 2015, Vol. 35
2015, Vol. 35文章信息
- 徐耀粘, 江明喜
- XU Yaozhan, JIANG Mingxi
- 森林碳库特征及驱动因子分析研究进展
- Forest carbon pool characteristics and advances in the researches of carbon storage and related factors
- 生态学报, 2015, 35(3): 926-933
- Acta Ecologica Sinica, 2015, 35(3): 926-933
- http://dx.doi.org/10.5846/stxb201304190745
-
文章历史
- 收稿日期:2013-04-19
- 网络出版日期:2014-04-03






2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
森林生态系统作为一个重要的碳库在区域以及全球碳循环中起到重要的作用[1],是陆地生态系统最大的碳库。据报道,森林中储存的碳占到总量的45%左右[2],年固碳量约占陆地生态系统固碳总量的三分之二[3]。并且森林生态系统对人为因素和自然干扰比较敏感,当森林遭受破坏和退化时变成一个碳源,储存的碳以二氧化碳的方式迅速释放到大气中,而当森林在保护和恢复演替过程中则成为一个碳汇[4]。在1990到2007期间,由于森林砍伐以及土地利用方式改变导致热带地区每年(1.3±0.5) Pg C 净释放到大气中,成为一个巨大的碳源,而全球森林生态系统每年的净碳汇为(2.4±0.4)Pg C[5]。随着人类意识到森林作为一个巨大的碳汇以及在转移大气二氧化碳方面的作用,森林保护和管理活动逐渐增强[6],但是对自然和人为因素如何影响森林碳储存过程和机理,以及森林碳储量表现出的空间异质性和时间尺度上的复杂性,使得森林碳储量和储存机制研究存在很大的不确定性[7]。
目前,森林碳储量研究成为气候变化科学研究领域和国际社会关注的热点,研究的时空尺度也逐渐增大[5, 8],可以模拟和估算长时间、大尺度上的碳储量的动态变化,但是目前的研究尚集中在单一生态因子对森林碳储量的影响,而对于多个因子综合影响考虑较少。通过研究大量文献,本文阐述了气候、土壤、地形和生物因子对森林碳储量的影响,分析了生态因子之间的相互作用关系,厘清生态因子对森林碳储量的影响过程和机理,以期解决以往研究中出现的单一和混乱的局面,并提出有效的解决方法,为森林碳储存机制的研究提供新的思路和途径。
1 森林碳库特征森林生态系统通过光合作用将固定的二氧化碳主要储存到3个有机碳库:活植物碳库、土壤有机质碳库和死植物体碳库。森林生态系统中还有一些小并且难以测定碳库,例如动物和挥发有机质碳库,在研究中通常忽略。碳库容量在不同森林类型以及不同林龄中所占的比例也不尽相同,在成熟森林中,活有机体和粗木质残体中储存的碳约占森林碳总量的60%,土壤和凋落物中的碳约占40%[6]。
1.1 森林活植物碳库特征森林生态系统的碳主要储存在植被和土壤中,而森林活生物量,主要包括植物各器官(枝、干、叶和根),其中地上生物量(AGB)占有很大的比重。AGB可以通过构建异速生长方程进行比较准确的估算,其时空变化也可以通过建立永久森林样地进行重复观测来获得,而估算其它碳库的大小和变化比较困难,因此AGB的变化常作为测定森林碳动态的指标。AGB对人类活动和自然干扰比较敏感,例如:火灾、砍伐、土地利用类型改变、风暴以及病虫害,因此AGB在森林生态系统碳储存短期变化中起主要作用。AGB虽然不是唯一的碳库,但是AGB是森林生态系统中最具有动态的碳库,因此森林AGB的动态是森林碳库研究的热点区域[8]。森林地下生物量由于估算比较困难,常通过与AGB的比值进行换算[9],也有树干、皮、枝生物量估算方法的报道[10]。
1.2 森林土壤碳库特征土壤有机碳库是森林生态系统最为重要的碳库之一,也是陆地生态系统容量最大但是周转周期最慢的一个碳库。土壤有机碳库的组分主要有:腐殖质、微生物、代谢产物以及未完全分解的动植物残体,土壤表层的有机碳含量最高,随土壤深度增加而递减,一般认为100 cm以下,有机质的含量已经很少[11]。土壤有机质影响着土壤肥力以及生态系统的生产力和稳定性,土壤碳循环也是土壤氮、磷、硫循环的驱动因子。储藏在土壤中的碳由于其物理化学性质不容易被氧化,因而相对于植被碳库较稳定[12, 13],但是土壤碳库呈现出很高的空间变异性,很难预测土壤碳库大小的变化规律,尤其在土地利用方式发生改变时,例如森林砍伐和造林,Guo和Gifford[14]认为土壤碳库变化的大小和方向依赖于地上植被、土壤性质和气候。
然而最近的一项研究发现土壤中固定的碳70%来自于树根和周围生长的真菌,并认为根和真菌固定的碳要比凋落物对土壤的贡献度更大。共生真菌通过与树根的共生作用,增强了宿主植物对大气二氧化碳的吸存能力,促进植物生长并且直接将植物的光合作用产物转运并且储藏到土壤中,用于自身繁殖,死亡后个体可被土壤微生物迅速分解或者长时间储藏在土壤中。这个结果强调了北方森林中菌根真菌在土壤碳封存中的核心作用[15, 16]。而在热带森林中,也有发现固氮植物根瘤可以提高森林固碳能力的报道[17],因而有必要更深层次的研究其在森林固碳中的作用。
1.3 森林死植物碳库相比于活植被碳库和土壤碳库,森林死植被碳库比较小,在成熟林中,粗木质残体以及凋落碳库占总碳库的10%—20%[18]。森林凋落物碳库在植被碳库和土壤碳库之间起纽带的作用,促进森林植被-凋落物-土壤系统的养分循环[19]。森林凋落物通过微生物分解以有机物的形式归还土壤中,森林凋落物分解速度的快慢,直接影响着森林生态系统碳循环的速率,进而影响二氧化碳的净吸收量。相对于植被和土壤碳库,凋落物碳库表现出更大的变异性,例如在火灾或者森林砍伐下,凋落物碳库表现出快速的氧化分解或者积累。而粗木质残体碳库也类似于凋落物碳库,表现出很强的空间异质性,由于这类碳库与AGB和森林年龄没有很强的相关性,因此缺少有效的方法对这类碳库进行计量,从而也会造成了较大的估测误差[20]。
2 森林碳储量影响因子分析森林生态系统主要通过光合作用、呼吸作用来获得和散失碳,森林是否净获得或者净丢失碳依赖于这些过程的平衡,而森林碳储存能力则取决于这些生态过程对环境变化的敏感性和受影响程度[20]。森林生物量在区域和全球尺度上表现出一些清晰的分布格局[21],然而,在影响这些格局的生态因子以及从尺度向外推的程度的问题上尚存在争议[22, 23]。此文中选取气候、土壤、地形和生物4类因子对森林生物量的影响进行分析(表 1)。
从表 1可以看出,这4类生态因子均可以解释部分的AGB 空间变异,但是在因子与AGB相关关系上尚未得出一致的结论。AGB与降雨量、断面积、大树密度、平均树高以及平均胸径呈正相关,而与旱季长度呈现负相关,说明AGB的最大值应该出现在降雨量丰富、气候比较稳定(短暂或者没有旱季)并且大树比较多的区域,因此生物量最大值出现在热带[24, 25]以及部分亚热带地区[34, 41]。
2.1 气候因子对森林生物量的影响在全球尺度上,森林生物量分布随纬度梯度呈现一定的地带性规律,可见森林生物量空间分布的确受到温度、降雨量等气候因子的影响,但是温度和降雨量等气候因子与AGB是否呈现正相关关系呢?这对很多生物量以及生产力模型的校正有重要的意义。
水分调节群落和生态系统对全球变化的响应,尤其是在干旱和半干旱地区[42]。在热带地区,成熟林的生物量随着降雨梯度增大而增大[24, 43, 44],降雨量也被认为是森林生物量上限的最重要的因子[43],这可能是因为更多的降雨量延长了生长季长度从而使群落每年积累更多的生物量[26],而在比较干旱的森林中,过少的降雨量影响土壤营养的淋溶以及长时间的干旱使植物缺少水从而限制生长[45]。另外降雨的季节性也对森林生物量也有影响,研究发现AGB和胸高断面积随着降雨的季节性和旱季长度增加而降低[24, 28],造成此现象的原因可能是由于水分资源缺少加剧了根部对于水分的竞争导致地上部分获取水分减少从而限制了树的顶端生长,导致了生物量比较低[46]。
总体上,AGB呈现随温度增加而逐渐增加的趋势,但并不明显,并且预计最高生物量在亚热带湿润的常绿森林中[34, 47],而Stegen等[40]认为在湿润的热带森林中AGB随年均温升高而降低,热带云雾林则相反,另一项研究也得出AGB在温带区域内会随温度的升高而降低的结论[27]。因此在区域和全球尺度上,AGB均没有很明显的随温度的增加而增大,反而与降雨梯度上比较一致[48],Larjavaara 和Muller-Landau[27]从方法论的角度认为可能是由于取样的面积过小或有过干扰历史造成的。
2.2 土壤对森林生物量的影响在景观尺度上研究土壤与AGB的关系时发现,土壤因子对AGB空间变异的解释度达到三分之一[29, 32]。由此可见,土壤的物理化学性质在一定程度上决定着森林AGB的空间分布。从表 1可以看出几类土壤成分指标与AGB并没有得出一致性的结论,正相关、负相关,以及相关关系不明显的报道均有。肥沃的土壤可以增加树的密度和个体大小,因而积累更多的生物量;但是也有研究发现在肥沃的土壤上生物量更小的情况[22],因为肥沃的土壤可能会提高森林周转率,导致了较低的生物量[29];同时也存在土壤营养与AGB相关性不明显的情况[32, 49]。在不同的研究中,对AGB空间变异有明显相关的指标发生也很大的变化,甚至相同指标在不同的森林类型中也表现出很大的差异。例如在Laurance等[31]的研究中,粘土中的营养梯度是唯一显著的预测因子,但在Paoli等[29]的研究中则是沙土含量和有效磷影响显著。
土壤主要通过两个方式影响森林生物量:一是土壤影响群落物种组成和结构,间接的影响生物量;二是不考虑物种组成因素,森林是否在肥沃的土壤条件下获得更多的生物量,或者大树趋向于生长在更肥沃的土壤上[31]。然而,Vitousek 和Sanford[50]认为土壤肥力影响物种组成、结构以及生物量只在一些极端的情况下才能出现,也有假说认为土壤营养与AGB之间存在一种单峰曲线的关系,但是到目前为止,这个假说还没有很好的被证实[22]。
2.3 地形因素对森林生物量的影响在局域尺度上,地形通过多种方式影响森林的物种组成、结构和生物量[8]。地形因子通过影响其他变量的变化来影响AGB,例如土壤类型[4]、土壤含水量[51]和土壤营养循环[52],以及光照条件,山谷地形常会出现沟谷河流和大石等异常情况[8]。地形也会影响森林动态[53, 54]。尽管如此,地形因子在决定森林活生物量中起到重要作用[34, 41],也相对比较容易获得,借助地形因子有助于理解AGB的空间分布特点,以及环境因子之间的相互作用。
地形因子主要包括海拔、坡度、坡位、坡向和凹凸度,由表 1可以看出,地形因子在不同地区与AGB的关系表现出很大的差异。研究发现,AGB最大值出现在地形比较平坦和低海拔区域,同时海拔和凹凸度是AGB最强的预测因子[34]。而Lin等[8]研究中发现亚热带低海拔山脊拥有最大的生物量,其次是低海拔和高海拔山谷。Mascaro等[36]在热带森林中研究发现坡度是最强的驱动因子并且AGB在陡峭的坡上拥有最大值。Marshall等[33]认为最大AGB出现在坡度小的中海拔区域,在高海拔地区,植物生长可能受到环境胁迫。可见,地形因子在不同的森林类型中起到的作用并不相同,有可能是在不同森林类型中地形因子影响着不同的环境因子,从而造成地形因子对AGB的影响存在较大的争议。
2.4 生物因子对森林碳储量的影响影响AGB的生物因子主要包括物种组成和森林结构,由于胸径、树高和木材密度常被作为生物量方程的参数,所以这些生物因子与AGB有着很强的正相关性[39]。另外,树密度和胸高断面积也与AGB有很强的相关性[24],从AGB与胸径断面积和大树密度的强烈相关关系可以看出,更高的AGB值出现在大树密度比较大的森林[28, 29]。
表 1中,木材密度是唯一有争议的生物因子。木材密度是一个重要的估测变量,也是一个常见的生物量方程参数,尤其是在物种丰富木材密度范围比较大的森林中,木材密度在森林碳储存方面起到的作用更为明显[28, 39]。植物在不同的生长速率以及生活史策略情况下,木材密度表现出很大的差异[55],另外,木材密度在不同演替阶段的森林生物量差异中起着重要作用[56]。因此研究木材密度与AGB的关系有着重要意义。Baker等[37]认为如果忽略木材密度作为方程参数,AGB的估测质量将会很差,文章研究发现木材密度可以解释很大部分的AGB从东到西的变异,但Stegen等[40]发现木材密度与AGB之间没有一致性的关系,可以是正相关的、负相关的,还可以是无关联。从这些研究结果可以看出,木材密度对森林碳储存的确起到了一定的作用,但是影响的程度可能与不同的地区不同的森林类型有关。
3 误差分析和研究新方向在研究中,选用不同样地和物种,以及实验方法的差异都可以忽略或屏蔽某一因子的作用,或者过分夸大对某一因子的依赖,导致不能得出广泛有效的结论[21]。本文对选取的文献的研究方法和主要结论进行分析和总结,认为主要有三方面因素限制了对生态因子与AGB相互关系的理解:数据质量、取样方法以及生态因子的处理方法。
首先,很少有对样地的森林类型、干扰情况以及各类环境因子进行完整的描述;另外,物种鉴定以及构建生物量方程的解析木以及木材密度样本过少也能形成较大误差。其次,研究的样地常常很难形成一个较大的环境梯度,利用较小的梯度分析时通常会得出不同甚至相反的结果;取样时应考虑地形因素以及森林类型和干扰情况;取样的尺度也应该考虑,样方过小没有代表性,过大缺乏重复并且大尺度常会忽略一些生态过程。
最后,对于气候、土壤等环境因子,采取和生物因子相同的方法进行处理是不合适的(图 1和图 2),图 1中各因子解释度的加和(A-F)很可能大于1,图 2描述的是各类环境因子与AGB的关系,可以看出气候、土壤因子并不是直接的影响AGB,而是直接或者间接影响构建生物量方程的参数,进而影响AGB,将环境因子和生物因子放在同一层次来研究对森林生物量的影响是不恰当的[22],后文会提出两种有效的方法去区别和控制环境因子之间的交互作用。

|
| 图 1 生物、土壤和生物因子与AGB的空间变异关系的分解图 Fig. 1 Venn diagram illustrating the decomposition of spatial variation in AGB between stand,soil and climate variables |

以前的研究未曾考虑过不同因素之间的层次关系,而是利用同一模型或方法研究不同环境因子对AGB的影响,得出的研究结果难免有差异。Baraloto等[22]建立了一个层次模型将生物因素和气候、土壤因子放在不同层次上来研究与AGB的关系:
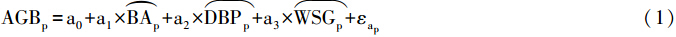
气候、土壤和生物因子对AGB空间变异的解释度从没用层次模型之前的33%、18%和90%变成用之后的3%、2%和60%,通过结果可以看到层次模型显著地提高了各因素对AGB的预测的准确度。
3.3 利用多元回归树进行生境分类环境因子和生物因子共同驱动着森林的碳储存,如何对生态因子进行有效的控制是一个挑战。Lin等[8]利用多元回归树将古田山25 hm2样地分成高、低海拔山谷和山脊4种生境,4类生境中,AGB 的极差达到最大值的四分之一,并且物种在不同的生境对AGB的贡献不同,表现出明显的生境偏好[8]。因此基于多元回归树的方法可以很好的研究生物因子对AGB空间变异的影响,为研究森林碳储存提供了一种新的思路。
4 总结和展望现在对理解森林碳储存过程和机制以及最终对森林碳储量进行有效的管理的需求日益增长[34]。虽然在不同地区均做过一些相关研究,然而气候、土壤、地形和生物因子与AGB的相关关系仍然存在较大的争议,相同的因子在不同地区对AGB的空间变异的解释度表现出极大的差异,甚至同一因子的不同指标在解释局部AGB的空间变异时也大相径庭,让研究者认为在不同地区不同森林类型可能存在着不同碳储存机制。在目前的研究中,常常将生物因子和环境因子放在同等地位上研究与AGB的关系,而在实际情况下,气候土壤等环境因子并不是直接影响AGB,而是通过影响生物量方程的参数来间接的影响AGB,这也可能是造成了不同的生态因子对AGB的空间变异影响出现混淆的原因,也让森林碳储存机制的研究止步于研究单一生态因子与AGB的相关关系。因此,以后的研究中,应该尽可能的考虑多个生态因子在不同的层次上对森林生物量的影响。另外取样方法、研究的时空尺度等因素都在都在限制着对这种关系的理解,目前这方面的研究还很少,有待于今后更多的较大时空尺度研究来进一步探究森林碳储存机制。
| [1] | Dixon R K, Solomon A M, Brown S, Houghton R A, Trexier M C, Wisniewski J. Carbon pools and flux ofglobal forest ecosystems. Science, 1994, 263(5144): 185-190. |
| [2] | Bonan G B. Forests and climate change: forcings, feedbacks, and the climate benefits of forests. Science, 2008, 320(5882): 1444-1449. |
| [3] | 王效科,冯宗炜,欧阳志云. 中国森林生态系统的植物碳储量和碳密度研究. 应用生态学报,2001, 12 (1):13-16. |
| [4] | Chauvel A, Lucas Y, Boulet R. On the genesis of the soil mantle of the region of Manaus, Central Amazonia. BrazilExperientia, 1987,43(3):234-241. |
| [5] | Pan Y D, Birdsey R A, Fang J Y, Houghton R, Kauppi P E, Kurz W A, Phillips O L, Shvidenko A, LewisS L, Canadell J G,Ciais P, Jackson R B, Pacala S W, McGuire A D, Piao S, Rautiainen A, Sitch S, Hayes D. A large and persistent carbon sink in the world's forests. Science, 2011, 333(6045): 988-993. |
| [6] | Fahey T J, Woodbury P B, Battles J J, Goodale C L, Hamburg S P, Ollinger S V, Woodall C W. Forest carbon storage: ecology, management, and policy. Frontiers in Ecology and the Environment, 2010, 8(5): 245-252. |
| [7] | 赵敏, 周广胜. 中国森林生态系统的植物碳贮量及其影响因子分析. 地理科学, 2004, 24(1): 50-54. |
| [8] | Lin D M, Lai J S, Muller-Landau H C, Mi X C, Ma K P. Topographic variation in abovegroundbiomass in a subtropical evergreen broad-leaved forest in China. PLoSONE, 2012, 7(10): e48244. |
| [9] | Cairns M A, Brown S, Helmer E H, Baumgardner G A. Root biomass allocation in the world's upland forests. Oecologia, 1997, 111(1): 1-11. |
| [10] | 曾立雄, 王鹏程, 肖文发, 万睿, 黄志霖, 潘磊. 三峡库区主要植被生物量与生产力分配特征. 林业科学, 2008, 44(8): 16-22. |
| [11] | 王绍强, 周成虎, 李克让, 朱松丽, 黄方红. 中国土壤有机碳库及空间分布特征分析. 地理学报, 2000, 55(5): 533-544. |
| [12] | Davidson E A, JanssensI A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature, 2006, 440(7081): 165-173. |
| [13] | Post W M, Kwon K C. Soil carbon sequestration and land-use change: processes and potential. Global Change Biology, 2000, 6(3): 317-327. |
| [14] | Guo L B, Gifford R M. Soil carbon stocks and land use change: a meta-analysis. Global Change Biology, 2002, 8(4): 345-360. |
| [15] | Clemmensen K E, Bahr A, Ovaskainen O, Dahlberg A, Ekblad A, Wallander H, Stenlid J, Finlay R D, Wardle D A, Lindahl B D. Roots and associated fungi drive long-term carbon sequestration in boreal forest. Science, 2013, 339(6127): 1615-1618. |
| [16] | Treseder K K, Holden S R. Fungal carbon sequestration. Science, 2013, 339(6127): 1528-1529. |
| [17] | Ruiz-Jaen M C, Potvin C. Can we predict carbon stocks in tropical ecosystems from tree diversity? Comparing species and functional diversity in a plantation and a natural forest. New Phytologist, 2011, 189(4): 978-987. |
| [18] | Chaturvedi R K, Raghubanshi A S, Singh J S. Carbon density and accumulation in woody species of tropical dry forest in India. Forest Ecology and Management, 2011, 262(8): 1576-1588. |
| [19] | 杨晓菲, 鲁绍伟, 饶良懿, 耿绍波, 曹晓霞, 高东. 中国森林生态系统碳储量及其影响因素研究进展. 西北林学院学报, 2011, 26(3): 73-78. |
| [20] | Ryan M G, Harmon M E, Birdsey R A, Giardina C P, Heath L S, HoughtonR A, Jackson R B, McKinley D C, Morrison J F, Murray B C,Pataki D E, Skog K E. A synthesis of the science on forests and carbon for U.S. forests. Ecological Society of America: Issues in Ecology, 2010, 13: 1-16. |
| [21] | 吕超群, 孙书存. 陆地生态系统碳密度格局研究概述. 植物生态学报, 2004, 28(5): 692-703. |
| [22] | Baraloto C, Rabaud S, Molto Q, Blanc L, Fortunel C, Herault B, Davila N, Mesones I, Rios M, Valderrama E. Disentangling stand and environmental correlates of aboveground biomass in Amazonian forests. Global Change Biology, 2011, 17(8): 2677-2688. |
| [23] | Houghton R A, Lawrence K T, Hackler J L, Brown S. The spatial distribution of forest biomass in the Brazilian Amazon: a comparison of estimates. Global Change Biology, 2001, 7(7): 731-746. |
| [24] | Malhi Y, Wood D, Baker T R, Wright J, Phillips O L, Cochrane T, Cochrane T, Meir P, Chave J, Almeida S, Arroyo L, Higuchi N, Killeen T J, Laurance S G, Laurance W F, Lewis S L, Monteagudo A, Neill D A, Vargas P N, Pitman N C A, Quesada C A, Salomao R, Silva J N M, Lezama A T, Terborgh J, Martinez R V, Vinceti B. The regional variation of aboveground live biomass in old-growth Amazonian forests. Global Change Biology, 2006, 12(7): 1107-1138. |
| [25] | Chave J, Condit R, Aguilar S, Hernandez A, Lao S, Perez R. Error propagation and scaling for tropical forestbiomass estimates. Philosophical Transactions of the Royal Society B: Biological Sciences, 2004, 359(1443): 409-420. |
| [26] | Baker T R, Swaine M D, Burslem D F R P. Variation in tropical forest growth rates: combined effects of functional group composition and resource availability. Perspectives in Plant Ecology, Evolution and Systematics, 2003, 6(1/2): 21-36. |
| [27] | Larjavaara M, Muller-Landau H C. Temperature explains global variation in biomass among humid old-growth forests. Global Ecology and Biogeography, 2012, 21(10): 998-1006. |
| [28] | SlikJ W F, AibaSI, Brearley F Q, Cannon C H, Forshed O, Kitayama K, Nagamasu H, Nilus R, Payne J, Paoli G,Poulsen A D, Raes N, Sheil D, Sidiyasa K, Suzuki E, van Valkenburg J L C H. Environmental correlates of tree biomass, basal area, wood specific gravity and stem density gradients in Borneo's tropical forests. Global Ecology and Biogeography, 2010, 19(1): 50-60. |
| [29] | Paoli G D, Curran L M, Slik J W F. Soil nutrients affect spatial patterns of aboveground biomass and emergent tree density in southwestern Borneo. Oecologia, 2008, 155(2): 287-299. |
| [30] | Quesada CA, Lloyd J, Schwarz M, Baker TR, Phillips O L, PatiLño S, Czimczik C, Hodnett M G, Herrera R,Arneth A, Lloyd G, Malhi Y, Dezzeo N, Luizão F J, Santos A J B, Schmerler J, Arroyo L, Silveira M, Filho N P, Jimenez E M, Paiva R, Vieira I, Neill D A, Silva N, PeLñuela M C, Monteagudo A, Vásquez R, Prieto A, Rudas A, Almeida S, Higuchi N, Lezama A T, López-González G, Peacock J, Fyllas N M, Dávila E A, Erwin A, di Fiore A, Chao K J, Honorio E, Killeen T, PeLña C A, Pitman N, Vargas P N, Salomão R, Terborgh J, Ramírez H. Regional and large-scale patterns in Amazon forest structure and function are mediated by variations in soil physical and chemical properties. Biogeosciences Discussions, 2009, 6: 3993-4057. |
| [31] | Laurance W F, Fearnside P M, Laurance S G, Delamonica P, Lovejoy T E, Rankin-de Merona J M, Chambers J Q, Gascon C. Relationship between soils and Amazon forest biomass: a landscape-scale study. Forest Ecology and Management, 1999, 118(1/3): 127-138. |
| [32] | DeWaltS J, ChaveJ. Structure and biomass of four lowland Neotropical forests. Biotropica, 2004, 36(1): 7-19. |
| [33] | Marshall A R, Willcock S, Platts P J, Lovetta J C, Balmfordd A, Burgessd N D, Lathama J E, Munishih P K T, Saltera R, Shirimah D D, Lewisc S L. Measuring and modelling above-ground carbon and tree allometry along a tropical elevation gradient. Biological Conservation, 2012, 154: 20-33. |
| [34] | McEwan R W, LinYC, Sun I, Hsieh C F, Su S H, Chang L W, Song G Z M, Wang H H, Hwong J L, Lin K C, Yang K C, Chiang J M. Topographic and biotic regulation of aboveground carbon storage in subtropical broad-leaved forests of Taiwan. Forest Ecology and Management, 2011, 262(9): 1817-1825. |
| [35] | de Castilho C V, MagnussonWE, de AraújoR N O, LuizãoRCC, LuizãoF J, LimaAP, HiguchiN. Variation in aboveground tree live biomass in a central Amazonian Forest: Effects of soil and topography. Forest Ecology and Management, 2006, 234(1/3): 85-96. |
| [36] | Mascaro J, Asner G P, Muller-Landau H C, van Breugel M, Hall J, Dahlin K. Controls over aboveground forest carbon density on Barro Colorado Island, Panama. Biogeosciences, 2011, 8(6): 1615-1629. |
| [37] | Baker T R, Phillips O L, Malhi Y, Almeida S, Arroyo L, Di Fiore A, Erwin T, Killeen T J, Laurance S G, LauranceWF,Lewis S L, Lloyd J, Monteagudo A, Neill D A, PatiLño S, Pitman N C A, Silva J N M, Martínez R V. Variation in wood density determines spatial patterns inAmazonian forest biomass. Global Change Biology, 2004, 10(5): 545-562. |
| [38] | Chave J, Condit R, Lao S, Caspersen JP, Foster RB, Hubbell SP. Spatial and temporal variation of biomass in a tropical forest: results from a large census plot in Panama. Journal of Ecology, 2003, 91(2): 240-252. |
| [39] | Chave J, Andalo C, Brown S, Cairns M A, Chambers J Q, Eamus D, Fölster H, Fromard F, Higuchi N, KiraT, Lescure J P, Nelson B W, Ogawa H, Puig H, Riéra B, Yamakura T. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia, 2005, 145(1): 87-99. |
| [40] | Stegen J C, Swenson N G, Valencia R, Enquist B J, Thompson J. Above-ground forest biomass is not consistently related to wood density in tropical forests. Global Ecology and Biogeography, 2009, 18(5): 617-625. |
| [41] | McEwan R W, Muller R N. Spatial and temporal dynamics in canopy dominance of an old-growth central Appalachian forest. Canadian Journal of Forest Research, 2006, 36(6): 1536-1550. |
| [42] | Smith S D, Huxman T E, Zitzer S F, Charlet T N, Housman D C, Coleman J S, Fenstermaker L K, Seemann J R, Nowak R S. Elevated CO2 increases productivity and invasive species success in an arid ecosystem. Nature, 2000, 408(6808): 79-82. |
| [43] | Brown S, Lugo A E. The storage and production of organic matter in tropical forests and their role in the global carbon cycle.Biotropica, 1982, 14(3): 161-187. |
| [44] | Saatchi S S, Houghton R A, Dos Santos Alvalá R C, Soares J V, YuY F. Distribution of aboveground live biomass in the Amazon basin. Global Change Biology, 2007, 13(4): 816-837. |
| [45] | Eamus D. Ecophysiological traits of deciduous and evergreen woody species in the seasonally dry tropics. Trends in Ecology and Evolution, 1999, 14(1): 11-16. |
| [46] | Meinzer F C, Andrade J L, Goldstein G, Holbrook N M, Cavelier J, Wright S J. Partitioning of soil water among canopy trees in a seasonally dry tropical forest. Oecologia, 1999, 121(3): 293-301. |
| [47] | LuyssaertS, Inglima I, JungM, RichardsonAD, ReichsteinM, Papale D, Piao S L, Schulze E D, Wingate L, MatteucciG,Aragao L, Aubinet M, Beer C, Bernhofer C, Black K G, Bonal D, Bonnefond J M, Chambers J, Ciais P, Cook B, Davis K J, Dolman A J, Gielen B, Goulden M, Grace J, Granier A, Grelle A, Griffis T, Grünwald T, Guidolotti G, Hanson P J, Harding R, Hollinger D Y, Hutyra L R, Kolari P, Kruijt B, Kutsch W, Lagergren F, Laurila T, Law B E, Le Maire G, Lindroth A, Loustau D, Malhi Y, Mateus J, Migliavacca M, Misson L, Montagnani L, Moncrieff J, Moors E, Munger J W, Nikinmaa E, Ollinger S V, Pita G, Rebmann C, Roupsard O, Saigusa N, Sanz M J, Seufert G, Sierra C, Smith M L, Tang J, Valentini R, Vesala T, Janssens I A. CO2 balance of boreal, temperate, and tropical forests derived from a global database. Global Change Biology, 2007, 13(12): 2509-2537. |
| [48] | Keith H, Mackey B G, Lindenmayer D B. Re-evaluation of forest biomass carbon stocks and lessons from the world's most carbon-dense forests. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(28): 11635-11640. |
| [49] | Ashton P S, Hall P. Comparisons of structure among mixed dipterocarp forests of north-western Borneo. Journal of Ecology, 1992, 80(3): 459-481. |
| [50] | Vitousek P M, Sanford R L Jr. Nutrient cycling in moist tropical forest. Annual Review of Ecology and Systematics, 1986, 17(1): 137-167. |
| [51] | Becker P, RabenoldPE, IdolJR, SmithAP. Water potential gradients for gaps and slopes in a Panamanian tropical moist forest's dry season. Journal of Tropical Ecology, 1988, 4(2): 173-184. |
| [52] | Luizão R C C, Luizão F J, Paiva R Q, Monteiro T F, Sousa L S, Kruijt B. Variation of carbon and nitrogen cycling processes along a topographic gradient in a central Amazonian forest. Global Change Biology, 2004, 10(5): 592-600. |
| [53] | Bellingham P J, Tanner E V J. The Influence of Topography on Tree Growth, Mortality, and Recruitment in a Tropical Montane Forest.Biotropica, 2000, 32(3): 378-384. |
| [54] | Robert A. Simulation of the effect of topography and tree falls on stand dynamics and stand structure of tropical forests. Ecological Modelling, 2003, 167(3): 287-303. |
| [55] | Whitmore T C, Burslem D F R P. Major disturbances in tropical rainforests // Newbery D M, Prins H H T, Brown N D. Dynamics of Tropical Communities. Oxford: Blackwell Science, 1998: 549-565. |
| [56] | Valencia R, Condit R, Muller-Landau H C, Hernandez C, Navarrete H. Dissecting biomass dynamics in a large Amazonian forest plot. Journal of Tropical Ecology, 2009, 25(5): 473-482. |