 2015, Vol. 35
2015, Vol. 35文章信息
- 刘茂柯, 唐玉明, 赵珂, 任道群, 姚万春, 田新惠
- LIU Maoke, TANG Yuming, ZHAO Ke, REN Daoqun, YAO Wanchun, TIAN Xinhui
- 浓香型白酒窖泥放线菌的群落结构及其多样性
- Analysis of actinobacteria community and diversity in the pit mud of chinese luzhou-flavour liquor
- 生态学报, 2015, 35(3): 858-864
- Acta Ecologica Sinica, 2015, 35(3): 858-864
- http://dx.doi.org/10.5846/stxb201304100664
-
文章历史
- 收稿日期:2013-04-10
- 网络出版日期:2014-04-03






2. 四川农业大学资源环境学院, 温江 611130
2. College of Resource and Environment, Sichuan Agricultural University, Wenjiang 611130, China
浓香型白酒深受广大消费者喜爱,其酿造生产依靠窖泥微生物的协同作用[1]。揭示窖泥微生物群落结构及其多样性可为酿酒功能微生物的系统选育,人工窖泥配方的优化改良提供理论依据。放线菌是与人类生活密切相关的一类特殊细菌,其代谢产物一直是食品、医学工业的研究热点。已有研究显示,放线菌可能促进浓香型白酒主体香味物质的形成[2]。但目前对窖泥放线菌的研究仍然较少,对其系统发育、多样性结构亦鲜有报道。这必将弱化对浓香型白酒发酵机理的正确认识。
近年来,以PCR 为基础的免培养技术已成为现代微生物生态学研究的重要手段,在对极端环境群落结构的监测方面具有传统方法不具备的优势[3]。因此,本研究采用PCR-DGGE 分析浓香型白酒窖泥放线菌的群落结构及其多样性,以期为相关研究的深入奠定一定的理论基础。
1 材料与方法 1.1 实验材料泸州老窖1、50、100 和400 年窖池的窖壁泥和窖底泥。窖壁泥取样方法是从窖池四壁中心的一点各取50 g 窖泥,混合均匀。窖底泥为窖底四个角落和中心的一点各取50 g 窖泥,混合均匀。样品冷冻密封保存。
1.2 实验方法 1.2.1 窖泥样品预处理及总DNA 的提取按黄永光等[4]的方法进行样品预处理后提取总DNA。用DNA 纯化试剂盒 (上海生工)纯化总DNA 后,于-20 ℃ 保存备用。
1.2.2 放线菌16S rRNA 基因的PCR 扩增首先采用引物F243 和R1378 进行扩增,再以PCR 产物的10 倍稀释液为模板,使用F984-GC 和R1378 进行嵌入式扩增[5]。反应体系(50 μL):10×缓冲液2 μL,dNTPs (2.5 mmol/L) 2 μL,引物(10 μmol/L)各0.5 μL,Taq DNA 聚合酶(5 U/μL) 2 μL,BSA(25 mg/mL) 2.5 μL,模板DNA 1 μL,ddH2O 补足50 μL。两轮PCR 程序相同,参照李小林等[6]的方法进行。用1.0% 琼脂糖凝胶电泳检测PCR 产物。
1.2.3 扩增产物的DGGE 与测序PCR 产物采用Bio-Rad 变性梯度凝胶电泳仪进行分析。聚丙烯酰胺凝胶浓度为8%,变性范围40%—70%。使用1×TAE 缓冲液,100 V 电泳10 min 进胶,再在70 V、60 ℃ 条件下电泳12 h,银染法染色[7],最后用数码相机照相。将DGGE 图谱上的主要条带切胶回收,用F984 和R1378[5]引物对片断进行PCR 扩增后克隆测序。
1.2.4 DGGE 图谱分析DGGE 图谱用Quantity One 4.1.1 软件(Bio-Rad) 分析,测定图谱条带数与光强度,计算条带优势度指数(di)、多样性指数(H)、均匀度指数(EH) 和群落结构相似性系数(SC)[8]。
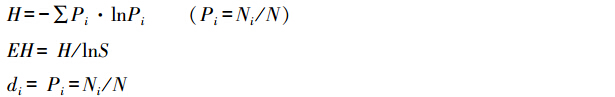
式中,Ni 为DGGE 图谱中条带i 的光强度;N 为所有条带的光强度之和;S 为条带数目。
1.2.5 系统发育分析将序列通过DNAMAN 进行处理,用BLAST 将序列与GenBank 数据库进行相似性比较,获得相近模式菌株序列,用MEGA 5.0 中Neighbor-Joining 法构建系统发育树。
2 结果与分析 2.1 窖泥放线菌16S rRNA 基因的PCR 扩增窖泥总DNA 经PCR 扩增后获得放线菌的16S rRNA 基因目的片断,经1% 琼脂糖电泳检测,得到的片段大小约为400 bp (图 1)。

|
| 图 1 窖泥放线菌16S rRNA 基因的PCR 产物电泳图谱 Fig. 1 Electrophoresis graph of the amplified 16S rRNA fragments from pit mud M:DL2000 Marker;图中数字1—4 代表 1、50、100 和 400 年窖壁泥;5—8 代表 1、50、100 和 400年窖底泥 |
图 2为窖泥放线菌16S rRNA 基因片段的DGGE 图谱。各样品PCR 产物经DGGE 分离后共得到17 条不同位置的条带。B1—B10 为本实验的测序条带。其中,B3、B5 和B10 为所有样品共有,表明不同窖泥的主要条带一致,放线菌群落结构具有相似性。其余条带为部分样品所有,其中B9 仅在50a以上窖壁泥中检出,B7、B8 在100a以上窖底泥中检出,说明群落结构因窖池窖龄和部位的不同而存在差异。这可能导致酿造白酒的酒质不同。

|
| 图 2 窖泥放线菌16S rRNA 基因片段的DGGE 图谱 Fig. 2 DGGE patterns of 16S rRNA gene extracted from Actinobacteria in the pit mud M:DL2000 Marker;图中数字1—4 代表 1、50、100 和 400 年窖壁泥;5—8 代表 1、50、100 和 400年窖底泥 |
优势度(di) 反映某物种在所有物种中所占的数量比例。结果显示,DGGE 图谱上每一条带在不同样品中的di 值差异较大,但共有条带B3、B5 和B10 的di 值在不同样品中均维持较高水平,表明它们代表窖泥中的优势菌群。对于窖壁泥样品,以上条带di 值随窖龄延长而逐步降低;在窖底泥中,B3、B5 逐渐上升,B10 则表现出先降后升的趋势(图 3)。此结果揭示,窖泥放线菌群落结构的演替可能具有一定的规律。

|
| 图 3 不同窖龄窖泥主要条带的优势度变化 Fig. 3 Changing laws of dominant degree of the main bands in the pit with different ages M:DL2000 Marker;图中数字1—4 代表 1、50、100 和 400 年窖壁泥;5—8 代表 1、50、100 和 400年窖底泥 |
多样性指数(H) 主要基于物种数量反映生态环境中的物种多样性,均匀度指数(EH) 反映物种的分配情况。结果如图 4所示,不同样品H 和EH 值存在一定差异。但同窖龄样品中,除1a样品外,其余窖底泥H 值较低,EH 值较高。不同窖龄样品中,相对1 年窖壁泥(1.74),50a窖壁泥H 值(2.16) 有明显上升,随后上升程度减小;100 和400 年间无明显变化,分别为2.28 和2.27。窖底泥H 值则随窖龄延长而平稳下降,维持在1.73—2.07 之间。另一方面,窖壁和窖底泥EH 值均随窖龄延长而波动下降,分别为0.986—0.991 和 0.971—0.994。窖底泥的波动幅度较大,且以1到50年的变化最明显。

|
| 图 4 不同窖龄窖泥的多样性指数和均匀度指数 Fig. 4 Shannon-weaver index and evenness for the pit with different ages |
此结果提示,在窖泥老熟过程中,窖壁泥放线菌主要表现为新种属的生长;窖底泥主要为原种属的消亡。在此过程中,1到50年间放线菌群落结构变化较大。
群落结构相似性系数(SC) 反映不同样品菌群种类的相似程度。窖泥放线菌SC 结果见图 5。由图可知,当SC 值小于0.53,窖泥样品分为3 个大族。3 号样品单独为一个族;2、4 和5 号为第二族,其余样品为第三族。其中,窖底泥SC 值较高,相近窖龄样品SC 值随窖龄延长而上升,为0.46—0.82;其次是窖壁泥,为0.31—0.62;同窖龄样品SC 值较低,为0.31—0.46。结果说明,窖泥空间分布可能是影响放线菌种类差异的主要原因。在窖泥老熟过程中,窖底泥菌群种类逐渐稳定。

|
| 图 5 窖泥放线菌群落结构相似性树状图 Fig. 5 Dendrogram of homology coefficient of Actinobacteria in the pit mud M:DL2000 Marker;图中数字1—4 代表 1、50、100 和 400 年窖壁泥;5—8 代表 1、50、100 和 400年窖底泥 |
序列同源性结果显示,序列B5、B9、B10 和最似模式菌株Olsenella profusa CCUG 45371T、Corynebacterium minutissimum NCTC 10288T、Atopobium parvulum VPI 0546T的同源性较低,分别为95%、90% 和93%。其余序列和最似模式菌株同源性为97%—100%。
系统发育分析结果如图 6 所示,所有序列大致聚为3个族。B1、B2、B4 与Streptomyces 属模式菌株聚在第1个族;B3、B5、B10和Olsenella 属以及Atopobium 属聚在第2个族,并以与Olsenella 属的同源性较高;其余序列与Corynebacterium 属聚在第3个族。结果显示窖泥放线菌可归属于以上4 个属,具有一定的多样性。

|
| 图 6 部分放线菌DGGE 条带系统发育分析 Fig. 6 Phylogenetic analysis of 16S rRNA gene sequences of Actinobacteria from DGGE bands |
传统观点认为,放线菌在浓香型白酒酿造中的作用有限,使得相关研究一直未受重视。有关窖泥放线菌多样性的研究仅见于张超等[9]在2011 年的报道。他们采用平板法从多粮浓香型窖泥分离出Streptomyces、Massilia、Nocardiopsis 3 个属,其中以Streptomyces 属占优。这与本实验结果有差异,主要原因可能与分析手段不同有关。在环境微生物的检测中,往往需将可培养和免培养技术结合,以获得更详尽的科学数据。本实验和前人的研究互为补充,进一步揭示出窖泥蕴藏着丰富的放线菌资源,应当引起研究者足够重视。其次,原因可能还与本实验样品为单粮窖泥有关。单粮和多粮酿造采用的原料与工艺不同,但这是否会对群落结构及多样性造成影响有待证实。
本实验发现1 到50年间放线菌群落结构变化较大,此后趋于稳定。这与已报道的窖泥菌群结构变化[10, 11]相似,表明窖泥微生物演替具有共性。罗 惠波等[12]认为,窖泥细菌菌群相似性以同窖龄样品最高。同部位样品的相似性较高,演替更显规律性,说明放线菌群落结构和演替具有自身特殊性。放线菌的生物总量随窖泥老熟而递增[1]。这与本实验窖壁泥光强度总值(N) 变化一致。不同的是,窖底泥变化截然相反,这可能和放线菌的好氧特性有关。具体到某一种群,无论窖壁或窖底泥,N 值变化均不明显。由此可见,放线菌生物总量的变化主要受多样性影响。
实验检测到的种属中,Streptomyces 属为多数样品共有。王涛等[2]的研究显示,来源于酿造环境的Streptomyces 属放线菌可产生酯、酸、醛、酮和醇类等浓香型白酒的香味物质。他推测该种属可能在酿造中扮演重要角色。Streptomyces 属在张超等[9]和本研究中均被检出,说明其在窖泥中稳定存在,可能对浓香型白酒酿造具有普遍作用。此外,其余种属作用尚无报道。优势属Atopobium 和Olsenella 的主要生境是人体,两者因亲缘关系较近常被合并研究[13, 14, 15]。它们存在于窖泥中有其合理性,并可能对酿造生产起到积极作用。首先,其专性厌氧特性与酿造环境相适应,这可能是其在窖底泥的优势度不断提高,同时也是未被张超等[9]分离到的原因之一;其次,代谢产物如乳酸,乙酸和丁酸等[15, 16, 17]与浓香型白酒香味物质密切相关;最后,一些菌种如A.parvulum、O.umbonata 均具有硫化物分解能力[13, 15],这可能起到预防窖泥老化板结的作用。另外,Corynebacterium 属主要来源于乳制品、蔬菜和土壤等生境[18]。岳元媛等[19]发现1573 窖底泥也有该种属存在。本实验显示,该种属在50a以上窖泥中均可检出,并以在窖底泥的丰度较高。同时,某些菌种如C. tuberculostearicum 可能为100a以上的高龄窖底泥特有。虽然其作用尚不清楚,但这提示是否可从菌群的特有性来衡量窖泥老熟程度?这也是值得深入研究的课题。
4 结语本实验采用DGGE 技术初步解析了浓香型白酒窖泥放线菌的菌落结构及其多样性。结果表明,窖泥放线菌归属于Olsenella、Atopobium、Streptomyces 和Corynebacterium 4 个属。不同窖龄窖泥放线菌群落结构和多样性具有一定差异,但同部位窖泥的菌群演替显示出一定规律性。此结果有助于人们进一步认识放线菌在浓香型白酒酿造中所扮演的角色,从中筛选一些功能菌种并加以利用可能对人工窖泥配方的优化改良大有裨益。
本实验供试窖泥采自泸州老窖不同年份窖池,酿造产品分别面向不同层次消费市场,样品来源具有代表性。由于窖泥样品,尤其是高龄窖泥的稀有性,在今后工作中,应加强样品的收集,增加样品的全面性,使研究结果具有更广泛的适用性,以利于相关研究的深入发展。
| [1] | 张肖克, 黄永光, 胡晓瑜. 窖泥糟醅发酵过程微生物多态性特征. 酿酒科技, 2006, (1): 65-68, 72-72. |
| [2] | 王涛, 游玲, 赵冬, 冯瑞章, 王松, 冯学愚, 林强. 浓香型白酒酿造相关放线菌挥发性产物分析. 食品科学, 2012, 33(14): 184-187. |
| [3] | Pace N R. A molecular view of microbial diversity and the biosphere. Science, 1997, 276(5313): 734-740. |
| [4] | 黄永光, 黄平, 涂华彬. 窖泥微生物总DNA的提取纯化研究. 酿酒科技, 2004, (3): 41-42. |
| [5] | Heuer H, Krsek M, Baker P, Smalla K, Wellington E M. Analysis of actinomycete communities by specific amplification of genes encoding 16S rRNA and gel-electrophoresis separation in denaturing gradients. Applied and Environmental Microbiology, 1997, 63(8): 3233-3241. |
| [6] | 李小林, 辜运富, 张小平, 涂仕华, 伍仁军. 烟草成熟期根际硝化细菌种群的结构及其多样性. 中国农业科学, 2011, 44(12): 2462-2468. |
| [7] | Bassam B J, Caetano-Anollés G, Gresshoff P M. Fast and sensitive silver staining of DNA in polyacrylamide gels. Analytical Biochemistry, 1991, 196(1): 80-83. |
| [8] | Hill T C J, Walsh K A, Harris J A, Moffett B F. Using ecological diversity measures with bacterial communities. FEMS Microbiology Ecology, 2006, 43(1): 1-11. |
| [9] | 张超, 赵东, 王涛, 游玲, 冯瑞章, 王松, 何建蓉. 多粮浓香型白酒生产中放线菌的多样性. 食品科学, 2011, 32(23): 192-196. |
| [10] | 施思, 王海英, 张文学, 邓宇, 赖登燡, 范鏊. 浓香型白酒不同窖泥的微生物群落特征分析. 酿酒科技, 2011, (5): 38-41. |
| [11] | 王海英, 张文学, 施思, 范鏊, 邓宇. 水井坊窖底泥微生物群落结构的DGGE分析. 中国酿造, 2012, 31(2): 38-41. |
| [12] | 罗惠波, 甄攀, 黄治国. 浓香型白酒窖池细菌群落. 微生物通报, 2010, 37(11): 1621-1627. |
| [13] | Copeland A, Sikorski J, Lapidus A, Nolan M, Rio T G D, Lucas S S, Chen F, Tice H, Pitluck S, Cheng J F, Pukall R, Chertkov O, Brettin T, Han C, Detter J C, Kuske C, Bruce D, Goodwin L, Ivanova N, Mavromatis K, Mikhailova N, Chen A, Palaniappan K, Chain P, Rohde M, Göker M, Bris-tow J, Eisen J, Markowitz V, Hugenholtz P, Kyrpides N C, Klenk H P, Detter J C. Complete genome sequence of Atopobium parvulum type strain (IPP 1246T). Standards in Genomic Sciences, 2009, 1(2): 166-173. |
| [14] | Dewhirst F E, Paster B J, Tzellas N, Coleman B, Downes J, Spratt D A, Wade W G. Characterization of novel human oral isolates and cloned 16Sr DNA sequences that fall in the family Coriobacteriaceae: description of Olsenella gen. nov., reclassification of Lactobacillus uli as Olsenella uli comb. nov. and description of Olsenella profusa sp. Nov.. International Journal of Systematic and Evolutionary Microbiology, 2001, 51(5): 1797-1804. |
| [15] | Kraatz M, Wallace R J, Svensson L. Olsenella umbonata sp. nov., a microaerotolerant anaerobic lactic acid bacterium from the sheep rumen and pig jejunum, and emended descriptions of Olsenella, Olsenella uli and Olsenella profusa. International Journal of Systematic and Evolutionary Microbiology, 2011, 61(4): 795-803. |
| [16] | Jovita M R, Collins M D, Sjödén B, Falsen E. Characterization of a novel Atopobium isolate from the human vagina: description of Atopobium vaginae sp. nov.. International Journal of Systematic and Evolutionary Microbiology, 1999, 49(4): 1573-1576. |
| [17] | Collins M D, Wallbanks S. Comparative sequence analyses of the 16S rRNA genes of Lactobacillus minutus, Lactobacillus rimae and Streptococcus parvulus: Proposal for the creation of a new genus Atopobium. FEMS Microbiology Letters, 1992, 95(2-3): 235-240. |
| [18] | Feurer C, Clermont D, Bimet F, Candréa A, Jackson M. Glaser P, Bizet C, Catherine D. Taxonomic characterization of nine strains isolated from clinical and environmental specimens, and proposal of Corynebacterium tuberculostearicum sp. nov.. International Journal of Systematic and Evolutionary Microbiology, 2004, 54(4): 1055-1061. |
| [19] | 岳元媛, 张文学, 刘霞, 胡承, 张宿义. 浓香型白酒窖泥中兼性厌氧细菌的分离鉴定. 微生物学通报, 2007, 34(2): 251-255. |