净光合速率的直接效应大于气孔导度的直接作用效果,两者均通过彼此对净光合速率的间接影响共同构成对净光合速率的总效应,分别为0.99、0.98。(3) Tr主要通过自身的直接效应对胡杨的净光合速率产生作用,通径系数为0.99。具体结果见表 4。
| 植物种类 Plant species | 影响因子 Affecting factor | 直接效应 Direct effect | 间接效应 Indirect effect | 总效应 Total effect | ||
| by Tr | by PAR | by Gs | ||||
| 芦苇Phragmites australis | Tr | 0.63 | 0.19 | 0.16 | 0.98 | |
| PAR | 0.51 | 0.19 | 0.22 | 0.92 | ||
| Gs | 0.50 | 0.18 | 0.25 | 0.93 | ||
| 柽柳Tamarix ramosissima | Tr | 0.72 | 0.26 | 0.98 | ||
| Gs | 0.67 | 0.31 | 0.98 | |||
| 胡杨Populus euphratica | Tr | 0.99 | 0.99 | |||
人工设置不同光强下,芦苇、柽柳和胡杨叶片的净光合速率在较低光强时均随光照强度的增加呈线性增加趋势,待光强上升到一定范围,净光合速率上升的趋势变缓,直至最高点前仍呈正相关关系,而当光强超过一定范围,净光合速率下降,转变为负相关关系(图 2)。芦苇、柽柳和胡杨叶片净光合速率对光强的响应曲线均以一元二次曲线方程拟合较为理想(表 5)。根据所拟合的一元二次曲线方程,计算3种植物的光补偿点(LCP)、光饱和点(LSP)、最大光合速率(Pmax)和表观量子效率(AQY),结果见表 6。

|
| 图 2 3种植物叶片光合作用的光响应曲线 Fig. 2 The light-photosynthetic response curves in the leaves of three plants |
| 植物种类 Plant species | Pn-PFD曲线回归方程 Regression equations between Pn and PFD | Pn-CO2曲线回归方程 Regression equations between Pn and CO2 contents |
| 芦苇Phragmites australis | y=-9×10-6x2+0.0278x+0.0428 R2=0.99,P<0.0010 | y=-2×10-5x2+0.0468x-2.6739 R2=0.99,P<0.0010 |
| 柽柳Tamarix ramosissima | y=-1×10-5x2+0.0283x-2.1602 R2=0.99,P<0.0010 | y=-2×10-5x2+0.0466x-2.2075 R2=0.98,P<0.0010 |
| 胡杨Populus euphratica | y=-1×10-5x2+0.031x-0.3846 R2=0.99,P<0.0010 | y=-1×10-5x2+0.0413x-2.4749 R2=0.98,P<0.0010 |
| 植物种类 Plant species | 最大净光 合速率 Maximal net photosynthetic rate/ (μmol m-2 s-1) | 光补偿点 Light compensation point/ (μmol m-2 s-1) | 光饱和点 Light saturation point/ (μmol m-2 s-1) | 表观量子效率 Apparent quantum yield/ (mol/mol) | CO2补偿点 CO2 compensation point/ (μmol/mol) | CO2饱和点 CO2 saturation point/ (μmol/mol) | 羧化效率 Carboxylation efficiency/ (mol m-2 s-1) |
| 芦苇Phragmitesaustralis | 24.09a | 29.40a | 1548.80a | 0.039a | 70.30a | 1658.80a | 0.048a |
| 柽柳Tamarix ramosissima | 19.70a | 83.50b | 1690.20a | 0.034b | 76.80a | 1434.20b | 0.055a |
| 胡杨Populus euphratica | 23.70a | 33.70a | 1607.50a | 0.041a | 89.80b | 1870.30c | 0.049a |
本研究的3种植物,光补偿点的大小依次为柽柳>胡杨>芦苇,柽柳与芦苇和胡杨间差异显著;光饱和点大小依次为柽柳>胡杨>芦苇,但差异不显著;表观量子效率大小依次为胡杨>柽柳>芦苇,柽柳与芦苇和胡杨间差异显著(表 6)。
2.4 净光合速率对CO2的响应图 3表示在饱和光强下,3种干旱区植物叶片净光合速率与CO2浓度的响应关系,从设置的不同CO2浓度所对应的净光合速率可以看出,随CO2浓度的增加,净光合速率由负变正,并迅速上升,当浓度达到400 μmol/mol左右后,净光合速率增大的趋势逐渐减弱,随后缓慢下降。经统计分析和比较,芦苇、柽柳和胡杨叶片净光合速率对CO2浓度的响应曲线均以一元二次曲线方程拟合较为理想(表 5)。根据所拟合的一元二次曲线方程,计算3种植物的CO2补偿点(CCP)、CO2饱和点(CSP)和羧化效率(CE),具体结果见表 6。3种植物的CO2补偿点的大小依次为胡杨>柽柳>芦苇,差异显著。CO2饱和点的大小依次为胡杨>芦苇>柽柳,3者之间差异显著。羧化效率反映了植物在给定环境条件下对CO2的同化能力。由表 3可以看出,3种植物的羧化效率在0.05 μmol m-2 s-1左右,差异不显著,表明3者在自然条件下对CO2的同化能力差异不大。

|
| 图 3 3种植物叶片光合作用的CO2响应曲线 Fig. 3 The CO2-photosynthetic response curves in the leaves of three plants |
光合作用是植物体内极为重要的代谢过程,它的强弱对于植物生长、产量及其抗逆性都具有十分重要的影响,因而可将光合作用作为判断植物生长和抗逆性强弱的指标[12]。本研究中净光合速率由高到低依次是胡杨、芦苇、柽柳,同时胡杨也具有较高的最大净光合速率和气孔导度,表明3种不同生活型植物中胡杨具有较强的光合能力。罗青红[12] 、邓雄[13]对荒漠植物光合特性的研究也得出同样结论,并认为胡杨属高光合树种,通过保持较高的气孔导度来保证较高的碳同化速率,以适应当地恶劣的气候条件。柽柳属深根系植物,依赖地下水补给维持正常生理活动[14],以高水分消耗为代价而最大化光合作用的碳获取[15],研究也表明柽柳的蒸腾速率高于胡杨,且显著高于芦苇,但是其净光合速率为3者中最小。
3 种植物叶片净光合速率的日变化呈不对称双峰曲线,同化高峰出现在13:00—14:00之间,与光照强度和气孔导度日变化规律相似,表明这3种植物具有在不利环境条件出现之前的短暂时间内充分利用水和光资源的能力,这是对环境适应的表现。另外,3种植物净光合速率日变化的次峰值出现在16:00—17:00之间,较同化高峰分别下降24.7%、46.0%、30.3%,这主要是由于午间光合抑制效应在午后尚未完全消除[16],且不同物种间植物的光合能力及适应光合抑制的能力存在差异。
3种植物存在明显的光合“午休”现象,这与大多数对荒漠植物光合特性研究的结论一致[4, 12, 13, 17, 18, 19]。引起光合“午休”现象的因素很多,最主要的影响因素有:(1)午间气温升高,羧化效率下降;(2)由于缺水而引起的气孔导度降低或造成叶片局部水分胁迫[20]。根据Farquhar和Sharkey[21]关于光合速率降低原因的判断原则,当Pn和Gs下降时,若Ci降低而LS增加,则表明气孔导度的降低是光合速率减小的主要原因,若Ci增加而LS降低,则为非气孔限制造成净光合速率的降低。据此分析,14点以前,芦苇和胡杨Pn下降的主要由气孔限制引起,即随着辐射的增强、空气湿度降低、温度升高,使原本较适宜的光合作用环境变得恶劣,Gs降低导致CO2供应不足,Ci浓度降低引起Pn下降;而15:00以前,柽柳Pn的下降为非气孔限制引起,即因柽柳叶肉光合活性下降造成[22],这与王珊珊对新疆古尔班通古特沙漠南缘多枝柽柳光合特性的研究一致[3]。18:00以后,3种植物净光合速率的下降主要由于光合有效辐射的减弱。此外,研究区内的大气温度在中午时分已高达40 ℃以上,高温对植物叶肉细胞的光合活性也会产生一定影响,因此,是否因高温引起植物羧化效率下降而造成Pn降低有待进一步验证。
3.2 3种植物净光合速率影响因子的比较植物叶片光合效率除与自身因素如叶绿素含量、叶片厚度、叶片成熟程度密切相关,还受光照强度、气温、空气相对湿度、土壤水分含量等外界环境因子影响[17]。偏相关分析及通径分析结果表明,影响芦苇、柽柳和胡杨叶片净光合速率的主要生理生态因子及作用效应不尽相同。其中,Tr、PAR和Gs是芦苇Pn的主要影响因子,Tr、Gs是柽柳Pn的主要影响因子,胡杨Pn主要受Tr的直接影响作用,而且Tr对3种植物叶片净光合速率的直接效应大于其它环境因子。这表明3种植物的净光合速率在气孔严格调控的基础之上,与叶片蒸腾速率存在紧密联系,净光合速率的变化是植物自身生理和环境生态因子综合作用的复杂过程[23]。然而,环境因子与光合作用间的关系并不是恒定不变的,会随着天气条件、土壤水分条件、土壤养分、植物种类以及生育期的不同而发生大幅度的变化,例如:付为国等[9]对湿地芦苇光合特性的研究得出芦苇叶片净光合速率与光合有效辐射相关极显著,与蒸腾速率和气孔导度相关显著,而段晓男[24]对乌梁素海野生芦苇光合作用影响因子的研究则认为光合速率与光合有效辐射关系最为密切,周洪华[2]对塔里木河下游胡杨气体交换特性的研究得出胡杨净光合速率主要受大气CO2浓度和光合有效辐射的影响。因此,今后的研究工作需要结合植物的光合生理指标及抗逆性生理指标,从深层次上揭示影响植物光合作用的生理生态因子及其作用机理,以综合评价荒漠植物对环境的适应特性和机制。
3.3 3种植物光合模型参数的比较光饱和点(LSP)和光补偿点(LCP)反映植物对光照条件的适应范围,分别体现了植物对强光和弱光的利用能力。本研究中,芦苇和胡杨均具有较高的LSP和较低的LCP,说明在同一生境下,2种植物对弱光和强光的利用能力都较强。表观量子效率反映了植物在弱光下吸收、转化和利用光能能力[25],一般情况下,植物表观量子效率介于0.03与0.07之间,值越高说明植物叶片对光能的转化效率越高。本研究显示,芦苇和胡杨的表观量子效率显著高于柽柳,说明芦苇和胡杨对荒漠环境具有较好的适应性。3种不同生活型植物间表观量子效率的差异可能与各自可利用的土壤水分含量有关,水分亏缺和水分过多都会导致植物表观量子效率降低,但不同植物间,土壤水分与表观量子效率的定量关系仍不清楚,需要更深入的研究[26]。
| [1] | 樊自立, 马英杰, 王让会, 季方, 张立远. 干旱区内陆河流域生态系统的类型及整治途径——以新疆为例. 中国沙漠, 2000, 20(4): 393-397. |
| [2] | 周洪华, 陈亚宁, 李卫红, 陈亚鹏. 塔里木河下游胡杨气体交换特性及其环境解释. 中国沙漠, 2008, 28(4): 665-672. |
| [3] | 王珊珊, 陈曦, 王权, 包安明, 曹晓明, 李平衡. 新疆古尔班通古特沙漠南缘多枝柽柳光合作用及水分利用的生态适应性. 生态学报, 2011, 31(11): 3082-3089. |
| [4] | 张利刚, 曾凡江, 刘波, 刘镇, 安桂香, 李海峰, 袁娜. 绿洲-荒漠过渡带四种植物光合及生理特征的研究. 草业学报, 2012, 21(1): 103-111. |
| [5] | 韩刚, 赵忠. 不同土壤水分下4种沙生灌木的光合光响应特性. 生态学报, 2010, 30(15): 4019-4026. |
| [6] | 陈荷生. 克里雅河流域生态环境变化与水资源合理利用. 中国沙漠, 1990, 10(3): 1-12. |
| [7] | 海米提, 依米提, 冷中笑, 高前兆. 塔里木盆地南缘于田绿洲近50a来气候变化特征分析. 中国沙漠, 2010, 30(3): 654-659. |
| [8] | Berry J A, Downton W J S. Environmental regulation of photosynthesis. In: Govindjee NY ed. Photosynthesis Vol. Ⅱ. New York: Academic Press, 1982: 263-343. |
| [9] | 付为国, 李萍萍, 卞新民, 吴沿友, 曹秋玉. 镇江北固山湿地芦苇光合日变化的研究. 西北植物学报, 2006, 26(3): 496-501. |
| [10] | 潘晓云, 曹琴东, 王根轩, 杨晓燕. 扁桃与桃光合作用特征的比较研究. 园艺学报, 2002, 29(5): 403-407. |
| [11] | 朱万泽, 王金锡, 薛建辉, 鄢武先. 四川桤木光合生理特性研究. 西南林学院学报, 2001, 21(4): 196-204. |
| [12] | 罗青红, 李志军, 伍维模, 韩路. 胡杨、灰叶胡杨光合及叶绿素荧光特性的比较研究. 西北植物学报, 2006, 26(5): 983-988. |
| [13] | 邓雄, 李小明, 张希明, 叶万辉. 四种荒漠植物的光合响应(英文). 生态学报, 2003, 23(3): 598-605. |
| [14] | Anderson J E. Factors controlling transpiration an photosynthesis in Tamarax Chinensis lour. Ecology, 1982, 63(1): 48-56. |
| [15] | 许皓, 李彦, 谢静霞, 程磊, 赵彦, 刘冉. 光合有效辐射与地下水位变化对柽柳属荒漠灌木群落碳平衡的影响. 植物生态学报, 2010, 34(4): 375-386. |
| [16] | 葛滢, 王晓月, 常杰. 不同程度富营养化水中植物净化能力比较研究. 环境科学学报, 1999, 19(6): 690-692. |
| [17] | 常宗强, 冯起, 苏永红, 吴雨霞, 司建华, 席海洋. 额济纳绿洲胡杨的光合特征及其对光强和CO2浓度的响应. 干旱区地理, 2006, 29(4): 496-502. |
| [18] | 朱军涛, 李向义, 张希明, 林丽莎, 杨尚功. 塔克拉玛干沙漠南缘3种荒漠植物光合特性的比较. 干旱区地理, 2012, 35(1): 171-177. |
| [19] | 田新民, 赵长明, 邓建明, 张晓玮, 陈拓, 任贾文, 王根轩. 沿民勤绿洲荒漠过渡带分布的4种优势植物光合生理响应. 草业学报, 2011, 20(4): 108-115. |
| [20] | Küppers M, Wheeler A M, Küppers B I L, Krischbaum M U F, Farquhar G D. Carbon fixation in eucalypts in the field. Oecologia, 1986, 70(2): 273-282. |
| [21] | Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis. Annual Rreview of Plant Physiology, 1982, 33(1): 317-345. |
| [22] | 付士磊, 周永斌, 何兴元, 陈玮. 干旱胁迫对杨树光合生理指标的影响. 应用生态学报, 2006, 17(11): 2016-2019. |
| [23] | 郭天财, 王之杰, 王永华. 不同穗型小麦品种旗叶光合作用日变化的研究. 西北植物学报, 2002,22(3): 554-560. |
| [24] | 段晓男, 王效科, 冯兆忠, 欧阳志云. 乌梁素海野生芦苇光合和蒸腾特性研究. 干旱区地理, 2004,27(4): 637-641. |
| [25] | 李合生.现代植物生理学.北京:高等教育出版社,2002: 125-125. |
| [26] | Davies F S, Flore J A. Short-term flooding effects on gas exchange and quantum yield of rabbiteye blueberry (Vaccinium ashei Reade). Plant physiology, 1986, 81(1): 289-292. |